Dimensionality and dynamics in the behavior of C. elegans
a r X i v :0705.1548v 2 [q -
b i o .O T ] 17 M a y 2007
Dimensionality and dynamics in the behavior of C.elegans
Greg J Stephens,a ?c Bethany Johnson–Kerner,a William Bialek a,b and William S Ryu a
a
Lewis–Sigler Institute for Integrative Genomics,b Joseph Henry Laboratories of Physics,
and c Center for the Study of Brain,Mind and Behavior,Princeton University,Princeton,New Jersey 08544USA
(Dated:February 1,2008)
A major challenge in analyzing animal behavior is to discover some underlying simplicity in complex motor actions.Here we show that the space of shapes adopted by the nematode C.elegans is surprisingly low dimensional,with just four dimensions accounting for 95%of the shape variance,and we partially reconstruct ‘equations of motion’for the dynamics in this space.These dynamics have multiple attractors,and we ?nd that the worm visits these in a rapid and almost completely deterministic response to weak thermal stimuli.Stimulus-dependent correlations among the di?erent modes suggest that one can generate more reliable behaviors by synchronizing stimuli to the state of the worm in shape space.We con?rm this prediction,e?ectively “steering”the worm in real time.
INTRODUCTION
The study of animal behavior is rooted in two diver-gent traditions.One approach creates well-controlled situations,in which animals are forced to choose among a small discrete set of behaviors,as in psychophysical experiments [1].The other,taken by ethologists [2],de-scribes the richness of the behaviors seen in more natural contexts.One might hope that simpler organisms pro-vide model systems in which the tension between these approaches can be resolved,leading to a fully quantita-tive description of complex,naturalistic behavior.
Here we explore the motor behavior of the nematode,Caenorhabditis elegans ,moving freely on an agar plate [3,4,5,6,7].Though lacking the full richness of a nat-ural environment,this unconstrained motion allows for complex patterns of spontaneous motor behaviors [8],which are modulated in response to chemical,thermal and mechanical stimuli [9,10,11].Using video mi-croscopy of the worm’s movements,we ?nd a low di-mensional but essentially complete description of the macroscopic motor behavior.Within this low dimen-sional space we reconstruct equations of motion which reveal multiple attractors—candidates for a rigorous def-inition of behavioral states.We show that these states are visited as part of a surprisingly reproducible response of C.elegans to small temperature changes.Correlations among ?uctuations along the di?erent behavioral dimen-sions suggest that some of the randomness in the behav-ioral responses could be removed if sensory stimuli are delivered only when the worm is at a well de?ned initial state.We present experimental evidence in favor of this idea,showing that worms can be “steered”in real time by appropriately synchronized stimuli.
EIGENWORMS
We use tracking microscopy with high spatial and tem-poral resolution to extract the two-dimensional shape
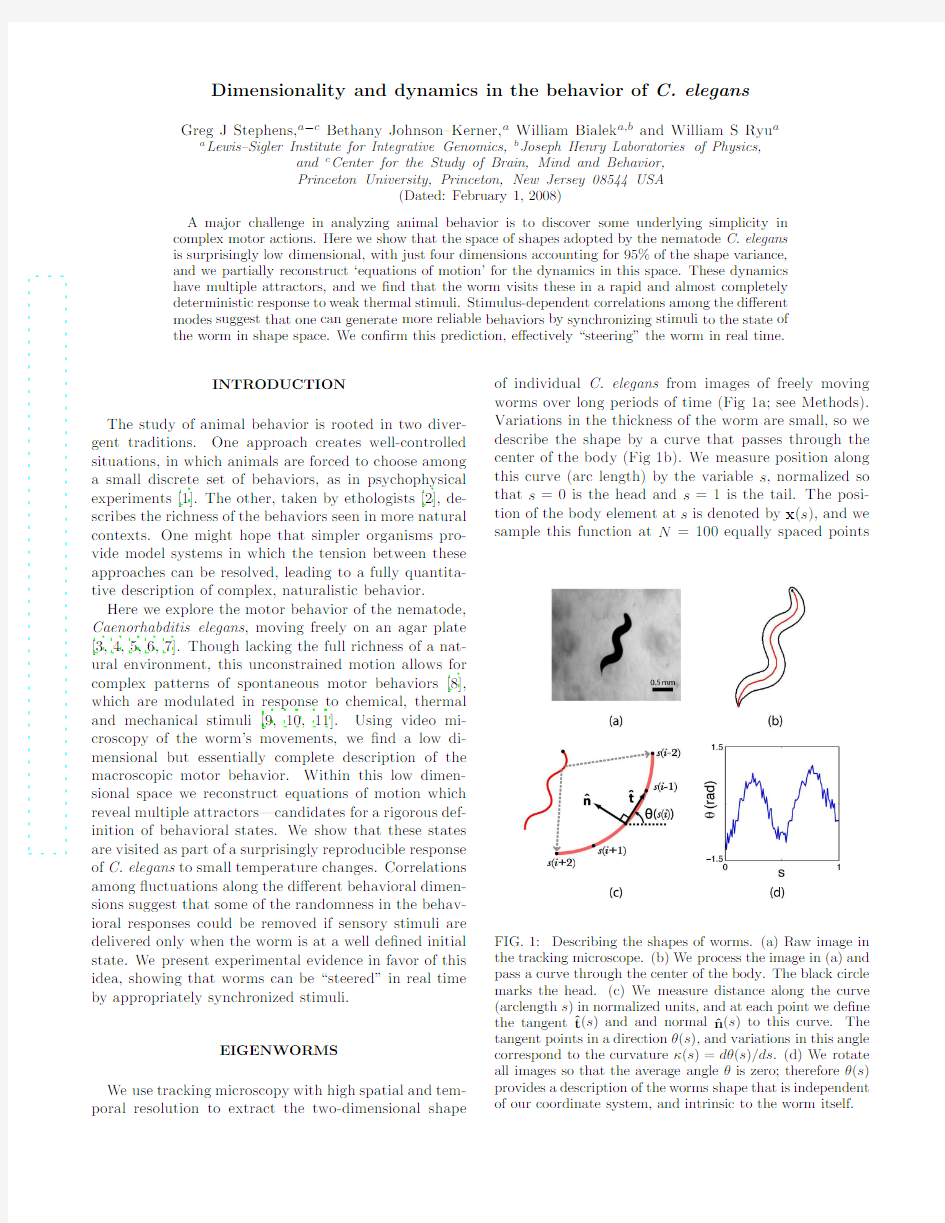
of individual C.elegans from images of freely moving worms over long periods of time (Fig 1a;see Methods).Variations in the thickness of the worm are small,so we describe the shape by a curve that passes through the center of the body (Fig 1b).We measure position along this curve (arc length)by the variable s ,normalized so that s =0is the head and s =1is the tail.The posi-tion of the body element at s is denoted by x (s ),and we sample this function at N =100equally spaced points
FIG.1:Describing the shapes of worms.(a)Raw image in the tracking microscope.(b)We process the image in (a)and pass a curve through the center of the body.The black circle marks the head.(c)We measure distance along the curve (arclength s )in normalized units,and at each point we de?ne the tangent ?t (s )and and normal ?n (s )to this curve.The tangent points in a direction θ(s ),and variations in this angle correspond to the curvature κ(s )=dθ(s )/ds .(d)We rotate all images so that the average angle θis zero;therefore θ(s )provides a description of the worms shape that is independent of our coordinate system,and intrinsic to the worm itself.
2
along the body.These variables provide an essentially complete description of the motor output.
We analyze the worm’s shapes in a way intrinsic to its own behavior,not to our arbitrary choice of coordinates. The intrinsic geometry
of a curve in the plane is de?ned by the Frenet equations[12,13],
d x(s)
ds
=κ(s)?n(s),(2)
where t(s)is the unit tangent vector to the curve,?n(s)is the unit normal to the curve,andκ(s)is the scalar cur-vature(Fig1b).If the tangent vector points in a direc-tionθ(s),thenκ(s)=dθ(s)/ds.Curvature as a function of arc length,κ(s),thus provides a“worm–centered”de-scription,but in practice this involves taking two deriva-tives and thus is noisy.As an alternative,we describe the curve byθ(s),but remove the dependence on our choice of coordinates by rotating each image so that the mean value ofθalong the body always is zero(Fig1c); this rotated version ofθ(s)contains exactly the same information asκ(s).
