microRNA与心肌缺血损伤研究新进展

中国组织工程研究第16卷第2期 2012–01–08出版
Chinese Journal of Tissue Engineering Research January 8, 2012 Vol.16, No.2 ISSN 1673-8225 CN 21-1581/R CODEN: ZLKHAH
357
microRNA与心肌缺血损伤研究新进展★
杨娇,程晓曙
Recent progress in microRNA and myocardial ischemic injury Yang Jiao, Cheng Xiao-shu
Abstract
BACKGROUND: Myocardial ischemia and hypoxia is the predominant cause of various cardiac diseases. Recent studies have
shown that numerous microRNAs show dynamic regulation during myocardial ischemia and hypoxia, suggesting their involvement
in the regulation of cardiovascular disease.
OBJECTIVE: To introduce the effects, mechanism and therapeutic strategy of microRNA on myocardial ischemic injure during the
past 5 years.
METHODS: Taking “microRNA, cardiac, ischemia, hypoxia” as English search terms, the articles during the past 5 years in
PubMed database and ISI Web of Knowledge database were retrieved by computer. The relevant literatures were included, the
literature of irrelevant purpose and repetitive content were excluded, and 42 of them were involved for further analysis.
RESULTS AND CONCLUSION: microRNAs are small endogenous RNA with post-transcriptional regulatory activity. They act as
negative regulators of gene expression by targeting 3’ UTR of mRNA, and are finally correlated with the cardiovascular disease.
Recent research has revealed that microRNAs have participated in the pathological progress related to myocardial ischemia,
post-ischemic cardiac remodeling and arrhythmia secondary to myocardial infarction. The microRNA expression in human
intervention can exacerbate or prevent the progress of myocardial ischemic and hypoxic injury. microRNA may become the target
molecular treatment of cardiovascular disease.
Yang J, Cheng XS. Recent progress in microRNA and myocardial ischemic injury. Zhongguo Zuzhi Gongcheng Yanjiu. 2012;16(2):
357-361. [https://www.360docs.net/doc/63445140.html, https://www.360docs.net/doc/63445140.html,]
摘要
背景:研究发现多种microRNA在心肌缺血缺氧时表达明显异常,它们对心血管疾病的发生和发展起着重要作用。
目的:介绍近5年来microRNA对心肌缺血损伤的影响、作用机制及可能的治疗方案。
方法:分别以“microRNA、心肌、缺血、缺氧”为检索词,应用计算机检索PubMed 数据库和ISI Web of Knowledge平
台检索近5年有关文章,排除与研究目的无关和内容重复者,保留42篇文献做进一步分析。
结果与结论:microRNA是一类具有转录后调节活性的内源性小分子RNA,通过与靶mRNA的3`端非编码区结合负性调控
基因的表达而参与心血管疾病的发生发展。目前研究表明microRNA参与了心肌缺血、缺血后心脏重塑、心肌梗死后继发性
心律失常等相关的病理过程,人工干预microRNA的表达可以加剧或预防心肌缺血缺氧损伤的进展。microRNA可能成为治
疗心血管疾病的靶向分子。
关键词:心肌缺血;microRNA(miRNA);缺血性心律失常;靶向治疗;综述文献
doi:10.3969/j.issn.1673-8225.2012.02.038
杨娇,程晓曙.microRNA与心肌缺血损伤研究新进展[J].中国组织工程研究,2012,16(2):357-361.
[https://www.360docs.net/doc/63445140.html, https://www.360docs.net/doc/63445140.html,]
0 引言
研究发现多种microRNA在心肌缺血缺氧时表达明显异常[1],表明不同的microRNA明显调控着心肌损伤、心律失常、心肌肥厚、心脏重塑等的发生和发展[2-3]。miRNA的表达具有组织特异性,一种miRNA可能在某种组织或细胞中高表达,而在其他组织及细胞中低表达或不表达。例如:miR-1具有骨骼肌及心脏特异性[4-5],而miR-145具有平滑肌细胞特异性[6]。miRNA在心血管的高度表达及心肌损伤时的异常表达给心肌缺血疾病的治疗带来了新的希望。1 资料和方法
1.1 资料来源
检索人相关内容:第一作者。
检索时间范围:2004-01/2011-06。
检索数据库:应用计算机检索Pubmed 数据
库有关文章ISI Web of Knowledge平台。
检索关键词:microRNA,心肌,缺血,缺氧。
检索文献量:共检索到89篇文献。
1.2 入选标准
纳入标准:文章所述内容必须属于
microRNA领域,主要围绕心肌缺血缺氧展开。
主要选择近5年、优先选择近3年内发表的具有
Department of
Cardiology, Second
Hospital of Nanchang
University, Nanchang
330006, Jiangxi
Province, China
Yang Jiao★,
Studying for master’s
degree, Department
of Cardiology,
Second Hospital of
Nanchang University,
Nanchang 330006,
Jiangxi Province,
China
jiaoyang_1963@163.
com
Correspondence to:
Cheng Xiao-shu,
Doctor, Professor,
Doctoral supervisor,
Department of
Cardiology, Second
Hospital of Nanchang
University, Nanchang
330006, Jiangxi
Province, China
xiaoshumenfan@
https://www.360docs.net/doc/63445140.html,
Received: 2011-07-13
Accepted: 2011-08-07
南昌大学第二附
属医院心内科,江
西省南昌市
330006
杨娇★,女,1987
年生,江西省萍乡
市人,汉族,南昌
大学在读硕士。
jiaoyang_1963@
https://www.360docs.net/doc/63445140.html,
通讯作者:程晓