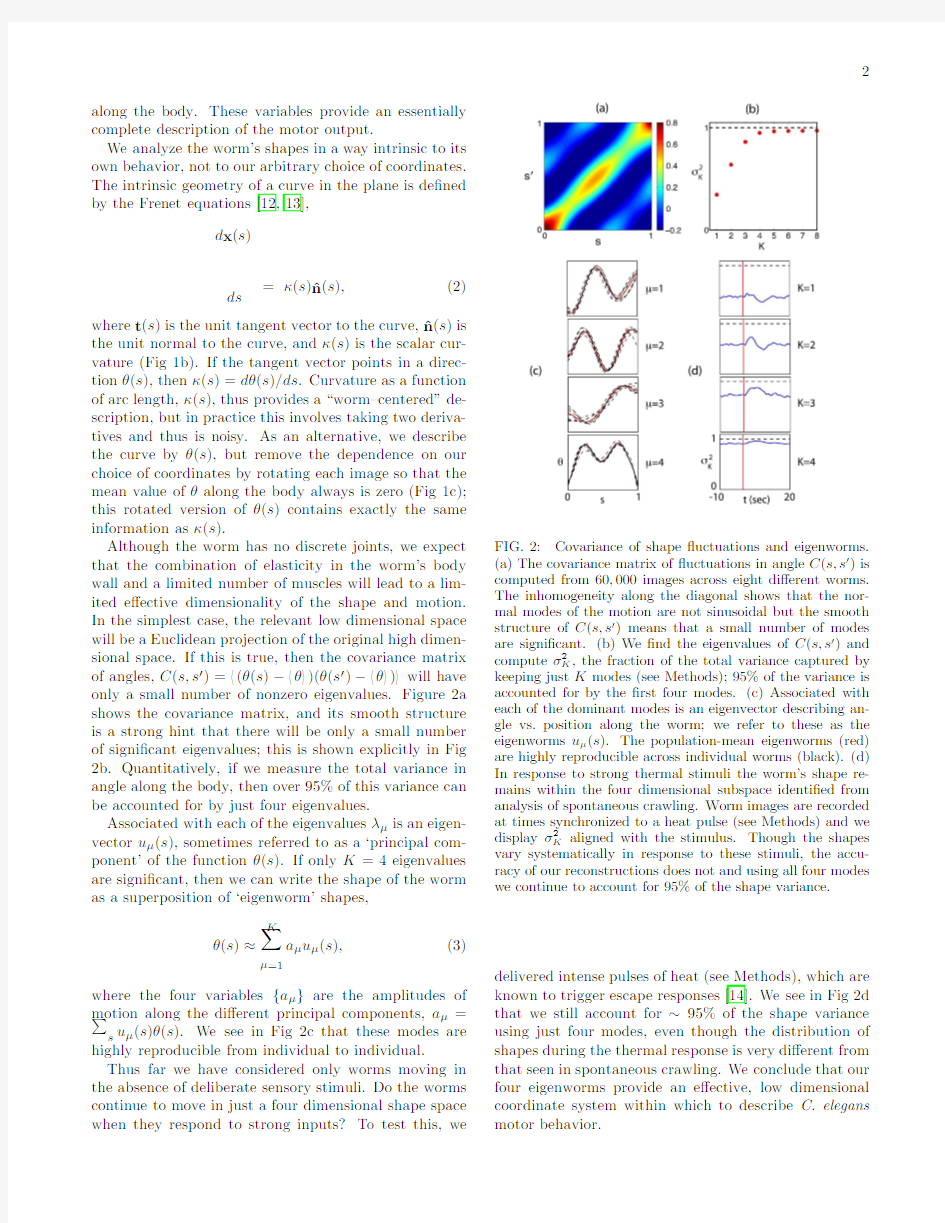
Although the worm has no discrete joints,we expect that the combination of elasticity in the worm’s body wall and a limited number of muscles will lead to a lim-ited e?ective dimensionality of the shape and motion. In the simplest case,the relevant low dimensional space will be a Euclidean projection of the original high dimen-sional space.If this is true,then the covariance matrix of angles,C(s,s′)= (θ(s)? θ )(θ(s′)? θ ) will have only a small number of nonzero eigenvalues.Figure2a shows the covariance matrix,and its smooth structure is a strong hint that there will be only a small number of signi?cant eigenvalues;this is shown explicitly in Fig 2b.Quantitatively,if we measure the total variance in angle along the body,then over95%of this variance can be accounted for by just four eigenvalues.
Associated with each of the eigenvaluesλμis an eigen-vector uμ(s),sometimes referred to as a‘principal com-ponent’of the functionθ(s).If only K=4eigenvalues are signi?cant,then we can write the shape of the worm as a superposition of‘eigenworm’shapes,
θ(s)≈
K
μ=1
aμuμ(s),(3)
where the four variables{aμ}are the amplitudes of motion along the di?erent principal components,aμ= s uμ(s)θ(s).We see in Fig2c that these modes are highly reproducible from individual to individual. Thus far we have considered only worms moving in the absence of deliberate sensory stimuli.Do the worms continue to move in just a four dimensional shape space when they respond to strong inputs?To test this,we FIG.2:Covariance of shape?uctuations and eigenworms.
(a)The covariance matrix of?uctuations in angle C(s,s′)is computed from60,000images across eight di?erent worms. The inhomogeneity along the diagonal shows that the nor-mal modes of the motion are not sinusoidal but the smooth structure of C(s,s′)means that a small number of modes are signi?cant.(b)We?nd the eigenvalues of C(s,s′)and computeσ2K,the fraction of the total variance captured by keeping just K modes(see Methods);95%of the variance is accounted for by the?rst four modes.(c)Associated with each of the dominant modes is an eigenvector describing an-gle vs.position along the worm;we refer to these as the eigenworms uμ(s).The population-mean eigenworms(red) are highly reproducible across individual worms(black).(d) In response to strong thermal stimuli the worm’s shape re-mains within the four dimensional subspace identi?ed from analysis of spontaneous crawling.Worm images are recorded at times synchronized to a heat pulse(see Methods)and we displayσ2K aligned with the stimulus.Though the shapes vary systematically in response to these stimuli,the accu-racy of our reconstructions does not and using all four modes we continue to account for95%of the shape variance. delivered intense pulses of heat(see Methods),which are known to trigger escape responses[14].We see in Fig2d that we still account for~95%of the shape variance using just four modes,even though the distribution of shapes during the thermal response is very di?erent from that seen in spontaneous crawling.We conclude that our four eigenworms provide an e?ective,low dimensional coordinate system within which to describe C.elegans motor behavior.
3
FIG.3:Motions along the?rst two eigenworms.(a)The joint probability density of the?rst two amplitudes,ρ(a1,a2), with units such that a21 = a22 =1.The ring structure sug-gests that these modes form an oscillator with approximately ?xed amplitude and varying phaseφ=tan?1(?a2/a1).(b) Images of worms with di?erent values ofφshow that variation in phase corresponds to propagating a wave of bending along the worms body.(c)Dynamics of the phaseφ(t)shows long periods of linear growth,corresponding to a steady rotation in the{a1,a2}plane,with occasional,abrupt reversals.(d) The joint densityρ(|v|,|ω|).The phase velocityω=dφ/dt in shape space predicts the speed at which the worm crawls on the agar.The crawling speed was de?ned as the time derivative of the worm’s center of mass.
WHAT DO THE MODES MEAN?
The projection of worm shapes onto the low-dimensional space of eigenworms provides a new and quantitative foundation for the
classical,qualitative de-scriptions of C.elegans behavior[8].The?rst two modes are sinuous(although not exactly sinusoidal)oscillations of the body shape(Fig2c);they form a quadrature pair,so that di?erent mixtures of the two modes cor-respond to di?erent phases of a wave along the body. Indeed,the probability distribution of the mode ampli-tudes,ρ(a1,a2),shows a ring of nearly constant ampli-tude(Fig3a).Sampling images around this ring reveals a traveling wave along the body(Fig3b).There are relatively long periods of time where the shape changes by a continuous accumulation of the phase angle(Fig 3c),and the speed of this rotation predicts the speed at which the worm crawls(Fig3d).
In contrast to the?rst two modes,the third mode u3(s)contributes to a nearly constant curvature throughout the middle half of the body(Fig2c).The distribution of the mode amplitude a3has a long tail (Fig4a),and body shapes chosen from these tails(Fig 4b)exhibit the?con?guration classically identi?ed with turning behavior[8].Large amplitudes of a3also corre-spond to regions of high curvature in the worm trajectory along the agar(Fig4c).
The connections between mode amplitudes and the motion of the worm along the agar—as in Figs3d and 4c—are genuine tests of the functional meaning of our low dimensional description.Quite explicitly,our anal-ysis of worm shapes is independent of the extrinsic co-ordinates and hence our de?nition of modes and ampli-tudes is blind to the actual position and orientation of the worm.Of course,in order to move the worm must change shape,and our description of the shape in terms of mode amplitudes captures this connection to move-FIG.4:Motions along the third eigenworm.(a)The dis-tribution of amplitudesρ(a3),shown on a logarithmic scale. Units are such that a23 =1,and for comparison we show the Gaussian distribution;note the longer tails inρ(a3).(b)Im-ages of worms with values of a3in the negative tail(left),the middle(center)and positive tail(right).Large amplitudes of a3correspond to?-like shapes.(c)A two minute trajectory of the center of mass sampled at4Hz.Periods where|a3|>1 are colored red,illustrating the association between turning and large displacements along this mode.
4
ment.Thus,to crawl smoothly forward or backward the worm changes its shape by rotating clockwise or counter-clockwise in the plane formed by the mode amplitudes a1and a2;the speed of crawling is set by the speed of the rotation.Similarly,to change direction the worm changes shape toward larger magnitudes of the mode amplitude a3,and we see this connection even without de?ning discrete turning events.