曙,博士,教授,
博士生导师,南昌
大学第二附属医
院心内科,江西省
分子医学重点实
验室,江西省南昌
市 330006
xiaoshumenfan
@https://www.360docs.net/doc/63445140.html,
中图分类号:R318
文献标识码:B
文章编号: 1673-8225
(2012)02-00357-05
收稿日期:2011-07-13
修回日期:2011-08-07
(20110713005/W·S)
杨娇,等.microRNA 与心肌缺血损伤研究新进展
P .O. Box 1200, Shenyang 110004 https://www.360docs.net/doc/63445140.html,
358 www.CRTER .org
该领域代表性的文献,并选择了少数几篇引文中具有代表性的参考文献作为本文的参考文献。
排除标准:文章侧重点与microRNA 不相关或重复
性、陈旧文献。
1.3 资料提取 通过调查被引用次数,浏览标题、摘要和出版年限,再仔细阅读文献,最后确定42篇文献可用。
1.4 质量评估 尝试更多的关键词进行搜索,选择较多前沿的外文文献及专利,选择被引用次数较高的文献以保证文献的全面性和高质量。符合纳入标准的42篇文献中均为国外的相关研究报道及综述。
2 结果
2.1 microRNA 概况 microRNA(微小RNA ,miRNA)是近年发现的小的、内源性的单链非编码RNA ,由18~25个核苷酸组成,由一段具有发夹环结构的长度为70~80核苷酸的单链RNA 前体(pre-miRNA)剪切后生成[3]。人类基因组被检测出大约有1 000种microRNA ,在RNA 诱导的沉默复合体(RISC)的作用下靶向到达mRNA ,通过碱基互补配对的原则与靶mRNA 的3`端非编码区结合,促进靶mRNA 的降解或抑制其翻译负性调控基因的表达而发挥生物学功能[3]。miRNA 基因可以作为独立的转录本进行表达,也可以随多顺反子嵌入编码蛋白基因的内含子中进行表达,这两者几乎是一致的[2]。miRNA 在各种病理应激条件下表达明显改变[1],充分显示了它们在各种疾病中所发挥的作用。许多时候miRNA 都表现出集体功能的特点,而并非单一miRNA 所能替代[2]。在体内通过使用寡核苷酸抑制剂(antagomir)或miRNA 类似物(mimics)可人工调控miRNA 的表达,因此miRNA 应用于疾病的治疗成为了可能。
2.2 miRNA 与心肌缺血 心肌缺血是导致心肌细胞死亡的直接原因之一,缺血再灌损伤与缺血预适应保护现象的发现使研究者对这一领域有了新的认识。近年来随着miRNA 的发现,又将心肌缺血损伤与保护的研究推向深入。科研人员通过计算机分析及荧光素酶报告方法寻找miRNA 的靶基因[7],用miRNA 转基因及化学合成反义寡核苷酸(antagomir)或类似物(mimics)的方法[8]
,从靶mRNA 及各种信号通路探索研究,发现miRNA 在心肌缺血损伤中发挥着重要的作用。体外及体内实验均表明心肌细胞在缺氧条件下miRNA 水平可出现明显改变。miRNA 的上调或下调通过作用于靶基因激活相应的分子信号通路对心肌缺血损伤进行调控。 2.2.1 microRNA 的上调与心肌保护
miR-499/208:miR-499、miR-208a 、miR-208b 被称之为肌凝蛋白基因编码的miRNA 家族,分别由
Myh7b 、Myh6/αMHC 、 Myh7/βMHC 的内含子编 码[9-10]。在小鼠心脏中,βMHC 及miR-208 b 面对应激及甲状腺机能减退(甲减)环境下的反应性上调,需要miR-208a 的参与,同样miR-208a 也调节Myh7b 及miR-499的表达,但两者的调节阶段明显不同[10]。Myh7b/miR-499在心脏的发育分化过程中及正常生理情况下大量表达[11],而βMHC/miR-208b 在正常成年小鼠心脏不表达,只有在应激及甲减时才表达[10, 12]。miR-208a 在小鼠心脏发育过程中表达逐渐增加,到成年时达高峰,而miR-208b 的表达正好相反[12]。
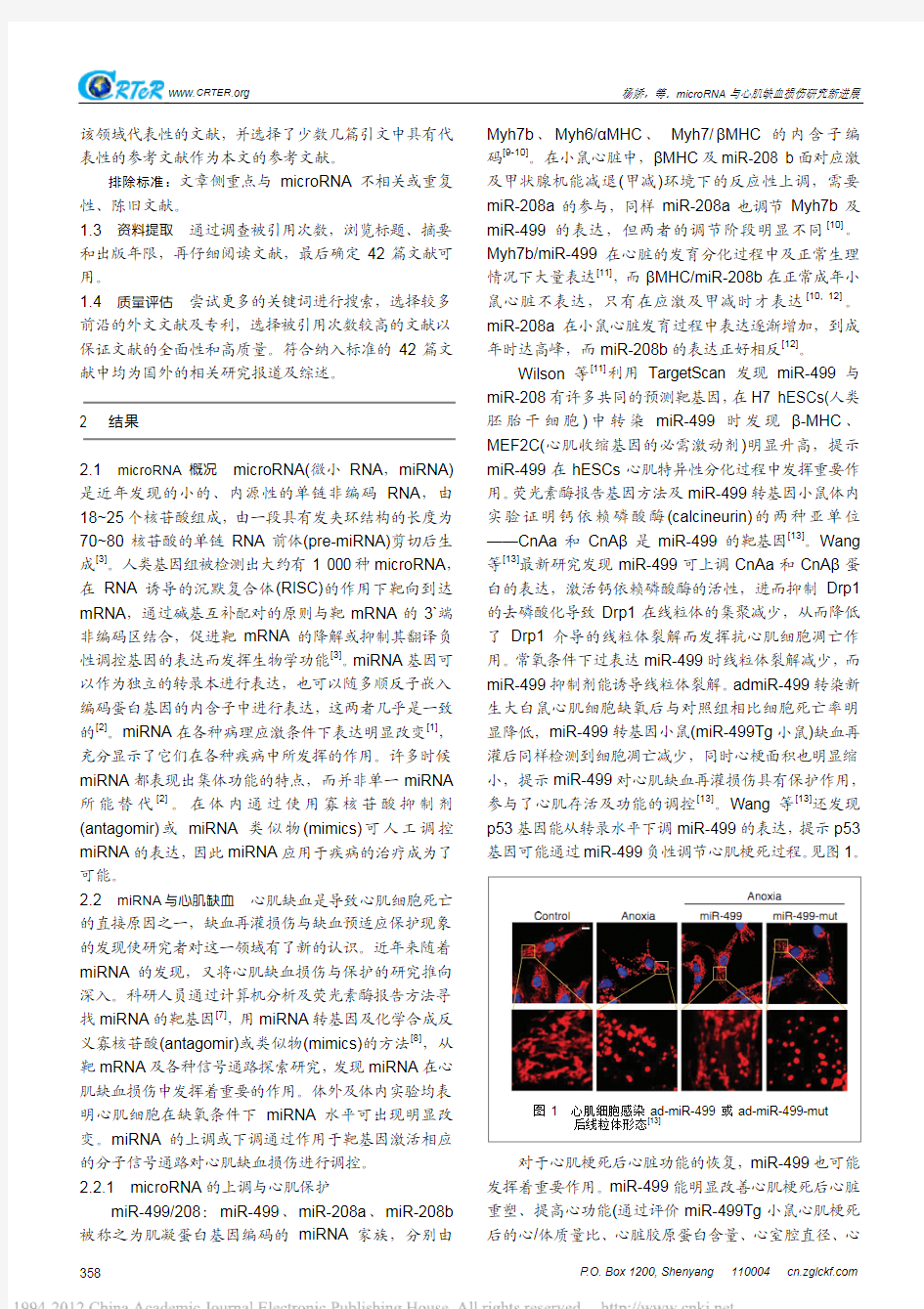
Wilson 等[11]利用TargetScan 发现miR-499与miR-208有许多共同的预测靶基因,在H7 hESCs(人类胚胎干细胞)中转染miR-499时发现β-MHC 、MEF2C(心肌收缩基因的必需激动剂)明显升高,提示miR-499在hESCs 心肌特异性分化过程中发挥重要作用。荧光素酶报告基因方法及miR-499转基因小鼠体内实验证明钙依赖磷酸酶(calcineurin)的两种亚单位——CnAa 和CnA β是miR-499的靶基因[13]。Wang 等[13]最新研究发现miR-499可上调CnAa 和CnA β蛋白的表达,激活钙依赖磷酸酶的活性,进而抑制Drp1的去磷酸化导致Drp1在线粒体的集聚减少,从而降低了Drp1介导的线粒体裂解而发挥抗心肌细胞凋亡作用。常氧条件下过表达miR-499时线粒体裂解减少,而miR-499抑制剂能诱导线粒体裂解。admiR-499转染新生大白鼠心肌细胞缺氧后与对照组相比细胞死亡率明显降低,miR-499转基因小鼠(miR-499Tg 小鼠)缺血再灌后同样检测到细胞凋亡减少,同时心梗面积也明显缩小,提示miR-499对心肌缺血再灌损伤具有保护作用,参与了心肌存活及功能的调控[13]。Wang 等[13]还发现p53基因能从转录水平下调miR-499的表达,提示p53基因可能通过miR-499负性调节心肌梗死过程。见图1。
对于心肌梗死后心脏功能的恢复,miR-499也可能发挥着重要作用。miR-499能明显改善心肌梗死后心脏重塑、提高心功能(通过评价miR-499Tg 小鼠心肌梗死后的心/体质量比、心脏胶原蛋白含量、心室腔直径、心
图1 心肌细胞感染ad-miR-499或ad-miR-499-mut
后线粒体形态[13]
杨娇,等.microRNA与心肌缺血损伤研究新进展ISSN 1673-8225 CN 21-1581/R CODEN: ZLKHAH
359 www.CRTER
.org
脏功能)[13]。miR-499可促进心脏干细胞的分化和发育,在人类心肌干细胞中表达miR-499时,可通过抑制其靶基因Sox6和Rod1的表达从而促进心肌梗死后心肌细胞的再生,明显改善心室功能[14]。
相关临床实验研究表明miR-499/208可能成为急性心肌梗死的重要生物学标记。Corsten等[15]对4种心脏病(急性心肌梗死、病毒性心肌炎、心脏舒张功能不全、急性心衰)患者血浆中提取的microRNAs选择性地进行RT-PCR检测,发现在急性心肌梗死患者血浆中miR-208b和miR-499 显著升高,分别上升1 600倍(P <0.005)和100倍(P < 0.000 5),并与肌钙蛋白T的升高密切相关。可见心脏受损时可启动心肌特异性microRNAs-208b 和miR-499释放入血循环。在去甲肾上腺素诱导的大白鼠心肌梗死模型中,Ji等[16]发现血浆中miR-208的浓度明显增加,且几乎与肌钙蛋白上升的水平及出现的时间窗一致。Bostjancic等[17]研究也检测到miR-208在急性心肌梗死患者血清中明显升高。
miR-21:miR-21在正常心脏中表达很低,但在各种心衰、心肌肥厚、缺血缺氧等条件下明显升高[18-19]。miR-21已被充分证明是导致心衰发展的一个重要调节因子,心衰时miR-21在成纤维细胞中选择性上调抑制了Spry1的表达,激活ERK–MAP激酶信号通路导致心肌纤维化而加剧心衰的发展[19]。
许多研究也证明miR-21参与了心肌细胞凋亡及缺血再灌损伤。在暴露于H2O2的心肌细胞中miR-21明显上调,Pre-miR-21可以明显减少H2O2诱导的心肌细胞凋亡,而miR-21抑制剂可直接上调靶基因PDCD4的表达抑制miR-21的心肌保护效应[20]。见图2。
Cheng等[21]在大白鼠缺血再灌模型中也证实了miR-21可作用于PDCD4发挥抗凋亡作用进而改善了心肌缺血损伤效应。过表达miR-21能明显减少心梗面积、降低左心室直径,而miR-21抑制剂逆转了此保护效应[22-23]。在急性心肌梗死后24 h,预先经Ad-miR-21转染的小鼠其心肌梗死面积比Ad-GFP组明显减少,梗死病灶边缘区miR-21明显升高,但在梗死区域内miR-21反而降低[22]。miR-21参与心肌缺血的保护效应已值得到肯定,除了能作用于PDCD4外,其他的预测靶基因及信号通路还需要进一步证实。
其他microRNA:microRNA-494可作用于促凋亡蛋白(ROCK1, PTEN,CAMKIIδ)及抗凋亡蛋白(FGFR2, LIF),激活蛋白激酶B(Akt/PKB)-线粒体信号通路发挥抗细胞凋亡及心肌保护作用[24]。
过表达miR-494的小鼠离体心脏缺血再灌后,标志细胞凋亡的指标LDH、组蛋白相关DNA片段、TUNEL 阳性细胞均减少,caspase-3的活性下降;相反利用antagomiR-494下调内源性miR-494时,细胞凋亡指标均上升;miR-494 TG小鼠心脏缺血再灌后心梗面积明显减小。miR-494 TG小鼠心脏中促凋亡蛋白(ROCK1, PTEN,CAMKIIδ)及抗凋亡蛋白(FGFR2, LIF)均明显下调,导致磷酸化的Akt升高2倍,从而激活了BAD(Bcl-2-associated death promoter,Ak的下游靶基因),进而线粒体信号分子抗凋亡蛋白Bcl-xL and Bcl-2上调,最终发挥了心肌细胞保护效应。
在体外及体内实验中miR-24可下调Bcl-2家族成员之一Bim (BH3-only domain–containing protein)抑制心肌细胞的凋亡[25]。
小鼠心肌梗死模型中心肌梗死后24 h梗死边缘区(BZ)miR-24明显下调,注入miR-24类似物的小鼠心梗边缘区凋亡细胞(TUNEL+、Caspase 3+细胞)明显减少、心梗面积也减小,12周时评价其心功能与对照组相比也有明显改善。在原代心肌细胞中也发现miR-24具有抗凋亡的作用。在小鼠体内实验中,当抑制内源性miR-24时Bim表达升高,激活了Caspase 3, Caspase 12和Caspase 9从而促进了心肌细胞的凋亡。Bim siRNA与miR-24抑制剂共同注入小鼠体内时,明显减弱了miR-24抑制剂的促凋亡作用。
miR-210可作用于Efna3和Ptp1b靶基因从抗细胞凋亡及促进血管生成方面明显改善心肌梗死后心脏功能[26]。见图3。
图2 miR-21对H2O2诱导心肌细胞凋亡的影响[20]
图3 马氏染色示心梗后8周MC-210组心梗面积明显减小
杨娇,等.microRNA 与心肌缺血损伤研究新进展
P .O. Box 1200, Shenyang 110004 https://www.360docs.net/doc/63445140.html,
360 www.CRTER .org
HL-1心肌细胞缺氧后用微阵列法检测到活细胞中的miR-210明显升高,HL-1心肌细胞转染Pre-miR-210后,可释放Leptin 、白细胞介素1α、肿瘤坏死因子α,降低了caspase 3/7的活性,从而促进血管生成、抑制细胞凋亡。在LAD 结扎的心肌梗死小鼠模型中注射MC-miR-210(携带miR-210的非病毒微环),左心室功能明显改善,心梗面积明显缩小、凋亡细胞减少、毛细血管密度明显升高。
2.2.2 microRNA 的上调与心肌损伤 miRNA-1在心肌细胞凋亡[27]、心律失常[28]、平滑肌细胞分化等方面都发挥着重要的作用[28-29]。miRNA-1具有抗心肌细胞肥大的作用[30],但却负性调节心肌缺血损伤,可通过调节靶基因HSP60、HSP70、 Bcl-2促进心肌细胞的凋
亡[27, 31-32]。
正常心肌细胞几乎不表达miR-1,但在各种病理条件下可大量表达,暴露于缺氧、H 2O 2、高糖环境下miR-1的水平明显升高[27, 31-32]。抑制miR-1可以减少髙糖诱导的心肌细胞死亡,而上调miR-1可以增加心肌细胞死 亡[27]。在髙糖处理的小鼠心肌细胞中,miR-1可通过抑制HSP60蛋白表达从而促进心肌细胞的凋亡,血清反应因子(SRF)及MEK1/2通路在miR-1的上游参与了此调节过程[27]。miR-1的上调受β肾上腺素受体–cAMP–蛋白激酶K 信号通路的调控,β肾上腺素受体可以下调miR-1的表达
[33]
。Cheng 等
[34]
建立Triton X-100诱导的
心肌细胞坏死模型,发现miR-1大量释放入培养基中并24 h 内保持稳定水平;在大白鼠心肌梗死模型中,同样检测到血清中的miR-1迅速增加,6 h 达高峰(超过200倍),同时还发现血清中miR-1的水平与心梗面积大小呈明显的正相关性。该组研究人员还进一步检测了心肌梗死患者循环血中的miR-1水平,也发现明显上升,并与CK-MB 呈明显的正相关性。
2.3 microRNA 与心肌梗死后心律失常 心律失常是心肌梗死后严重并发症之一,已有研究表明急性心肌梗死后microRNA 水平的改变可直接或间接介导离子通道表达失调而引起心脏电生理紊乱,目前研究较多的有miR-1 和miR-133。
急性心肌梗死中miR-1表达明显上调[17],而阻断miR-1可以降低心肌梗死后继发心律失常的发生[35]。过表达miR-1可作用于PP2A 选择性增加高磷酸化的RyR2通道,引起Ca 2+的释放激活心脏兴奋-收缩偶联导致心律失常的发生[36]。在大白鼠心肌梗死模型中过表达miR-1加剧了心律失常的发生,相反使用miR-1反义核苷酸抑制剂可减少心肌梗死后心律失常的发生
[37]
。
miR-1转录后抑制KCNJ2和GJA1的表达,降低心肌细胞传导性,使细胞膜去极化而促进了心律失常的发 生[37]。
急性心肌梗死中miR-133a 和miR-133b 的表达也
同样明显上调[17],心室肌细胞中过表达miR-133a 能降低TAC(主动脉缩窄术)相关的Ito,f(一过性外向K +电流),明显延长QT 间期及动作电位时程[38]。
前面已述miR-208a 在急性心肌梗死时明显升高,研究表明miR-208a 与心律失常的发生也有很大关系,miR-208a 转基因小鼠心电图可见PR 间期及房室传导阻滞,很可能是miR-208a 作用于GATA4介导Cn 40表达失调引起[12]。
2.4 microRNA 与心肌缺血治疗
2.4.1 药物对microRNA 调控心肌缺血的影响 虽然microRNA 对心血管疾病的调控机制研究近5年才成为焦点,但药物是否会影响microRNA 的表达进而控制疾病的发展这一问题已开始引起了部分学者的兴趣,最新已有很少的报道证实了这一现象的可能。
丹参酮IIA 可减轻心肌细胞缺血缺氧损伤、改善心功能,丹参酮IIA 通过p38 MAPK 信号通路(抑制活化的p38 MAPK 和心脏特异性转录因子SRF 及MEF2的表达)抑制缺氧诱导的心肌细胞中miR-1的上调从而发挥心肌保护效应[ 39]。有报道大鼠心肌梗死模型中丹参酮IIA 可下调miR-1从而部分恢复急性缺血时Kir2.1蛋白的表达及IK1电流强度[35],降低了缺血性心律失常的发生及心脏猝死率。Zhu 等[40]建立大白鼠心肌梗死模型,MI-PRO 组(口服心得安10 mg/(kg ·d)心梗面积明显减少,长期口服心得安使得心肌梗死时31种表达失调的miRNAs 中有18种逆转,提示心得安可能通过纠正缺血时miRNAs 表达的失调发挥心肌缺血保护作用。当然在这些实验中具体的作用机制并没很清楚,也许microRNA 只是在药物干扰下的一种反应性改变。 2.4.2 其他治疗方法 Hosoda [14]等将表达miR-499的心脏干细胞注入动物心肌梗死区,发现能明显促进心肌细胞的再生及分化,提高心功能。这意味着miR-499促进人类心脏干细胞的分化可能成为临床治疗心肌梗死后心衰的一种新治疗方案。静脉注射miRNA 反义核苷酸寡链(antagomir),可引起心脏等多种器官内相应miRNA 水平的显著下降[41],这表明miRNA 反义核苷酸寡链可在体内沉默特定的miRNA ,miRNA 也许可作为心血管疾病治疗的靶向分子。 3 讨论
miRNA 参与了心肌缺血缺氧过程中细胞的凋亡、再生、电传导等病理生理过程,在急性心肌缺血中的作用机制已逐渐得到认识,miRNA 可能成为心肌损伤的重要生物学标记[42],对预防及早期诊断急性心肌梗死带来帮助。miRNA 与心肌缺血治疗也有相关的研究表明了这一新方案的可能,由于miRNA 具有众多的靶基因,其作用也并非单一miRNA 所能实现的,需要其他一系
杨娇,等.microRNA 与心肌缺血损伤研究新进展
ISSN 1673-8225 CN 21-1581/R CODEN: ZLKHAH
361
www.CRTER .org
列复杂的信号分子参与,也许miRNA 也能相互干扰调节,这似乎妨碍了miRNA 作为治疗靶向分子的有效行,其安全性也引起了人们的关注。因此需要更深入地探索具有细胞特异性、基因特异性的miRNA 作用方式来调控基因表达以防治疾病的发生发展。
4 参考文献
[1] Bostjancic E,Zidar N,Glavac D.Microrna microarray expression
profiling in human myocardial infarction.Disease Markers.2009;27: 255-268.