ATTRACTORS AND BEHA VIORAL STATES The eigenworms provide a coordinate system for the postures adopted by C.elegans as it moves;to describe the dynamics of movement we need to?nd equations of motion in this low dimensional space.We start by focus-ing on the plane
formed by the?rst two mode amplitudes a1and a2.Figure3suggests that within this plane the system stays at nearly constant values of the radius,so
that the relevant dynamics involves just the phase angle φ(t).To account for unobserved and random in?uences these equations need to be stochastic,and to support both forward and backward motion they need to form a system of at least second order.Such a system of equa-tions would be analogous to the description of Brownian motion using the Langevin equation[15,16].Thus we search for equations of the form
dφ(t)
dt
=F[φ(t),ω(t)]+σ[φ(t),ω(t)]η(t).(4) Here F[φ(t),ω(t)]de?nes the average acceleration as a function of the phase and phase velocity,by analogy to the force on a Brownian particle.The noise is charac-terized by a random functionη(t)which we hope will have a short correlation time,and we allow the strength of the noiseσ[φ(t),ω(t)]to depend on the state of the system,by analogy to a temperature that depends on the position of the Brownian particle.
In Figure5a we show our best estimate of the mean acceleration F[φ,ω](see Methods for details).Once we know F,we subtract this mean acceleration from the instantaneous acceleration to recover trajectories of the noise,and the correlation function of this noise is shown in Fig5b.The correlation time of the noise is short, which means that we have successfully separated the dy-namics into two parts:a deterministic part,described by the function F[φ,ω],which captures the average motion in the{a1,a2}plane and hence the relatively long peri-ods of constant oscillation,and a rapidly?uctuating part η(t)that describes“jittering”around this simple oscil-lation as well as the random forces that lead to jumps from one mode of motion to another.
We can imagine a hypothetical worm which has the same deterministic dynamics as we have found for real FIG.5:Reconstructing the phase dynamics.(a)The mean acceleration of the phase as a function of the instantaneous phase and phase velocity,F(ω,φ)in Eq(5).(b)The correla-tion function of the noise in the phase dynamics, η(t)η(t+τ) withη(t)de?ned from Eq(5).The correlations are con?ned to very short times,especially when compared with the cor-relations of the phase velocity itself.(c)Trajectories in the deterministic dynamics.A selection of early-time trajectories are shown in black.At late times these same trajectories col-lapse to one of four attractors(red):forward and backward crawling and two pause states.(d)Attracting basins.Ini-tial conditions covering the{φ,ω}plane were evolved to long times and each initial phase point was colored corresponding to its asymptotic attractor.Note that the basins are asym-metric so that some forward motions fall into the reversal or pause attractor.This highlights the di?erence between a dy-namical de?nition of behavioral state and phenomenological one based on an instantaneous value such as sgn(ω(t)). worms,but no noise.We can start such a noiseless worm at any combination of phase and phase velocity,and fol-low the dynamics predicted by Eqs(4),but withσ=0. The results are shown in Fig5c.The dynamics are di-verse on short time scales,depending in detail on the initial conditions,but eventually all initial conditions lead to one of a small number of possibilities(Fig5d): either the phase velocity is always positive,always neg-ative,or decays to zero as the system pauses at one of two stationary phases.Thus,underneath the continu-ous,stochastic dynamics we?nd four discrete attractors which correspond to well de?ned classes of behavior.
PAUSE STATES AND REPRODUCIBILITY The behavior of C.elegans,particularly in response to sensory stimuli,traditionally has been characterized in probabilistic terms:worms respond by changing the
5
probability of turning or reversing
[17,18,19].This
randomness could re?ect an active strategy on the part of the organism,or it could re?ect the inability of the nervous system to distinguish reliably between genuine sensory inputs and the inevitable background of noise. Our ability to describe motor behavior more completely, with high time resolution,o?ers us the opportunity to revisit the“psychophysics”of C.elegans.
We consider the response to brief(75ms),small (?T~0.1?C)changes in temperature,induced by pulses from an infrared laser(see Methods).These stimuli are large enough to elicit responses[10]but well below the threshold for pain avoidance[14].In Figure6we show the distributionρt(ω)of phase velocities as a function of time relative to the thermal pulse.All of the worms were crawling forward at the moment of stimulation,so the initial phase velocities are distributed over a wide range of positive values.Within one second,the dis-tribution narrows dramatically,concentrating near zero phase velocity-the pause states described above. Arrival in the pause state is stereotyped both across
trials and across worms.By analogy with conventional psychophysical methods[1],we can ask how reliably an observer could infer the presence of the heat pulse using the worm’s response.We?nd that just measuring the phase velocityωat single moment in time after the pulse is su?cient to provide~75%correct detection of this small temperature change in single trials.
COUPLING THE MODES AND STEERING THE
WORM
Our discussion thus far has separated the dynamics of the worm into two very di?erent components:the {a1,a2}plane with its phase dynamics,responsible for crawling motions,and the mode a3which is connected with large curvature turns.Because we found these modes as eigenvectors of a covariance matrix,we are guaranteed that the instantaneous amplitudes of the modes are not linearly correlated,but this does not mean that the dynamics of the di?erent motions are completely uncoupled.We found the clearest indications of mode coupling between the phase in the{a1,a2}plane and the amplitude a3at later times,which is illustrated by the correlation function in Fig6b.Intuitively,these correla-tions make sense,since the direction in which the worm forms its?shape must depend on the phase of the wave passing along the worm’s body at the moment that the large amplitude bending is initiated.This pattern of correlations is perturbed strongly by thermal stimuli. The fact that the correlations between phase and the turning mode are stimulus dependent implies that the re-sponse of the turning mode to thermal stimuli depends on the phase which the worm?nds itself at the time of the stimulus.Perhaps some of the apparent random-FIG.6:Thermal responses,mode coupling and active steer-ing(see Methods for details).(a)The distribution of phase velocitiesρt(ω)in response to a brief thermal stimulus.These data are taken from a collection of13worms,each stimu-lated with20repetitions of a?T=0.1?C pulse.Note that within one second,the distribution becomes highly concen-trated nearω=0,corresponding to the pause states identi-?ed in Fig5.In this experiment only,the worms were raised at a lower temperature leading to a lower averageωbefore the thermal stimulus.(b)Correlations between phase in the {a1,a2}plane and a3,C(t,t′)= δa3(t)δsin(φ(t′)) .Shortly after the thermal impulse,the modes develop a strong anti-correlation which is distinct from normal crawling(c)Worm ‘steering’.A thermal impulse conditioned on the instan-taneous phase was delivered automatically and repeatedly, causing an orientation change˙Θin the worm’s trajectory.In this example lasting4minutes,asynchronous impulses pro-duced a time-averaged orientation change ˙Θ =0.01rad/s (black),impulses at positive phase produced a trajectory with ˙Θ =0.10rad/s(blue),and impulses at negative phase produced ˙Θ =?0.12rad/s(red).This trajectory response is consistent with the early-time mode correlations seen in Fig 6b.We found13out of20worms produced statistically di?er-ent orientation changes under stimulated and non-simulated conditions while only1out of20worms responded in the same fashion when the phase was randomized.The asterisk marks the start of the trajectories.
ness of turning responses thus is related to the fact that repeated thermal stimuli catch the worm at di?erent ini-tial phases.As a preliminary test of this idea,we imple-mented our analysis online,allowing an estimate of the phase with a delay of less than125ms.We then deliver an infrared pulse when the phase falls within a phase window that corresponds to either dorsal–or ventral–directed head swings.To understand the predicted con-sequences of these pulses,consider an idealized example
6
in which the target phase isπ/2;then the correlation structure in Fig6b predicts a negative bias in a3which should generate right hand turns.Similarly,if the tar-get phase is?π/2,then continued thermal stimulation should generate left hand turns.Note that the thermal pulse itself does not have a handedness,so that if the pulses are not synchronized to the state of the worm there should be no systematic preference for right vs. left handed turns.These predictions are con?rmed in Fig6c.
DISCUSSION
Our central result is that C.elegans motor behavior is vastly simpler than it could be,even when the or-ganism moves freely.Conceptually similar results have been obtained for aspects of motor control in humans and other primates,where postures or trajectories of limbs,hands or eyes are con?ned to spaces of low dimen-sionality despite the potential for more complex motions [20,21,22,23].For C.elegans itself,recent quantitative work has focused on simplifying behavior by matching to a discrete set of template behaviors,such as forward and backward motion of the center of mass[5],sinusoidal undulations of the body[6],or?bends[7].Our results combine and generalize these ideas.Motor behaviors are described by projection of the body shape onto a small set of templates(the eigenworms),but the strengths of these projections vary continuously.The templates are sinuous,but not sinusoidal,because the?uctuations in posture are not homogeneous along the length of the worm.Our description of shape is intrinsic to the worm and invariant to the center of mass position,but mo-tion in shape space predicts the center of mass motion. There are discrete behavioral states,but these emerge as attractors of the underlying dynamics.Most impor-tantly,our choice of four eigenworms is driven not by hypotheses about the relevant components of behavior, but by the data itself.