[2] Small EM, Olson EN. Pervasive roles of micrornas in cardiovascular biology. Nature. 2011;469:336-342.
[3] Bartel DP.Micrornas: Genomics, biogenesis, mechanism, and function.Cell. 2004;116:281-297.
[4] Zhao Y,Samal E,Srivastava D.Serum response factor regulates a muscle-specific microrna that targets hand2 during cardiogenesis. Nature. 2005;436:214-220.
[5] Rao PK,Toyama Y,Chiang HR,et al.Loss of cardiac
microrna-mediated regulation leads to dilated cardiomyopathy and heart failure. Circulation Research. 2009;105:585-594. [6] Cordes KR,Sheehy NT,White MP,et al.Mir-145 and mir-143
regulate smooth muscle cell fate and plasticity. Nature. 2009;460: 705-U780.
[7]
Vo NK,Dalton RP,Liu N,et al.Affinity purification of microrna-133a with the cardiac transcription factor, hand2. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:19231-19236.
[8] Montgomery RL, van Rooij E. Therapeutic advances in microrna targeting. Journal of Cardiovascular Pharmacology. 2011;57:1-7. [9] McGuigan K,Phillips PC,Postlethwait JH.Evolution of sarcomeric myosin heavy chain genes: Evidence from fish. Molecular Biology and Evolution. 2004;21:1042-1056.
[10] van Rooij E,Quiat D,Johnson BA,et al.A family of micrornas encoded by myosin genes governs myosin expression and muscle performance. Developmental Cell. 2009;17:662-673.
[11]
Wilson KD,Hu SJ,Venkatasubrahmanyam S,et al.Dynamic microrna expression programs during cardiac differentiation of human embryonic stem cells role for mir-499.
Circulation-Cardiovascular Genetics. 2010;3:426-U497.
[12] Callis TE,Pandya K,Seok HY,et al.Microrna-208a is a regulator of cardiac hypertrophy and conduction in mice. Journal of Clinical Investigation. 2009;119:2772-2786
[13] Wang JX,Jiao JQ,Li QA,et al.Mir-499 regulates mitochondrial dynamics by targeting calcineurin and dynamin-related protein-1. Nature Medicine. 2011;17:71-U243.
[14] Hosoda T,Zheng HQ,Cabral-da-Silva M,et al.Human cardiac stem cell differentiation is regulated by a mircrine mechanism. Circulation.2011;123:1287-U1100
[15] Corsten MF,Dennert R,Jochems S,et al.Circulating microrna-208b and microrna-499 reflect myocardial damage in cardiovascular disease. Circulation-Cardiovascular Genetics.2010;3:499-506. [16] Ji X,Takahashi R,Hiura Y,et al.Plasma mir-208 as a biomarker of myocardial injury. Clinical Chemistry.2009;55:1944-1949.
[17] Bostjancic E,Zidar N, Stajer D,et al.Micrornas mir-1, mir-133a, mir-133b and mir-208 are dysregulated in human myocardial infarction.Cardiology. 2010;115:163-169.
[18] Patrick DM, Montgomery RL,Qi XX, et al. Stress-dependent
cardiac remodeling occurs in the absence of microrna-21 in mice. Journal of Clinical Investigation. 2010;120:3912-3916.
[19] Thum T, Gross C, Fiedler J, et al. Microrna-21 contributes to myocardial disease by stimulating map kinase signalling in fibroblasts. Nature. 2008;456:980-U983.
[20]
Cheng YH,Liu XJ,Zhang SO,et al.Microrna-21 protects against the h2o2-induced injury on cardiac myocytes via its target gene pdcd4. Journal of Molecular and Cellular Cardiology. 2009; 47: 5-14.
[21]
Cheng YH,Zhu P,Yang JA,et al.Ischaemic preconditioning- regulated mir-21 protects heart against ischaemia/reperfusion injury via anti-apoptosis through its target pdcd4. Cardiovascular Research. 2010;87:431-439.
[22]
Dong SM,Cheng YH,Yang J,et al.Microrna expression signature and the role of microrna-21 in the early phase of acute myocardial infarction. Journal of Biological Chemistry. 2009;284:29514- 29525.
[23] Yin C, Wang XY, Kukreja RC. Endogenous micrornas induced by heat-shock reduce myocardial infarction following
ischemia-reperfusion in mice. Febs Letters. 2008;582:4137-4142. [24]
Wang XH,Zhang XW,Ren XP,et al.Microrna-494 targeting both proapoptotic and antiapoptotic proteins protects against
ischemia/reperfusion-induced cardiac injury.Circulation.2010; 122:1308-U1145.
[25] Qian L,Van Laake LW,Huang Y,et al.Mir-24 inhibits apoptosis and
represses bim in mouse cardiomyocytes. Journal of Experimental Medicine. 2011;208:549-560.
[26] Hu SJ, Huang M, Li ZJ, et al.Microrna-210 as a novel therapy for
treatment of ischemic heart disease. Circulation. 2010;122: S124-S131.
[27] Shan ZX,Lin QX,Deng CY,et al.Mir-1/mir-206 regulate hsp60
expression contributing to glucose-mediated apoptosis in cardiomyocytes. Febs Letters. 2010;584:3592-3600.
[28] Chen J,Yin H,Jiang YL,et al.Induction of microrna-1 by myocardin
in smooth muscle cells inhibits cell proliferation. Arteriosclerosis Thrombosis and Vascular Biology. 2011;31:368-U287.
[29] Xie CQ,Huang HR,Sun XA,et al.Microrna-1 regulates smooth
muscle cell differentiation by repressing kruppel-like factor 4. Stem Cells and Development. 2011;20:205-210.
[30] Elia L,Contu R,Quintavalle M,et al.Reciprocal regulation of
microrna-1 and insulin-like growth factor-1 signal transduction cascade in cardiac and skeletal muscle in physiological and pathological conditions. Circulation. 2009;120:2377-2385.
[31] Xu CQ, Lu YJ, Pan ZW, et al.The muscle-specific micrornas mir-1
and mir-133 produce opposing effects on apoptosis by targeting hsp60, hsp70 and caspase-9 in cardiomyocytes. Journal of Cell Science. 2007;120:3045-3052.
[32] Tang YH, Zheng JY, Sun Y, et al. Microrna-1 regulates
cardiomyocyte apoptosis by targeting bd-2. International Heart Journal. 2009;50:377-387.
[33] Lu YJ,Zhang Y,Shan HL,et al.Microrna-1 downregulation by
propranolol in a rat model of myocardial infarction: A new mechanism for ischaemic cardioprotection. Cardiovascular Research. 2009;84:434-441.
[34] Cheng YH,Tan N,Yang JA, et al.A translational study of
circulating cell-free microrna-1 in acute myocardial infarction. Clinical Science. 2010;119:87-95.
[35] Shan HL,Li XL,Pan ZW,et al.Tanshinone iia protects against
sudden cardiac death induced by lethal arrhythmias via repression of microrna-1. British Journal of Pharmacology. 2009;158:1227-1235.
[36] Terentyev D,Belevych AE,Terentyeva R,et al.Mir-1
overexpression enhances ca2+ release and promotes cardiac arrhythmogenesis by targeting pp2a regulatory subunit b56 alpha and causing camkii-dependent hyperphosphorylation of ryr2. Circulation Research. 2009;104:514-521.
[37] Yang BF, Lin HX, Xiao JN, et al.The muscle-specific microrna
mir-1 regulates cardiac arrhythmogenic potential by targeting gja1 and kcnj2. Nature Medicine. 2007;13:486-491.
[38] Matkovich SJ, Wang W, Tu YZ, et al.Microrna-133a protects
against myocardial fibrosis and modulates electrical repolarization without affecting hypertrophy in pressure-overloaded adult hearts. Circulation Research. 2010;106:166-U340.
[39] Zhang Y,Zhang L,Chu WF, et al.Tanshinone IIA inhibits mir-1
expression through p38 mapk signal pathway in post-infarction rat cardiomyocytes. Cellular Physiology and Biochemistry. 2010;26: 991-998.
[40] Zhu WL, Yang L, Shan HL, et al. Microrna expression analysis:
Clinical advantage of propranolol reveals key micrornas in myocardial infarction. Plos One. 2011;6.
[41] Krutzfeldt J,Rajewsky N,Braich R, et al.Silencing of micrornas in
vivo with 'antagomirs'. Nature. 2005;438:685-689.
[42] Meder B,Keller A,Vogel B,et al.Microrna signatures in total
peripheral blood as novel biomarkers for acute myocardial infarction. Basic Research in Cardiology. 2011;106:13-23.
心肌缺血再灌注损伤
心肌缺血再灌注损伤的发生机制和防治研究进展 1 9 6 0年J e n n i n g s等,第一次提出心肌缺血再灌注损伤的概念,证实再灌注会引起心肌超微结构不可逆坏死,并逐渐引起医学界的高度重视。缺血心肌恢复再灌注后,病情反而恶化,引起超微结构、功能、代谢及电生理方面发生进一步的损伤,是由于在缺血损伤的基础上再次引起的损伤,因此称为缺血.再灌注损伤( i s e h e m i a — r e p e r f u s i o n i n j u r y ,I R I ) k 2 J 。临床上表现为闭塞的冠状动脉再通、梗死区血液灌流重建后一段时间内,有的病例发生血压骤降、心功能不全、心律失常甚至猝死等一系列病情反而恶化的现象。因此, I R I 的发生机制与防治越来越引起人们的关注,并一直试图寻找能对 I R I 产生确切保护作用的药物。现就 I R I的发生机制和防治的研究进展作一综述。 1 . 心肌缺血再灌注损伤的发生机制 目前,缺血再灌注损伤发生的机制尚未完全阐明,研究表明自由基、钙超载、心肌纤维能量代谢障碍、中性粒细胞、血管内皮细胞、细胞黏附分子与细胞凋亡等均可能参与缺血再灌注损伤。 1 . 1 氧自由基( F R) 生成正常细胞内有自由基清除剂超氧化物歧化酶( S O D),使氧自由基转变为过氧化氢,后者又通过触酶及谷胱甘肽过氧化物酶的作用还原为水和分子氧,故小量氧自由基不造成损伤。再灌注时产生的大量氧自由基不能被清除,其中包括非脂质氧自由基和脂质氧自由基,如超氧阴离子、羟自由基、过氧化氢等。缺血再灌注后,它可与各种细胞成分,如膜磷脂、蛋白质、核酸等发生反应,造成细胞结构损伤和功能代谢障碍。C a s t e d o 等在动物实验中发现,再灌注后细胞内膜脂质过氧化增强,形成多种生物活性物质,如血栓素、前列腺素等,促进再灌注损伤。 1 9 8 6年,M u r r y等,首次在犬缺血/再灌注模型实验中发现反复短暂缺血发作可使心肌在随后持续性缺血中得到保护,从而提出了缺血预适应( I P C) 心脏保护的概念,为缺血心肌的保护及其机制探讨开辟了崭新的领域。自由基可能参与了预适应保护的触发机制。Z h o n g等证明预适应过程中产生的低浓度自由基对延迟心肌缺血再灌注损伤有保护作用。冉擘力等从细胞水平证明早期产生的氧自由基能诱导延迟保护作用产生,其机制可能是通过早期氧化反应一方面改变S O D形态结构而提高酶的活性,诱导延迟相S O D合成增加,另一方面诱导热休克蛋白信使核糖核酸转录和持续合成,保护心肌细胞对抗细胞外氧自由基的损伤;氧化氮合酶( N O S ) 产生的一氧化氮能有效对抗氧自由基的损害,延迟期心肌 N O S活性增加。延迟保护作用增强,其机制可能是氧自由基诱导了后期 N O S信使核糖核酸转录和合成增加,因为在缺血等应激状态下,氧化氮能够调控心脏基因的表达。总之,热休克蛋白、抗氧化酶和 N O S等不是孤立地对抗氧自由基损伤,而是有机地结合起来发挥作用。 1 . 2 钙超载。生理状态下,胞浆内钙浓度约为 l 0-7 m o l / L ,而细胞外及胞浆内的钙储存系统( 如内质网和线粒体) 中钙浓度为1 0 -3m o l /L 。正常状态下,细胞通过一系列转运机制可以保持这种巨大的浓度梯度,以维持细胞内低钙状态。但是再灌注后,钙离子向线粒体转移,导致线粒体功能障碍;钙离子浓度升高,可激活多种酶( 如激活膜磷脂酶 A , )同时促使心
心肌缺血再灌注损伤介绍和实验设计
心肌缺血再灌注损伤介绍和实验设计 Ⅰ.心肌缺血再灌注损伤: 它是指缺血心肌组织恢复血流灌注时,导致再灌注区心肌细胞及局部血管网显著的病理生理变化,这些变化共同作用可促使进一步的组织损伤。那这里的关键词就是缺血心肌组织。那为什么会产生缺血的心肌组织呢?这就与临床上的疾病有关了。一些心脏疾病,比如急性心肌梗死、冠心病等他们会使心脏发生缺血的症状,其基本的生理过程就是心肌缺血。 Ⅱ.心肌缺血的危害: 心肌缺血:指单位时间内的冠脉血流量减少,供给组织的氧量也减少,缺血必定存在缺氧表明缺血缺氧。心肌缺血比单纯性心肌缺氧无血流障碍要严重,因为前者除了缺氧的影响之外,缺血组织也不能获得足够的营养物质又不能及时清除各种代谢产物带来的有害影响。 一、心肌缺血的原因主要分为两种情况:1是冠脉血流量的绝对不足。这种情况是由自身疾病产生的,主要包括冠状动脉阻塞,冠状动脉痉挛。2是冠脉血流量的相对不足:包括供氧降低或耗氧增加,比如高原高空或通风不良的矿井吸入氧减少;肺通气或换气功能障碍,可致血氧含量降低红细胞数量和血红蛋白含量减少等。 二、缺血对心肌的危害主要包括以下几个方面:1是心肌收缩能力降低。2是导致心肌舒张功能降低。3是心肌组织的血流动力学发生改变,比如说血流的阻力增加等。4是心肌电生理的变化,比如说静息点位降低,传导速度减慢;室颤阈降低等。5是导致心肌形态学的改变。当然还有其他的危害,在这里就不一一列举了。 由于心肌缺血存在这么多的危害,临床上针对这一疾病采取了再灌注治疗方法,但随之而来的又是另外一个临床问题:缺血再灌注损伤。 下面具体介绍一下心肌缺血再灌注损伤。心肌缺血再灌注损伤英文缩写为MIRI,最早由詹宁斯等于1960年提出,发现其临床表现为再灌注心律失常、心肌顿抑、心肌能量代谢障碍等现象。随后又有学者在临床手术中也证实了这一观点,发现在冠脉搭桥术完成后,心肌坏死进一步加重的现象。接着布朗沃尔德教
氧自由基与心肌缺血再灌注损伤