The construction of the eigenworms guarantees that the instantaneous amplitudes along the di?erent dimen-sions of shape space are not correlated linearly,but the dynamics of the di?erent amplitudes are nonlinear and coupled;what we think of as a single motor action always involves coordinating multiple degrees of freedom.Thus, forward and backward motion correspond to positive and negative phase velocity in Fig3,but transitions between these behavioral states occur preferentially at particu-lar phases(data not shown).Similarly,?turns involve a large amplitude excursion along a3,but motion along this mode is correlated with phase in the({a1,a2})plane, and this correlation itself has structure in time(Fig6b). The problems of C.elegans motor control are simpler than for higher animals,but these nonlinear,coupled dynamics give a glimpse of the more general case. Perhaps because of the strong coupling between the turning mode a3and the wriggling modes a1,a2,we have not found an equation of motion for a3alone which would be analogous to Eq’s(4)for the phase.Further work is required to construct a fully three dimensional dynamics which could predict the more complex correlations such as those in Fig6b.Turning should emerge from these equations not as another attractor,but as an‘excitable’orbit analogous to the action potential in the Hodgkin–Huxley equations or to recent ideas about transient dif-ferentiation in genetic circuits[24].A major challenge would be to show that the stochastic dynamics of these equations can generate longer sequences of stereotyped events,such as pirouettes[25].
We have shown that a meaningful set of behavioral coordinates can uncover deterministic responses.A re-sponse might seem stochastic or noisy because it depends on one or more behavioral variables that are not being considered.In our experiments,nonlinear correlations among the behavioral variables suggest that some of the randomness in behavioral responses could be removed if sensory stimuli are delivered only when the worm is at a well de?ned initial state,and we con?rmed this pre-diction by showing that phase–aligned thermal stimuli can‘steer’the worm into trajectories with a de?nite chi-rality.A crucial aspect of these experiments is that the stimulus is scalar—a temperature change in time has no spatial direction or handedness—but the response, by virtue of the correlation between stimulus and body shape,does have a de?nite spatial structure.The align-ment of thermal stimuli with the phase of the worm’s movement in these experiments mimics the correlation between body shape and sensory input that occurs as the worm crawls in a thermal gradient,so the enhanced determinism of responses under these conditions may be connected to the computations which generate nearly deterministic isothermal tracking[19,26].
More generally,all behavioral responses have some mixture of deterministic and stochastic components.In humans and other primates,it seems straightforward to create conditions that result in highly reproducible, stereotyped behaviors,such as reaching movements[27]. At the opposite extreme,bacterial motility is modulated in response to sensory inputs,but these responses seem fundamentally probabilistic[28].Some of these di?er-ences may result from the physical nature of sensory stimuli in organisms of vastly di?erent size[29,30],but some of the di?erences may also result from di?erences of strategy or available computational power.The more stochastic the response,the more challenging it is to characterize behavior quantitatively and to link behav-ior with underlying molecular and neural components, as is clear from recent work on Drosophila olfaction(see, for example,Ref[31]).We hope that our approach to
7
the analysis of behavior may help to uncover more de-terministic components of the sensory–motor responses in other model organisms.
More than forty years of work on C.elegans has led to a fully sequenced genome[32]and to the complete wiring diagram of the nervous system[33].Signi?cant steps have been made toward the original dream[34] of connecting genes,neurons,and behavior[9,35,36]. Nonetheless,with the advances in molecular,cellular, and circuit analyses,our ability to probe the mechanisms which generate behavior substantially exceeds our ability to characterize the behavior itself.Perhaps our work provides a step toward addressing this imbalance.
METHODS
Tracking microscopy.The imaging system con-sists of a Basler?rewire CMOS camera(A601f,Basler, Ahrensburg,Germany)with4x lens(55-901,Ed-mund Optics,Barrington,NJ)and a?ber optic trans-illuminator(DC-950,Dolan-Jenner,Boxborough,MA) mounted to an optical rail(Thorlabs,Newton,NJ). The rail is attached to a XY translation stage(Deltron, Bethel,CT)which is driven by stepper motors(US Digi-tal,Vancouver,Washington).The stage driver is a home made unit utilizing a SimpleStep board(SimpleStep, Newton,NJ)and Gecko stepper motor drivers(Geck-odrive,Santa Ana,CA).Image acquisition,processing, and stage driver control was done using LabVIEW(Na-tional Instruments,Austin,TX).Images of worms were isolated and identi?ed using the image particle?lter.A raw unprocessed JPEG image and a?ltered process bi-nary PNG image were written to the hard drive at rates up to32Hz.Concurrently at4Hz,the center of mass of the worm was calculated and the distance from the center of the?eld of view in pixels was computed.An error signal was then calculated via a coordinate trans-formation between the camera reference frame and the translational stage reference frame and the XY stage was moved to center the worm in the?eld of view.
Worm preparation.The C.elegans strain,N2, was grown at20?C and maintained under standard con-ditions[37].Before each experiment,excess moisture from NGM assay plates(1.7%Bacto Agar,0.25%Bacto-Peptone,0.3%NaCl,1mM CaCl2,1mM MgSO4,25 mM potassium phosphate bu?er,5μg/mL cholesterol) was removed by leaving them partially uncovered for1 hr.A copper ring(5.1-cm inner diameter)pressed into the agar surface prevented worms from crawling to the side of the plate.Young adults were rinsed of E.coli by transferring them with a worm pick from OP50bacterial food plates into NGM bu?er(same inorganic ion concen-tration as NGM assay plates)and letting them swim for 1minute.Worms were transferred from the NGM bu?er to the center of the assay plate(9-cm Petri dish).The
plates were covered and tracking began after1minute
and lasted no longer than60minutes.In the rare cases where worms stopped moving before the completion of
the run,the data were excluded.
Eigenworms.Images of worms captured by the
worm tracker were processed using MATLAB(Math-works,Natick,MA).Cases of self-intersection were ex-
cluded from processing.Images of worms were thinned
to a single-pixel-thick backbone.A spline was?t through these points and then discretized into101segments,
evenly spaced in units of the backbone arclength.The
N=100angles between these segments were calcu-lated and an overall rotation mode was removed by
subtracting θ(s(i))/N from each angle.The shape covariance matrix C(s,s′)= (θ(s)? θ )(θ(s′)? θ )
was constructed from9freely crawling worms,sam-
pled at4Hz,for a period of30minutes.Each eigen-
worm uμ(s)is an eigenvector of the covariance matrix s′C(s,s′)uμ(s′)=λμuμ(s).The fractional variance captured by K eigenvectors is thusσ2K= Kμ=1λμ/σ2, whereσ2= μλμis the total variance of the measure-ments.The same eigenworms shown in Fig.2were used throughout the various analysis reported in the paper. The worm’s phase was de?ned asφ=tan?1(?a2/a1) where a1and a2were both normalized to unit variance. Equations of motion.For the analysis of phase dy-namics we sampled the worm shape at32Hz.Data for the construction of the equations of motion came from 12worms,5trials per worm,with4000frames per trial. We also?ltered each mode time series through a low-pass polynomial?lter so that for each frame(26≤m≤3974),?a(m)= 25n=?25 4j=0p j(m?n)j where{p j}are the best-?t polynomial coe?cients.Mode time derivatives were calculated using derivatives of the polynomial?lter. None of our results depend critically on the properties of the?lter.The Langevin equations governing the phase dynamics are shown Eq.(4)and we learn the functions {F(φ,ω),σ(φ,ω)}directly from the time series[38,39]. By construction σ[φ(t),ω(t)]η(t) =0and therefore the optimal rms estimate of F(φ,ω)is the conditional mean ˙ω|ω,φ .We estimate F by assuming a functional expansion F(ω,φ)= 5m=?5 5p=0αp m w k e?imφ,where the model parameters{αp m}were determined by mini-mizing the rms error?2= t(˙ω(t)?F[ω(t),φ(t)])2on training data(90%)and the hyperparameters{m max= 5,p max=5}were chosen to minimize error on held-out data(10%).Once F is known we can determine the noise in the system;we normalize η2 so that σ2(ω,φ)= (˙ω?F(ω,φ))2|ω,φ .The attractors con-tained within our derived dynamics were obtained by evolving initial conditions spanning the sampled{ω,φ} plane for long times(93.75s~47cycles).In the deter-ministic dynamics all trajectories evolve to one of four asymptotic states and we observed no switching.
8
Thermal impulse response(experiment). Worms were prepared as described earlier.A collimated beam with a1/e diameter of5.6mm(standard stim-
ulus)or1.5mm(painful)from a1440nm diode laser (FOL1404QQM,Fitel,Peachtree City,GA)was posi-tioned to heat the area covering the worm.The diode
laser was driven with a commercial power supply and controller(Thorlabs,Newton,NJ).Power and duration
of the beam was controlled through software using LabVIEW.For each worm,1000seconds of data was collected in cycles of50seconds.12.5seconds into each
cycle the laser was turned on for a duration of75ms at150mW(standard)or250ms at100mW(painful). The temperature increase caused by the laser pulses
was measured using a0.075mm T-type thermocouple (coco-003,Omega,Stamford,CT)placed on the surface
of the agar and sampled with a thermocouple data acquisition device(USB-9211,National Instruments). For each measurement,60trials of30s cycles were
averaged.The temperature increase was calculated by subtracting the maximum temperature(recorded immediately after the laser pulse)from the baseline
temperature(recorded9s after the laser pulse).The temperature increase for the standard pulse was0.12?C
and the increase of the painful pulse was0.73?C. Thermal impulse response(analysis).In Fig 6a,the time-dependent probability densityρt(ω)was
smoothed before the onset of the impulse with a gaus-sian low-pass?lter of size0.19s in the t direction and
0.17cycles/s in theωdirection.In Fig6b the correlation function C(t,t′)= (sinφ(t)? sinφ )(a3(t′)? a3 ) was calculated as follows.Far from the time of the
impulse(frames800to1574,impulse on frame400), we expect time-translation invariance C post(t,t′)= g(t?t′)+ξpost(t,t′)where g(?)= C(i,j) i?j=?
is the true correlation function andξpost(t,t′)charac-terizes statistical error.Similarly in a time window
around the impulse(frames24to800),C stim(t,t′)= g(t?t′)+ξstim(t,t′).However,the thermal im-pulse breaks this invariance andξstim(t,t′)contains
both sampling?uctuations and stimulus-dependent cor-relation dynamics.To separate these e?ects we use singular value decomposition to compareξpost
andξstim.We write each matrixξpost/stim(t,t′)= t′′U post/stim(t,t′′)S post/stim(t′′,t′′)V post/stim(t′′,t′)and ?nd that only two singular values ofξstim are signi?-cantly larger thenξpost.We then reconstruct the two-point function around the stimulus as?C stim(t,t′)= g(t?t′)+ 2t′′=1U stim(t,t′′)S stim(t′′,t′′)V stim(t′′,t′). Thermal steering.Preparation of worms and in-strumentation were the same as described for the ther-mal impulse response.However,instead of process-
ing worm images o?-line,real-time calculation of the eigenworms and shape phaseφwas done using custom dynamic-linked image processing libraries written in C along with supporting LabVIEW code.The modes were computed as previously described except that the spline interpolation algorithm was replaced with a Hermitian interpolation algorithm to reduce the processing time. The processing time was short enough to simultaneously track and calculate modes at8Hz.For phase dependent measurements,the laser was?red when the worm was moving forward andφfell within a prescribed interval (width1radian).The laser pulse(150mW)lasted for 75ms and caused a temperature increase of0.12?C.For each run a pair of triggering phase windows(0to-1,and 2.1and3.1radians)corresponding to the dorsal-and ventral-directed head swing was used.The sequence of each run started with a5minute period of no stimulus followed by the pair of phase dependent stimuli.The or-der of each pair of stimulus conditions was switched for each successive run.For the randomized pulse control experiments,the laser was?red with a uniform phase probability,but with conditions that restricted the?r-ing interval to be longer than2seconds.
Steering and turn identi?cation.The time-average change in orientation of the worms path, ˙Θ (rad/s),was calculated from the angular changes be-tween the positions of the center of mass of the worm during forward runs of at least4s in length.Given posi-tions(r1,r2,r3,...,r N),the angles between connecting segments(r2?r1,r3?r2,...,r N?r(N?1))were calcu-lated. ˙Θ was calculated in intervals of10s.Since the distributions were Gaussian(data not shown)with sim-ilar variance,we used the Student’s t-test to determine if the values of˙Θunder thermal stimulation were sig-ni?cantly di?erent than the control(p<0.05).Since we were interested in the change in orientation dur-ing forward motion we excluded trajectory data that contained large turns or reversals along with angular changes greater thanπ/4radians.These events were automatically detected by measuring the compactness of the worm https://www.360docs.net/doc/4818510844.html,pactness was calculated by mea-suring the longest distance between two points in the worm shape(also known as the max feret distance)and normalizing this with the maximum value for the entire data run.Turns were?agged when the compactness fell below0.6.
[1]DM Green&JA Swets,Signal Detection Theory and
Psychophysics(John Wiley&Sons,New York1966). [2]J Bolhuis&L Giraldeau,Editors,The Behavior of An-
imals:Mechanism,Function and Evolution(Blackwell Publishing,Oxford,2004).
[3]W Geng,P Cosman,JH Baek,C Berry&WR Schafer,
Quantitative classi?cation and natural clustering of Caenorhabditis elegans behavioral phenotypes.Genetics 165,1117–1126(2003).
[4]CJ Cronin,JE Mendel,S Mukhtar,Y Kim,RC Stirbl,J
9
Bruck&PW Sternberg,An automated system for mea-suring parameters of nematode sinusoidal movement.
BMC Genet6,5–24(2005).
[5]K Hoshi&R Shingai,Computer–driven automatic iden-
ti?cation of locomotion states in Caenorhabditis elegans.
J Neurosci Meth157,355–363(2006).
[6]J Karbowski,CJ Cronin,A Seah,JE Mendel,D Cleary
&PW Sternberg,Conservation rules,their breakdown, and optimality in Caenorhabditis sinusoidal locomotion.
J Theor Biol242,652–669(2006).
[7]KM Huang,PC Cosman&W Schafer,Machine vision
based detection of omega bends and reversals in C.ele-gans.J Neurosci Meth158,323–336(2006).
[8]N Croll,Components and patterns in the behavior of the
nematode Caenorhabditis elegans.J Zool176,159–176 (1975).
[9]CI Bargmann,Genetic and cellular analysis of behavior
in C.elegans.Annu Rev Neurosci16,47–71(1993). [10]EM Hedgecock&RL Russell,Normal and mutant ther-
motaxis in the nematode Caenorhabditis elegans.Proc Nat’l Acad Sci(USA)72,4061–4065(1975).
[11]M Chal?e,JE Sulston,JG White,E Southgate,JN
Thomson&S Brenner,The neural circuit for touch sen-sitivity in Caenorhabditis elegans.J Neurosci5,956–964 (1985).
[12]J Frenet,Sur quelque properiet`e s des courbes′a double
courbure.Th`e se,Toulouse,1847.Abstract in Jour de Math17,437(1852).
[13]DJ Struik,Lectures on Classical Di?erential Geometry
(Addison-Wesley,Reading,1961).
[14]N Wittenburg&R Baumeister,Thermal avoidance in
Caenorhabditis elegans:An approach to the study of nociception.Proc Nat’l Acad Sci(USA)96,10477–10482 (1999).
[15]NG van Kampen,Stochastic Processes in Physics and
Chemistry,2nd Edition(North Holland,Amsterdam, 2001).
[16]D Selmecai,S Mosler,PH Hagedorn,NB Larsen&
H Flyvbjerg,Cell motility as persistent random mo-
tion:Theories from experiment.Biophys J89,912–931 (2005).
[17]JM Gray,JJ Hill&CI Bargmann,A circuit for nav-
igation in Caenorhabditis elegans.Proc Nat’l Acad Sci (USA)102,3184–3191(2005).
[18]HA Zariwala,AC Miller,S Faumont&SR Lockery,Step
response analysis of thermotaxis in Caenorhabditis ele-gans.J Neurosci23,4369–4377(2003).
[19]WS Ryu&AD Samuel,Thermotaxis in Caenorhabdi-
tis elegans analyzed by measuring responses to de?ned thermal stimuli.J Neurosci22,5727–5733(2002). [20]A d’Avella&E Bizzi,Low dimensionality of sur-
paspinally induced force?elds.Proc Nat’l Acad Sci (USA)95,7711–7714(1998).
[21]M Santello,M Flanders&JF Soechting,Postural hand
strategies for tool use.J Neurosci18,10105–10115 (1998).
[22]TD Sanger,Human arm movements described by a low-
dimensional superposition of principal components.J Neurosci20,1066–1072(2000).
[23]LC Osborne,SG Lisberger&W Bialek,A sensory source
for motor variation.Nature437,412–416(2005). [24]GM Suel,J Garcia–Ojalvo,LM Liberman&MB Elowitz,
An excitable gene regulatory circuit induces transient cellular di?erentiation.Nature440,545–550(2006). [25]JT Pierce–Shimomura,TM Morse&SR Lockery,The
fundamental role of pirouettes in Caenorhabditis elegans chemotaxis.J Neurosci19,9557–9569(1999).
[26]L Luo,DA Clark,D Biron,L Mahadevan&AD
Samuel,Sensorimotor control during isothermal track-ing in Caenorhabditis elegans.J Exp Biol209,4652–4662(2006).
[27]M Jeannerod,The Neural and Behavioural Organization
of Goal-Directed Movements(Oxford Univ Press,Ox-ford,1988).
[28]HC Berg,E.coli in Motion(Springer–Verlag,New York,
2004).
[29]HC Berg&EM Purcell,Physics of chemoreception.Bio-
phys J20,193–219(1977).
[30]W Bialek,Physical limits to sensation and perception.
Annu Rev Biophys Biophys Chem16,455–478(1987).
[31]A Keller&LB Vosshall,ln?uence of odorant receptor
repertoire on odor perception in humans and fruit?ies.