缺血性心脏病是导致人类死亡的主要原因,在治疗上,早期成功恢复心肌再灌注是改善临床转归的最有效方法。但缺血心肌恢复血流的过程可造成损伤,这一现象称为心肌缺血/再灌注损伤(myocardial ischemia/reperfusion injury,MI/RI)[1 2]。而氧自由基(oxygen free radical,OFR)也是心血管疾病时诱导心肌细胞死亡的重要因素之一[3]。在正常生理条件下,细胞内存在抗氧化物质可以及时清除OFR,使自由基的生成与降解处于动态平衡,对机体无害,而在心肌缺血再灌注损伤情况下,由于OFR生成过多或机体抗氧化能力不足,引发氧化应激反应,介导心肌损伤[4 5]。本研究重点阐述OFR与心肌缺血再灌注损伤之间的关系。 1 OFR合成、清除及生物学作用 自由基(free radical)是指具有一个不配对电子的原子和原子团的总称。由氧诱发的自由基称为OFR,主要包括超氧阴离子(O-2)、过氧化氢(H2O2)和羟自由基(OH)[6]。H2O2本身并非自由基而是一种活性氧(reactive oxygen species,ROS),但它与OFR的产生有密切关系,易接收一个电子生成羟自由基(OH)。正常情况下OH不能形成,因为OH的形成要求O-2及H2O2同时存在。当O-2及H2O2在组织中过剩, O-2及H2O2在金属离子及金属离子复合物的催化下发生Haber Weiss反应,生成氧化性更强的OH。OH是十分不稳定的氧化物,几乎与细胞内所有的有机物反应,破坏核酸、蛋白质、氨基酸和脂类化合物,从而损害细胞功能[7]。在生理情况下,氧通常是通过细胞色素氧化酶系统接收4个电子还原生成H2O,同时释放能量,但也有1%~2%的氧接收1个电子生成O-2,或再接收1个电子生成H2O2。O-2寿命极短,可通过连锁反应产生OH,H2O2能直接或间接促进细胞膜脂质过氧化。 自由基反应的扩展较广,但生物体内存在一套完整的抗氧化酶和抗氧化剂系统,可以及时清除它们,所以对机体无害。抗氧化酶包括超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH PX)和过氧化氢酶(CA T)。它们存在于胞浆和线粒体中,其重要意义在于降低H2O2浓度,保护细胞不受强毒性OFR OH的损伤。抗氧化剂包括存在于细胞质的维生素E 和维生素A;细胞外液中的半胱氨酸、抗坏血酸、谷胱甘肽;存在胞浆中的还原型谷胱甘肽(GSH)和还原型病理辅酶Ⅱ(NADPH)等。在OFR清除系统功能降低或丧失,生成系统活性增强,一旦恢复组织血液供应和氧供,OFR便大量产生与急剧堆积,从而造成心肌细胞急性或慢性损伤[8]。特异靶向抑制NADPH氧化酶可以减弱心血管氧化应激[9]。 2 OFR在心肌缺血再灌注损伤中的作用及地位 目前关于心肌缺血再灌注损伤的发病机制有许多假设和报道,主要与心肌再灌注时与OFR损伤、细胞内Ca2+超载、心肌细胞能量代谢障碍[10]、微血管损伤和粒细胞浸润以及心肌细胞的凋亡等作用有关。MI/RI时OFR合成增多主要与线粒体单电子还原、黄嘌呤氧化酶形成增多、儿茶酚胺自氧化增强、细胞内钙超载以及中性粒细胞呼吸暴发等有关[11]。由于OFR产生过多以及抗氧化酶类活性下降,引发链式脂质过氧化反应,损伤细胞膜、细胞器乃至细胞核酸,导致细胞坏死凋亡。应用外源性OFR清除剂及抗氧化剂则能降低组织中OFR浓度,促进心功能恢复,表明OFR在心肌缺血再灌注损伤中起着重要作用。 3 OFR与脂质生物膜
心肌缺血再灌注损伤机制的研究进展
心肌缺血再灌注损伤机制的研究进展 摘要急性心肌梗死是临床常见急症重症,及时、有效的恢复心肌的血液灌注,挽救“濒死”的心肌是抢救成功的关键,因此探索缺血再灌注损伤的机制,减轻或防止再灌注损伤的发生,是临床的重要课题。本文综述了心肌缺血再灌注损伤发生机制研究领域的最新进展。 关键词心肌缺血再灌注;氧自由基;钙超载;中性白细胞;血管内皮细胞;一氧化氮;细胞黏附因子;细胞凋亡 急性心肌梗死(AMI)是临床常见急症重症,及时、有效的恢复心肌的血液灌注,挽救“濒死”的心肌是抢救成功的关键。探索心肌再灌注损伤(MRI)的机制,减轻或防止再灌注损伤的发生,是临床的重要课题。至今为止MRI的机制还没有完全清楚,目前主要认为与氧自由基、钙超载、活化的中性白细胞、心肌纤维能量代谢障碍、血管内皮细胞、一氧化氮、细胞黏附因子和细胞凋亡等都可能参与MRI的发病过程。[1、2] 1氧自由基与心肌缺血再灌注损伤 生理情况下,细胞内存在的抗氧化物质可以及时清除自由基,对机体并无有害影响。当组织细胞缺血、缺氧时,由于活性氧生成过多或机体抗氧化能力不足,可引起氧化应激反应,造成膜流动性与钙离子通透性增加,破坏膜结构完整性,钙跨膜内流与超负荷导致细胞损伤甚至死亡。氧化应激是缺血组织再灌注的特征之一。而且应用自由基清除剂辅酶Q10[3]可以减轻缺血再灌区细胞的损伤。 2钙超载与心肌缺血再灌注损伤 近年研究表明,细胞内Ca2+超载在心肌缺血再灌注损伤发病机制中起中心作用。钙超载可以造成线粒体功能障碍,激活磷脂酶类,使细胞膜及细胞器膜结构受到损伤。还可激活蛋白酶,促进细胞膜和结构蛋白的分解,同时促进氧自由基的生成。激活某些ATP酶和核酶,加速ATP消耗,引起染色体损伤。Ca2+超载还可引起再灌注心律失常。心肌缺血再灌注损伤的始动环节是能量代谢障碍,而直接损伤原因则是自由基,其结果导致细胞内钙超载,并形成恶性循环。钙超载
心肌缺血再灌注
大鼠心肌缺血/再灌注损伤 【实验目的】 1.复制大鼠在体与离体心肌缺血/再灌注损伤模型; 2.观察缺血/再灌注过程中心功能的变化 【实验动物】成年Wistar 大鼠(体重200-300g) 【仪器药品】 电子天平,肾形盘,动物呼吸机,BL-420F记录装置,眼科开睑器,微血管钳,组织镊,眼科镊,组织剪,眼科剪,眼科止血钳,止血钳,动脉夹,眼科缝合针,1号及00缝合线。Langedroff灌流装置。 20%乌拉坦,1ml注射器,5ml 注射器,纱布块 实验1 在体模型 【实验步骤】 1.实验采用体重200-300g健康雄性Wistar大鼠,20%乌拉坦腹腔注射麻醉(0.5ml/100g); 2.颈胸部备皮及手术,分离气管及右侧颈总动脉 3.气管插管连接呼吸机(呼吸肌参数:潮气量9ml,呼吸比=3:2,呼吸频率55~60) 4.经右侧颈总动脉逆行插管至左心室, 再经BL-420F软件输入计算机,(一通道描记心 电,(右上黄、右下黑、左下红)二通道描记心室内压,三通道描记微分)持续监测心脏左心室内压力及心电的变化情况 5. 沿胸骨左侧剪开2,3肋骨,开睑器开胸暴露心脏;寻找冠状动脉左前降支,穿线备 用; 6.采用结扎5min后再放开5min两次,造成缺血预处置;采用结扎30mim再放开30min 复制缺血/再灌注模型; 思考题: 1.如何判定缺血模型复制成功 2.如何判定有再灌注损伤发生
实验2 离体模型 【实验步骤】 (1) 大鼠称重,腹腔注射20%乌拉坦(0.5ml/100g)麻醉,仰卧固定于鼠板,上腹部及前胸部剪毛。 (2) 舌下/阴茎背静脉注入1%肝素(0.05ml/100g)后,切开胸腹部皮肤,用剪刀横行剪开腹腔,向上剪断隔膜,沿两侧肋骨向上平行剪开,翻起前胸壁,把心脏及胸膈周围的结缔组织拨到一侧,充分暴露心脏。 (3) 用镊子提起心脏根部,暴露出主动脉和肺动脉,在距主动脉起始部0.5cm处用手术剪切断血管,迅速取出心脏至于4℃生理盐水平皿中使之停搏。 (4) 经主动脉将心脏悬挂在灌流装置上,用丝线结扎固定,打开灌流液行逆向灌流,待心脏恢复自主跳动,小心减去心脏周围附着组织。 (5)用眼科剪剪去左心耳,通过左心耳经房室瓣插入左心室一乳胶球囊,球囊连接一个内充生理盐水的导管,导管经三通管和换能器与BL-420F连接。 (6) 在BL-420F仪的监测下,通过向球囊内注入一定量的生理盐水是左心室的舒张末压调整在0~10mmHg之间。 (7)连接心电导线,心尖、右心耳和地线,一通道设置记录, (8) 预灌流10~20分钟,观察心率,二通道记录心室内压、三通道取微分记录±dp/dtmax 等心动指标,同时描记ECG,待上述各指标平衡后开始以下实验。 心肌缺血-再灌注损伤 (1) 心脏用正常灌流液预灌流15分钟后完全停灌40分钟,然后恢复灌流20分钟,观察心脏在正常,停灌初期和再灌期的心功能变化。 (2) 分别收集正常灌流时,再灌流后3分钟时的心脏冠脉流出液1ml,测定其中乳酸脱氢酶的活性。 思考题: 1.如何判定缺血模型复制成功 2.如何判定有再灌注损伤发生
心肌缺血再灌注损伤的机制研究进展
? 文献综述 ? 63 心肌缺血再灌注损伤(myocardial ischemic reperfusion in j ury ,MIRI )指心肌缺血恢复血流供应后,造成代谢功能障碍及结构损伤加重的现象[1]。MIRI 是临床上常见的疾病,其病理过程与冠状动脉血管形成术,冠状动脉重建术,心脏移植等术后并发症密切相关[2]。MIRI 涉及的机制复杂,尚有待更深入的研究阐述。近年来,由于电生理学、基因组学和蛋白组学等技术的应用,对MIRI 机制的研究也获得了一定的进步,其主要机制概述如下:1 氧自由基与MIRI 自由基(free radical ),又称游离基,指在外层电子轨道上具有不配对的单个电子、原子、原子团或分子的总称[3] 。由机体内氧诱发化学性质活泼的自由基称为氧自由基,包括羟自由基和超氧阴离子。生理状态下自由基存在较少,在细胞缺血时,其氧自由基清除能力下降[4]。当组织恢复血液供应时,触发氧自由基“爆增”并累积,攻击自身和周围细胞,造成损伤[5]。自由基损伤细胞膜,致其结构破坏造成心肌酶溢漏;自由基氧化破坏机体蛋白,改变蛋白酶表面结构使功能受损;自由基诱导遗传物质DNA 、RNA 断键或破损,影响核酸正常功能[6]。自由基可导致心律失常,心肌损伤,细胞凋亡等事件[7]。2 炎症反应与MIRI MIRI 发生时心脏组织内皮结构受损触发功能障碍,而中性粒细胞趋集、黏附血管内皮是炎症“级联”反应的诱发阶段[8] 。激活的中性粒细胞合成释放肿瘤坏死因子、IL-1、IL-6 等炎症介质,介导其他炎症细胞共同攻击心肌组织[9] 。此外,白细胞浸润在MIRI 中涉及的主要机制为,MIRI 使细胞膜受损和膜磷脂降解,具有很强趋化作用的白三烯等代谢产物增多,使更多白细胞循环浸润,对心肌细胞造成多次损伤。MIRI 时,心肌缺血细胞生成大量的促炎介质如补体C 5a 、LPS 、IL-8等,激活并诱导心肌细胞多种黏附如ICAM-1,ICAM-2等分子表达[10]。膜表面的黏附分子作为受体和配体介导白细胞与内皮细胞、心肌细胞的黏附,并为炎性浸润提供物质基础。3 钙超载与MIRI 由于细胞内钙浓度显著升高并造成心脏功能代谢障碍的现象称为钙超载(Ca 2+ 超载)[11] 。生理条件下,钙浓度稳态维持着正常心功能。当心肌缺血时,钠泵功能障碍,Na + 与Ca 2+ 的交换紊乱,使细胞内Ca 2+大量积累,触发线粒体功能障碍、钙泵障碍等[12]。Ca 2+超载与细胞损伤有相关性。其可引起:①减少线粒体ATP 生成。②激活钙依赖性降解酶,损伤细胞结构。③诱导自由基生成,损害心肌细胞。④促使 Ca 2+与CaM 结合,影响细胞内信号转导。⑤引起心律失常。 4 能量代谢障与MIRI MIRI 发生时,心肌细胞依赖无氧代谢途径供能,但其生成ATP 的能力有限。而ATP 的明显不足会触发一系列代谢的异常和紊乱:①依赖性ATP 的细胞膜泵活性下降,膜电位改变。②Ca 2+内流增加,激活膜磷酶导致缺血性肌挛缩,并产生氧自由基进一步损害细胞。③酸中毒,破坏细胞的生存环境。④严重阻碍ATP 的生成[13]。研究表明,能量代谢障碍可造成有关基因及蛋白表达的异常,同时细胞内的ATP 含量是触发细胞凋亡促进因素之一。5 细胞凋亡与MIRI 细胞凋亡,又称程序性细胞死亡,指由促凋亡因素触发细胞内死亡程序而发生的细胞死亡过程[14]。细胞凋亡调控着机体中细胞稳态,并摒除体内有害的细胞、无功能的细胞、突变的细胞以及受损的细胞。而过度活跃的细胞凋亡进程会加重MIRI 病情。MIRI 中的细胞凋亡的机制涉及的凋亡途径多种途径,以多方式、多水平的交叉联系,构成复杂的信号通路网络。线粒体途径、细胞因子信号转导途径、JAK-STAT 途径、LOX-1通路、MAPKs 通路等均可介导心肌MIRI 发生发展,造成的心肌细胞凋亡。提示抗凋亡作用或特异性对抗有关信号通路是治疗MIRI 的有效措施之一。6 小 结 综上所述,心肌缺血再灌注损伤(MIRI )的发生机制涉及多因素的复杂过程,需要广大科研攻关者更全面、更深入的科学研究,积极寻求更有效的防治措施,为MIRI 造福。近年来,随着科学技术的不断发展,在基因调控、细胞凋亡、信号转导等角度的深层次研究也在逐步开展,期待对MIRI 机制研究取得重要的突破。 参考文献 [1] 赵亚玲,敖虎山.心肌缺血再灌注损伤的研究进展[J].中国循环杂 志,2011,26(5):396-398. [2] C astedo E,Segovia J,Escudero C,et a1.Ischemia-reperfusion in j ury during experimental heart transplantation. Evaluation of trimetazidine's cytoprotective effect[J].Rev Esp Cardiol. 2005,58(8):941-950. [3] C hen AF,Chen DD,Daiber A,et a1.Free radical biology of the cardiovascular system[J].Clin Sci (Lond),2012,123(2):73-91.[4] V al ko M,Leibf r itz D,Moncol J,et a1.Free radicals and antioxidants in normal physiological functions and human disease [J].Int J Biochem Cell Biol, 2007,39(1):44-84. [5] D r?ge W.Free radicals in the physiological control of cell function[J].Physiol Rev, 2002,82(1):47-95. [6] 林灼锋,李校坤,孟娟.活性氧自由基对心肌细胞损伤效应研究[J]. 心肌缺血再灌注损伤的机制研究进展 邓海英* 赖为国 (钦州市第二人民医院药剂科,广西 钦州 535099) 【摘要】冠心病严重危害人类的生命健康,主要临床表现为心绞痛或心肌梗死。心肌缺血后再获取血液供应,常会出现心律失常、梗死面积扩大、心功能低下等心肌细胞损伤现象,即心肌缺血再灌注损伤(MIRI )。国内外研究表明MIRI 发生机制较为复杂,目前认为与再灌注后机体氧自由基攻击,炎症反应浸润,Ca 2+超载,能量代谢障碍、细胞凋亡进程等有关。现对MIRI 的机制及治疗的研究进展综述如下。本文通过归纳并总结有关MIRI 研究进展的国内外文献,对MIRI 的机制做出综述。【关键词】心肌缺血再灌注;损伤;机制 中图分类号:R542.2 文献标识码:A 文章编号:1671-8194(2013)01-0063-02 *通讯作者:E-mail: denghaiying2012@https://www.360docs.net/doc/63445140.html,
心肌缺血及其损伤标志物简介
心肌缺血及其损伤标志物简介 发表时间:2019-08-16T10:24:42.777Z 来源:《中国保健营养》2019年第3期作者:周凤莲[导读] 心脏是人体最重要的器官之一,它和动脉、静脉、毛细血管组成机体的循环系统,是血液循环的动力器官,其节律的收缩和舒张,使血液在血管里循环流动,保证各器官和组织得到充分的血液供应,以获得氧和各种营养物质,并带走代谢的终产物如二氧化碳、尿素和尿酸等, (资阳市人民医院检验科四川资阳 641300) 【中图分类号】R473.52 【文献标识码】A 【文章编号】1004-7484(2019)03-0053-01心脏是人体最重要的器官之一,它和动脉、静脉、毛细血管组成机体的循环系统,是血液循环的动力器官,其节律的收缩和舒张,使血液在血管里循环流动,保证各器官和组织得到充分的血液供应,以获得氧和各种营养物质,并带走代谢的终产物如二氧化碳、尿素和尿酸等,使细胞维持正常的代谢和功能。心肌损伤是指伴心肌细胞坏死的疾病,多由心脏缺血所致,包括急性心肌梗死(acute myocardial infarction,AMI)、心绞痛和心肌炎等,其中急性心肌梗死是冠心病发展到严重阶段的一种类型,在我国发病率逐年增加,是临床上常见的急危重症之一,及早诊断和治疗是提高疗效、改善预后的关键。