Proc Nat’l Acad Sci(USA)104,5614–5619(2007). [32]The C.elegans Sequencing Consortium,Genome se-
quence of the nematode C.elegans:a platform for in-vestigating biology.Science282,2012–2018(1998). [33]JG White,E Southgate,JN Thomson&S Brenner,
The structure of the nervous system of the nematode Caenorhabditis elegans.Phil Trans R Soc B314,1–340 (1986).
[34]S Brenner,The genetics of Caenorhabditis elegans.Ge-
netics77,71–94(1974).
[35]M de Bono&AV Maricq,Neuronal substrates of com-
plex behaviors in C.elegans.Annu Rev Neurosci28, 451–501(2005).
[36]S Faumont&SR Lockery,The awake behaving worm:
simultaneous imaging of neuronal activity and behavior in intact animals at millimeter scale.J Neurophys95, 1976–1981(2006).
[37]JE Sulston&S Brenner,The DNA of Caenorhabditis
elegans.Genetics77,95–104(1974).
[38]R Friedrich,J Peinke&Ch Renner,How to quantify
deterministic and random in?uences on the statistics of the foreign exchange market.Phys Rev Lett84,5224–5227(2000).
[39]E Racca&A Porporato,Langevin equations from time
series.Phys Rev E71,027101–027103(1999).
We thank D Chigirev,SE Palmer,E Schneidman and G Tkaˇc ik for discussions and AR Chapman for help in the ini-tial building of the worm tracker and for programming the thinning algorithm used for real–time processing.This work was supported in part by Los Alamos National Laboratory, by a training grant from the National Institute of Mental Health,by NIH Grant P50GM071508and EY017210,and by the Swartz Foundation.WB also thanks LF Abbott, KD Miller,and the Center for Theoretical Neuroscience at Columbia University for their hospitality.
The way常见用法
The way 的用法 Ⅰ常见用法: 1)the way+ that 2)the way + in which(最为正式的用法) 3)the way + 省略(最为自然的用法) 举例:I like the way in which he talks. I like the way that he talks. I like the way he talks. Ⅱ习惯用法: 在当代美国英语中,the way用作为副词的对格,“the way+ 从句”实际上相当于一个状语从句来修饰整个句子。 1)The way =as I am talking to you just the way I’d talk to my own child. He did not do it the way his friends did. Most fruits are naturally sweet and we can eat them just the way they are—all we have to do is to clean and peel them. 2)The way= according to the way/ judging from the way The way you answer the question, you are an excellent student. The way most people look at you, you’d think trash man is a monster. 3)The way =how/ how much No one can imagine the way he missed her. 4)The way =because
The way的用法及其含义(二)
The way的用法及其含义(二) 二、the way在句中的语法作用 the way在句中可以作主语、宾语或表语: 1.作主语 The way you are doing it is completely crazy.你这个干法简直发疯。 The way she puts on that accent really irritates me. 她故意操那种口音的样子实在令我恼火。The way she behaved towards him was utterly ruthless. 她对待他真是无情至极。 Words are important, but the way a person stands, folds his or her arms or moves his or her hands can also give us information about his or her feelings. 言语固然重要,但人的站姿,抱臂的方式和手势也回告诉我们他(她)的情感。 2.作宾语 I hate the way she stared at me.我讨厌她盯我看的样子。 We like the way that her hair hangs down.我们喜欢她的头发笔直地垂下来。 You could tell she was foreign by the way she was dressed. 从她的穿著就可以看出她是外国人。 She could not hide her amusement at the way he was dancing. 她见他跳舞的姿势,忍俊不禁。 3.作表语 This is the way the accident happened.这就是事故如何发生的。 Believe it or not, that's the way it is. 信不信由你, 反正事情就是这样。 That's the way I look at it, too. 我也是这么想。 That was the way minority nationalities were treated in old China. 那就是少数民族在旧中
(完整版)the的用法
定冠词the的用法: 定冠词the与指示代词this ,that同源,有“那(这)个”的意思,但较弱,可以和一个名词连用,来表示某个或某些特定的人或东西. (1)特指双方都明白的人或物 Take the medicine.把药吃了. (2)上文提到过的人或事 He bought a house.他买了幢房子. I've been to the house.我去过那幢房子. (3)指世界上独一无二的事物 the sun ,the sky ,the moon, the earth (4)单数名词连用表示一类事物 the dollar 美元 the fox 狐狸 或与形容词或分词连用,表示一类人 the rich 富人 the living 生者 (5)用在序数词和形容词最高级,及形容词等前面 Where do you live?你住在哪? I live on the second floor.我住在二楼. That's the very thing I've been looking for.那正是我要找的东西. (6)与复数名词连用,指整个群体 They are the teachers of this school.(指全体教师) They are teachers of this school.(指部分教师) (7)表示所有,相当于物主代词,用在表示身体部位的名词前 She caught me by the arm.她抓住了我的手臂. (8)用在某些有普通名词构成的国家名称,机关团体,阶级等专有名词前 the People's Republic of China 中华人民共和国 the United States 美国 (9)用在表示乐器的名词前 She plays the piano.她会弹钢琴. (10)用在姓氏的复数名词之前,表示一家人 the Greens 格林一家人(或格林夫妇) (11)用在惯用语中 in the day, in the morning... the day before yesterday, the next morning... in the sky... in the dark... in the end... on the whole, by the way...
“the way+从句”结构的意义及用法
“theway+从句”结构的意义及用法 首先让我们来看下面这个句子: Read the followingpassageand talkabout it wi th your classmates.Try totell whatyou think of Tom and ofthe way the childrentreated him. 在这个句子中,the way是先行词,后面是省略了关系副词that或in which的定语从句。 下面我们将叙述“the way+从句”结构的用法。 1.the way之后,引导定语从句的关系词是that而不是how,因此,<<现代英语惯用法词典>>中所给出的下面两个句子是错误的:This is thewayhowithappened. This is the way how he always treats me. 2.在正式语体中,that可被in which所代替;在非正式语体中,that则往往省略。由此我们得到theway后接定语从句时的三种模式:1) the way+that-从句2)the way +in which-从句3) the way +从句 例如:The way(in which ,that) thesecomrade slookatproblems is wrong.这些同志看问题的方法
不对。 Theway(that ,in which)you’re doingit is comple tely crazy.你这么个干法,简直发疯。 Weadmired him for theway inwhich he facesdifficulties. Wallace and Darwingreed on the way inwhi ch different forms of life had begun.华莱士和达尔文对不同类型的生物是如何起源的持相同的观点。 This is the way(that) hedid it. I likedthe way(that) sheorganized the meeting. 3.theway(that)有时可以与how(作“如何”解)通用。例如: That’s the way(that) shespoke. = That’s how shespoke.
way 用法
表示“方式”、“方法”,注意以下用法: 1.表示用某种方法或按某种方式,通常用介词in(此介词有时可省略)。如: Do it (in) your own way. 按你自己的方法做吧。 Please do not talk (in) that way. 请不要那样说。 2.表示做某事的方式或方法,其后可接不定式或of doing sth。 如: It’s the best way of studying [to study] English. 这是学习英语的最好方法。 There are different ways to do [of doing] it. 做这事有不同的办法。 3.其后通常可直接跟一个定语从句(不用任何引导词),也可跟由that 或in which 引导的定语从句,但是其后的从句不能由how 来引导。如: 我不喜欢他说话的态度。 正:I don’t like the way he spoke. 正:I don’t like the way that he spoke. 正:I don’t like the way in which he spoke. 误:I don’t like the way how he spoke. 4.注意以下各句the way 的用法: That’s the way (=how) he spoke. 那就是他说话的方式。 Nobody else loves you the way(=as) I do. 没有人像我这样爱你。 The way (=According as) you are studying now, you won’tmake much progress. 根据你现在学习情况来看,你不会有多大的进步。 2007年陕西省高考英语中有这样一道单项填空题: ——I think he is taking an active part insocial work. ——I agree with you_____. A、in a way B、on the way C、by the way D、in the way 此题答案选A。要想弄清为什么选A,而不选其他几项,则要弄清选项中含way的四个短语的不同意义和用法,下面我们就对此作一归纳和小结。 一、in a way的用法 表示:在一定程度上,从某方面说。如: In a way he was right.在某种程度上他是对的。注:in a way也可说成in one way。 二、on the way的用法 1、表示:即将来(去),就要来(去)。如: Spring is on the way.春天快到了。 I'd better be on my way soon.我最好还是快点儿走。 Radio forecasts said a sixth-grade wind was on the way.无线电预报说将有六级大风。 2、表示:在路上,在行进中。如: He stopped for breakfast on the way.他中途停下吃早点。 We had some good laughs on the way.我们在路上好好笑了一阵子。 3、表示:(婴儿)尚未出生。如: She has two children with another one on the way.她有两个孩子,现在还怀着一个。 She's got five children,and another one is on the way.她已经有5个孩子了,另一个又快生了。 三、by the way的用法
The way的用法及其含义(一)
The way的用法及其含义(一) 有这样一个句子:In 1770 the room was completed the way she wanted. 1770年,这间琥珀屋按照她的要求完成了。 the way在句中的语法作用是什么?其意义如何?在阅读时,学生经常会碰到一些含有the way 的句子,如:No one knows the way he invented the machine. He did not do the experiment the way his teacher told him.等等。他们对the way 的用法和含义比较模糊。在这几个句子中,the way之后的部分都是定语从句。第一句的意思是,“没人知道他是怎样发明这台机器的。”the way的意思相当于how;第二句的意思是,“他没有按照老师说的那样做实验。”the way 的意思相当于as。在In 1770 the room was completed the way she wanted.这句话中,the way也是as的含义。随着现代英语的发展,the way的用法已越来越普遍了。下面,我们从the way的语法作用和意义等方面做一考查和分析: 一、the way作先行词,后接定语从句 以下3种表达都是正确的。例如:“我喜欢她笑的样子。” 1. the way+ in which +从句 I like the way in which she smiles. 2. the way+ that +从句 I like the way that she smiles. 3. the way + 从句(省略了in which或that) I like the way she smiles. 又如:“火灾如何发生的,有好几种说法。” 1. There were several theories about the way in which the fire started. 2. There were several theories about the way that the fire started.
way 的用法
way 的用法 【语境展示】 1. Now I’ll show you how to do the experiment in a different way. 下面我来演示如何用一种不同的方法做这个实验。 2. The teacher had a strange way to make his classes lively and interesting. 这位老师有种奇怪的办法让他的课生动有趣。 3. Can you tell me the best way of working out this problem? 你能告诉我算出这道题的最好方法吗? 4. I don’t know the way (that / in which) he helped her out. 我不知道他用什么方法帮助她摆脱困境的。 5. The way (that / which) he talked about to solve the problem was difficult to understand. 他所谈到的解决这个问题的方法难以理解。 6. I don’t like the way that / which is being widely used for saving water. 我不喜欢这种正在被广泛使用的节水方法。 7. They did not do it the way we do now. 他们以前的做法和我们现在不一样。 【归纳总结】 ●way作“方法,方式”讲时,如表示“以……方式”,前面常加介词in。如例1; ●way作“方法,方式”讲时,其后可接不定式to do sth.,也可接of doing sth. 作定语,表示做某事的方法。如例2,例3;
the-way-的用法讲解学习
t h e-w a y-的用法
The way 的用法 "the way+从句"结构在英语教科书中出现的频率较高, the way 是先行词, 其后是定语从句.它有三种表达形式:1) the way+that 2)the way+ in which 3)the way + 从句(省略了that或in which),在通常情况下, 用in which 引导的定语从句最为正式,用that的次之,而省略了关系代词that 或 in which 的, 反而显得更自然,最为常用.如下面三句话所示,其意义相同. I like the way in which he talks. I like the way that he talks. I like the way he talks. 一.在当代美国英语中,the way用作为副词的对格,"the way+从句"实际上相当于一个状语从句来修饰全句. the way=as 1)I'm talking to you just the way I'd talk to a boy of my own. 我和你说话就象和自己孩子说话一样. 2)He did not do it the way his friend did. 他没有象他朋友那样去做此事. 3)Most fruits are naturally sweet and we can eat them just the way they are ----all we have to do is clean or peel them . 大部分水果天然甜润,可以直接食用,我们只需要把他们清洗一下或去皮.
way的用法总结大全
way的用法总结大全 way的用法你知道多少,今天给大家带来way的用法,希望能够帮助到大家,下面就和大家分享,来欣赏一下吧。 way的用法总结大全 way的意思 n. 道路,方法,方向,某方面 adv. 远远地,大大地 way用法 way可以用作名词 way的基本意思是“路,道,街,径”,一般用来指具体的“路,道路”,也可指通向某地的“方向”“路线”或做某事所采用的手段,即“方式,方法”。way还可指“习俗,作风”“距离”“附近,周围”“某方面”等。 way作“方法,方式,手段”解时,前面常加介词in。如果way前有this, that等限定词,介词可省略,但如果放在句首,介词则不可省略。
way作“方式,方法”解时,其后可接of v -ing或to- v 作定语,也可接定语从句,引导从句的关系代词或关系副词常可省略。 way用作名词的用法例句 I am on my way to the grocery store.我正在去杂货店的路上。 We lost the way in the dark.我们在黑夜中迷路了。 He asked me the way to London.他问我去伦敦的路。 way可以用作副词 way用作副词时意思是“远远地,大大地”,通常指在程度或距离上有一定的差距。 way back表示“很久以前”。 way用作副词的用法例句 It seems like Im always way too busy with work.我工作总是太忙了。 His ideas were way ahead of his time.他的思想远远超越了他那个时代。 She finished the race way ahead of the other runners.她第一个跑到终点,远远领先于其他选手。 way用法例句
the_way的用法大全教案资料
t h e_w a y的用法大全
The way 在the way+从句中, the way 是先行词, 其后是定语从句.它有三种表达形式:1) the way+that 2)the way+ in which 3)the way + 从句(省略了that或in which),在通常情况下, 用in which 引导的定语从句最为正式,用that的次之,而省略了关系代词that 或 in which 的, 反而显得更自然,最为常用.如下面三句话所示,其意义相同. I like the way in which he talks. I like the way that he talks. I like the way he talks. 如果怕弄混淆,下面的可以不看了 另外,在当代美国英语中,the way用作为副词的对格,"the way+从句"实际上相当于一个状语从句来修饰全句. the way=as 1)I'm talking to you just the way I'd talk to a boy of my own. 我和你说话就象和自己孩子说话一样. 2)He did not do it the way his friend did. 他没有象他朋友那样去做此事. 3)Most fruits are naturally sweet and we can eat them just the way they are ----all we have to do is clean or peel them . 大部分水果天然甜润,可以直接食用,我们只需要把他们清洗一下或去皮. the way=according to the way/judging from the way 4)The way you answer the qquestions, you must be an excellent student. 从你回答就知道,你是一个优秀的学生. 5)The way most people look at you, you'd think a trashman was a monster. 从大多数人看你的目光中,你就知道垃圾工在他们眼里是怪物. the way=how/how much 6)I know where you are from by the way you pronounce my name. 从你叫我名字的音调中,我知道你哪里人. 7)No one can imaine the way he misses her. 人们很想想象他是多么想念她. the way=because 8) No wonder that girls looks down upon me, the way you encourage her. 难怪那姑娘看不起我, 原来是你怂恿的
the way 的用法
The way 的用法 "the way+从句"结构在英语教科书中出现的频率较高, the way 是先行词, 其后是定语从句.它有三种表达形式:1) the way+that 2)the way+ in which 3)the way + 从句(省略了that或in which),在通常情况下, 用in which 引导的定语从句最为正式,用that的次之,而省略了关系代词that 或in which 的, 反而显得更自然,最为常用.如下面三句话所示,其意义相同. I like the way in which he talks. I like the way that he talks. I like the way he talks. 一.在当代美国英语中,the way用作为副词的对格,"the way+从句"实际上相当于一个状语从句来修饰全句. the way=as 1)I'm talking to you just the way I'd talk to a boy of my own. 我和你说话就象和自己孩子说话一样. 2)He did not do it the way his friend did. 他没有象他朋友那样去做此事. 3)Most fruits are naturally sweet and we can eat them just the way they are ----all we have to do is clean or peel them . 大部分水果天然甜润,可以直接食用,我们只需要把他们清洗一下或去皮.
the way=according to the way/judging from the way 4)The way you answer the qquestions, you must be an excellent student. 从你回答就知道,你是一个优秀的学生. 5)The way most people look at you, you'd think a trashman was a monster. 从大多数人看你的目光中,你就知道垃圾工在他们眼里是怪物. the way=how/how much 6)I know where you are from by the way you pronounce my name. 从你叫我名字的音调中,我知道你哪里人. 7)No one can imaine the way he misses her. 人们很想想象他是多么想念她. the way=because 8) No wonder that girls looks down upon me, the way you encourage her. 难怪那姑娘看不起我, 原来是你怂恿的 the way =while/when(表示对比) 9)From that day on, they walked into the classroom carrying defeat on their shoulders the way other students carried textbooks under their arms. 从那天起,其他同学是夹着书本来上课,而他们却带着"失败"的思想负担来上课.
The way的用法及其含义(三)
The way的用法及其含义(三) 三、the way的语义 1. the way=as(像) Please do it the way I’ve told you.请按照我告诉你的那样做。 I'm talking to you just the way I'd talk to a boy of my own.我和你说话就像和自己孩子说话一样。 Plant need water the way they need sun light. 植物需要水就像它们需要阳光一样。 2. the way=how(怎样,多么) No one can imagine the way he misses her.没人能够想象出他是多么想念她! I want to find out the way a volcano has formed.我想弄清楚火山是怎样形成的。 He was filled with anger at the way he had been treated.他因遭受如此待遇而怒火满腔。That’s the way she speaks.她就是那样讲话的。 3. the way=according as (根据) The way you answer the questions, you must be an excellent student.从你回答问题来看,你一定是名优秀的学生。 The way most people look at you, you'd think a trash man was a monster.从大多数人看你的目光中,你就知道垃圾工在他们眼里是怪物。 The way I look at it, it’s not what you do that matters so much.依我看,重要的并不是你做什么。 I might have been his son the way he talked.根据他说话的样子,好像我是他的儿子一样。One would think these men owned the earth the way they behave.他们这样行动,人家竟会以为他们是地球的主人。
way的用法
一.Way:“方式”、“方法” 1.表示用某种方法或按某种方式 Do it (in) your own way. Please do not talk (in) that way. 2.表示做某事的方式或方法 It’s the best way of studying [to study] English.。 There are different ways to do [of doing] it. 3.其后通常可直接跟一个定语从句(不用任何引导词),也可跟由that 或in which 引导的定语从句 正:I don’t like the way he spoke. I don’t like the way that he spoke. I don’t like the way in which he spoke.误:I don’t like the way how he spoke. 4. the way 的从句 That’s the way (=how) he spoke. I know where you are from by the way you pronounce my name. That was the way minority nationalities were treated in old China. Nobody else loves you the way(=as) I do. He did not do it the way his friend did. 二.固定搭配 1. In a/one way:In a way he was right. 2. In the way /get in one’s way I'm afraid your car is in the way, If you are not going to help,at least don't get in the way. You'll have to move-you're in my way. 3. in no way Theory can in no way be separated from practice. 4. On the way (to……) Let’s wait a few moments. He is on the way Spring is on the way. Radio forecasts said a sixth-grade wind was on the way. She has two children with another one on the way. 5. By the way By the way,do you know where Mary lives? 6. By way of Learn English by way of watching US TV series. 8. under way 1. Elbow one’s way He elbowed his way to the front of the queue. 2. shoulder one’s way 3. feel one‘s way 摸索着向前走;We couldn’t see anything in the cave, so we had to feel our way out 4. fight/force one’s way 突破。。。而前进The surrounded soldiers fought their way out. 5.. push/thrust one‘s way(在人群中)挤出一条路He pushed his way through the crowd. 6. wind one’s way 蜿蜒前进 7. lead the way 带路,领路;示范 8. lose one‘s way 迷失方向 9. clear the way 排除障碍,开路迷路 10. make one’s way 前进,行进The team slowly made their way through the jungle.
the way的用法大全
在the way+从句中, the way 是先行词, 其后是定语从句.它有三种表达形式:1) the way+that 2)the way+ in which 3)the way + 从句(省略了that或in which),在通常情况下, 用in which 引导的定语从句最为正式,用that的次之,而省略了关系代词that 或in which 的, 反而显得更自然,最为常用.如下面三句话所示,其意义相同. I like the way in which he talks. I like the way that he talks. I like the way he talks. 如果怕弄混淆,下面的可以不看了 另外,在当代美国英语中,the way用作为副词的对格,"the way+从句"实际上相当于一个状语从句来修饰全句. the way=as 1)I'm talking to you just the way I'd talk to a boy of my own. 我和你说话就象和自己孩子说话一样. 2)He did not do it the way his friend did. 他没有象他朋友那样去做此事. 3)Most fruits are naturally sweet and we can eat them just the way they are ----all we have to do is clean or peel them . 大部分水果天然甜润,可以直接食用,我们只需要把他们清洗一下或去皮. the way=according to the way/judging from the way 4)The way you answer the qquestions, you must be an excellent student. 从你回答就知道,你是一个优秀的学生. 5)The way most people look at you, you'd think a trashman was a monster. 从大多数人看你的目光中,你就知道垃圾工在他们眼里是怪物. the way=how/how much 6)I know where you are from by the way you pronounce my name. 从你叫我名字的音调中,我知道你哪里人. 7)No one can imaine the way he misses her. 人们很想想象他是多么想念她. the way=because 8) No wonder that girls looks down upon me, the way you encourage her. 难怪那姑娘看不起我, 原来是你怂恿的 the way =while/when(表示对比) 9)From that day on, they walked into the classroom carrying defeat on their shoulders the way other students carried textbooks under their arms.
“the-way+从句”结构的意义及用法知识讲解
“the way+从句”结构的意义及用法 首先让我们来看下面这个句子: Read the following passage and talk about it with your classmates. Try to tell what you think of Tom and of the way the children treated him. 在这个句子中,the way是先行词,后面是省略了关系副词that 或in which的定语从句。 下面我们将叙述“the way+从句”结构的用法。 1.the way之后,引导定语从句的关系词是that而不是how,因此,<<现代英语惯用法词典>>中所给出的下面两个句子是错误的:This is the way how it happened. This is the way how he always treats me. 2. 在正式语体中,that可被in which所代替;在非正式语体中,that则往往省略。由此我们得到the way后接定语从句时的三种模式:1) the way +that-从句2) the way +in which-从句3) the way +从句 例如:The way(in which ,that) these comrades look at problems is wrong.这些同志看问题的方法不对。
The way(that ,in which)you’re doing it is completely crazy.你这么个干法,简直发疯。 We admired him for the way in which he faces difficulties. Wallace and Darwin greed on the way in which different forms of life had begun.华莱士和达尔文对不同类型的生物是如何起源的持相同的观点。 This is the way (that) he did it. I liked the way (that) she organized the meeting. 3.the way(that)有时可以与how(作“如何”解)通用。例如: That’s the way (that) she spoke. = That’s how she spoke. I should like to know the way/how you learned to master the fundamental technique within so short a time. 4.the way的其它用法:以上我们讲的都是用作先行词的the way,下面我们将叙述它的一些用法。
定冠词the的12种用法
定冠词the的12种用法 定冠词the 的12 种用法,全知道?快来一起学习吧。下面就和大家分享,来欣赏一下吧。 定冠词the 的12 种用法,全知道? 定冠词the用在各种名词前面,目的是对这个名词做个记号,表示它的特指属性。所以在词汇表中,定冠词the 的词义是“这个,那个,这些,那些”,可见,the 即可以放在可数名词前,也可以修饰不可数名词,the 后面的名词可以是单数,也可以是复数。 定冠词的基本用法: (1) 表示对某人、某物进行特指,所谓的特指就是“不是别的,就是那个!”如: The girl with a red cap is Susan. 戴了个红帽子的女孩是苏珊。 (2) 一旦用到the,表示谈话的俩人都知道说的谁、说的啥。如:
The dog is sick. 狗狗病了。(双方都知道是哪一只狗) (3) 前面提到过的,后文又提到。如: There is a cat in the tree.Thecat is black. 树上有一只猫,猫是黑色的。 (4) 表示世界上唯一的事物。如: The Great Wall is a wonder.万里长城是个奇迹。(5) 方位名词前。如: thenorth of the Yangtze River 长江以北地区 (6) 在序数词和形容词最高级的前面。如: Who is the first?谁第一个? Sam is the tallest.山姆最高。 但是不能认为,最高级前必须加the,如: My best friend. 我最好的朋友。 (7) 在乐器前。如: play the flute 吹笛子
Way的用法
Way用法 A:I think you should phone Jenny and say sorry to her. B:_______. It was her fault. A. No way B. Not possible C. No chance D. Not at all 说明:正确答案是A. No way,意思是“别想!没门!决不!” 我认为你应该打电话给珍妮并向她道歉。 没门!这是她的错。 再看两个关于no way的例句: (1)Give up our tea break? NO way! 让我们放弃喝茶的休息时间?没门儿! (2)No way will I go on working for that boss. 我决不再给那个老板干了。 way一词含义丰富,由它构成的短语用法也很灵活。为了便于同学们掌握和用好它,现结合实例将其用法归纳如下: 一、way的含义 1. 路线
He asked me the way to London. 他问我去伦敦的路。 We had to pick our way along the muddy track. 我们不得不在泥泞的小道上择路而行。 2. (沿某)方向 Look this way, please. 请往这边看。 Kindly step this way, ladies and gentlemen. 女士们、先生们,请这边走。 Look both ways before crossing the road. 过马路前向两边看一看。 Make sure that the sign is right way up. 一定要把符号的上下弄对。 3. 道、路、街,常用以构成复合词 a highway(公路),a waterway(水路),a railway(铁路),wayside(路边)
way与time的特殊用法
way/time的特殊用法 1、当先行词是way意思为”方式.方法”的时候,引导定语从句的关系词有下列3种形式: Way在从句中做宾语 The way that / which he explained to us is quite simple. Way在从句中做状语 The way t hat /in which he explained the sentence to us is quite simple. 2、当先行词是time时,若time表示次数时,应用关系代词that引导定语从句,that可以省略; 若time表示”一段时间”讲时,应用关系副词when或介词at/during + which引导定语从句 1.Is this factory _______ we visited last year? 2.Is this the factory-------we visited last year? A. where B in which C the one D which 3. This is the last time _________ I shall give you a lesson. A. when B that C which D in which 4.I don’t like the way ________ you laugh at her. A . that B on which C which D as 5.He didn’t understand the wa y ________ I worked out the problem. A which B in which C where D what 6.I could hardly remember how many times----I’ve failed. A that B which C in which D when 7.This is the second time--------the president has visited the country. A which B where C that D in which 8.This was at a time------there were no televisions, no computers or radios. A what B when C which D that