其诊断除根据临床症状和体征外,主要依靠实验室检查。心肌损伤标志物是指当心肌细胞损伤时,可大量释放到血液中的特异性物质,检测其在血中浓度的变化可反映心肌损伤及其程度,可以为急性心肌梗死及其他伴有心肌损伤疾病的早期诊断、疗效观察提供极有价值的信息,同时也是目前临床评估病情和判断预后的灵敏指标。目前临床上反映心肌缺血损伤的主要生化标志物包括心肌酶和心肌蛋白。 1 心肌酶谱 1.1 血清天门冬氨酸转氨酶天门冬氨酸转氨酶(AST)又叫谷草转氨酶(GOT),广泛分布于人体各组织,肝脏、骨骼肌、肾脏、心肌内均含量丰富,红细胞内AST约为血清的10倍,溶血会使其测定结果偏高。在急性心肌梗死发生后6-12小时血清AST水平升高,24-48小时达到峰值,大概持续5天到1周,随后会降低。由于其不具备组织特异性,敏感性也不高,其单纯升高不能诊断心肌损伤,当今学术界已不主张将其用于急性心肌梗死的诊断。 1.2 血清乳酸脱氢酶及其同工酶乳酸脱氢酶(lactate de hydrogenase,LD)是葡萄糖无氧酵解中调节丙酮酸转化为乳酸的极重要的酶,广泛存在于肝脏、心脏、骨骼肌、肺、脾脏、脑、红细胞、血小板等组织细胞的胞质和线粒体中。发生心肌损伤时,心肌细胞膜破裂,线粒体、胞质内物质外漏到细胞间液及外周血中。LD和LD1在急性心肌梗死发作后8-12小时出现在血中,48-72小时达峰值,LD的半衰期为57-170小时,7-12天恢复正常,如果连续测定LD,对于就诊较迟、CK已恢复正常的AMI患者有一定的参考价值,但是由于它存在于机体多种组织,故其诊断心肌损伤的特异性不高。另外,由于红细胞内含有丰富的LD,而溶栓疗法常致溶血使血清LD活力升高,所以LD无法用于溶栓疗效的评估。 1.3 血清肌酸激酶及其同工酶肌酸激酶(creatine kinase,CK)是心肌中重要的能量调节酶,它是一种二聚体,由M和B两个亚基组成,形成CK-MM、CK-MB和CKBB三种同工酶。CK-BB存在于脑组织,CK-MM和CK-MB存在于各种肌肉组织;不同肌肉同工酶比例不同,骨骼肌中98%-99%是CK-MM,1%-2%是CK-MB;心肌内80%左右是CK-MB。在20世纪60年代CK就用于诊断急性心肌梗死,1972年CK-MB首次用于临床,两者对于诊断AMI贡献卓著,是世界上应用最广泛的心肌损伤指标。它们既可以用于早期诊断AMI,也可以用于估计梗死范围的大小或再梗死。AMI发生后4-6小时CK和CK-MB即可超过正常上限,24小时达峰值,48-72小时恢复正常。另外,CK常用于再灌注的效果观察,溶栓后几小时内CK-MB还会继续升高,称“冲洗现象”,此后CK即下降。由于CK同工酶的特异性和敏感性高于总CK,临床倾向于用CK-MB替代CK作为心肌损伤的常规检查项目。 2 血清心肌蛋白 2.1 心肌肌钙蛋白T 血清中心肌肌钙蛋白T(cTnT)属于心肌肌原纤维蛋白,它以游离的形式存在于心肌细胞胞质中,当心肌细胞损伤时释放入血液中,是诊断AMI的确定性标志物。AMI发病后3-6小时血清cTnT即升高,10-24小时达峰值至参考值的30-40倍,恢复正常需要10-15天,对Q波性、亚急性心肌梗死或CK-MB无法诊断的患者更有价值;cTnT还可用于溶栓疗法成功与否的评估,观察冠状动脉是否复通;另外,监测不稳定心绞痛患者的cTnT水平可以发现一些轻度和小范围的心肌损伤;它同时也是诊断心肌炎时比CK-MB敏感得多的指标。 2.2 心肌肌钙蛋白I 心肌肌钙蛋白I(cTnI)是一个十分敏感和特异的急性心肌梗死标志物。心肌内cTnI含量很丰富,心肌损伤后4-6小时释放入血,达到诊断决定值,心肌缺血症状发作后14-36小时达到高峰,5-10天后恢复正常,7天后cTnI诊断AMI的敏感性超过LD1/LD2,有文献报道测定血清cTnI诊断AMI的敏感性为97%,特异性为98%,预测值为99.8%;和cTnT 一样,cTnI也可用于溶栓后再灌注的判断,在成功溶栓疗法使冠状动脉复通后30、60分钟,cTnI还会继续升高,其敏感性高于CK-MB和肌红蛋白;另外,cTnI还可敏感地测出小灶可逆性心肌损伤的存在。 2.3 肌红蛋白肌红蛋白(Mb)是一种氧结合蛋白,广泛存在于骨骼肌、心肌和平滑肌,Mb分子量小于CK-MB和LD,且位于细胞质内,故心肌损伤后出现较早,到目前为止,它是AMI发生后出现最早的可检测的标志物之一。当AMI患者发作后,细胞质中的Mb释放入血,2小时即升高,6-9小时达到高峰,24-36小时恢复至正常水平;Mb的阴性预测值为100%,在胸痛发作2-12小时内,如果Mb阴性可排除急性心肌梗死;由于Mb消除快,因而它也是判断再梗死的良好指标,再梗死发生后,血清中可出现新的Mb浓度峰;由于Mb特异性不高(60%-95%),单凭其水平决定是否使用溶栓疗法具有一定风险,而碳酸酐酶III(carbonic anhydrase III,CAIII)特异性高,仅见于骨骼肌损伤,联合检测CAIII可提高AMI诊断的特异性,当骨骼肌损伤时Mb和CAII两者均升高,当心肌梗死时Mb升高而CAIII却始终正常。 2.4 心脏型脂肪酸结合蛋白脂肪酸结合蛋白(fatty acid binding protein,FABP)可与长链脂肪酸发生可逆性非共价结合,是一种胞内蛋白质,在脂肪酸代谢活跃的组织中含量丰富,如心脏、肝脏、肠等。目前已发现9种FABP,具有不同的组织学分布特征。心脏型脂肪酸结合蛋白(heart-type fatty acid binding protein,H-FABP)在心肌损伤后释放入血液,可作为AMI 损伤的早期标志物。H-FABP在心肌损伤后释放入血液的特点与Mb类似,在心肌缺血或损伤0.5-2小时内即可显著升高,6小时达到峰值,24-36小时内恢复正常水平;在早期(胸痛发生6小时内)诊断AMI的敏感度等于甚至优于Mb,当心肌损伤后,血浆H-FABP升高速率高于肌红蛋白及肌钙蛋白,它和Mb一样可用于AMI的早期排除;此外,H-FABP也可用于评估心肌梗死大小、冠状动脉再灌注及冠状动脉旁路手术,同时也可作为心肌缺血的标志物。
心肌缺血再灌注损伤防治的研究现状及展望
广东医学2019年1月第40卷第2期Guangdong Medical Journal Jan.2019,Vol.40,No.2?305?心肌缺血再灌注损伤防治的研究现状及展望 罗峰,苏强A 广西医科大学第一附属医院心血管内科(广西南宁530022) 【摘要】尽管急诊经皮冠状动脉介入术(primary percutaneous coronary intervention,PPCI)治疗可以及早 恢复心肌的灌注,但急性ST段抬高型心肌梗死患者(ST-segment elevation myocardial infarction,STEMI)接受 PPCI治疗后,1年内仍有9%病死率及和10%的心力衰竭发病率。之所以出现这样的结果,主要是忽于心肌 再灌注损伤的防治。目前,有许多防治心肌再灌注损伤的治疗手段,大多数都是通过减少心肌梗死面积而起 效的,但这一系列的研究均为小样本临床研究,缺乏大规模临床研究证据:本综述就临床上心肌缺血再灌注 损伤餉防治研究现状和进展进行总结分析。 【关键词】急诊PCI;缺血再灌注损伤;心肌 【中图分类号】R542.2+2;R541.4 DOI:10.13820/https://www.360docs.net/doc/63445140.html,ki.gdyx.20171290 对于急性ST段抬高型心肌梗死(STEMI)患者而言,及早实施经皮冠状动脉介入术(PPCI)恢复心肌灌注是减少心肌梗死面积、维持左室收缩功能以及预防心力衰竭发生的最有效的治疗措施。心肌再灌注治疗的首要目的是救活尚存活的心肌细胞。然而,恢复冠脉血流的过程似乎会加重心肌损伤及心肌细胞的死亡,因此降低了心肌再灌注治疗的益处— —这种现象被称为心肌再灌注损伤[1'2]o事实上,就单纯的心肌再灌注治疗效果而言,能够减少50%的心肌梗死面积,这也意味着心肌再灌注损伤所致的心肌梗死面积可达到最终心肌梗死面积的50%。虽然随着支架介入治疗、新型抗血小板制剂(列如普拉格雷、替格瑞洛、阿昔单抗)、抗血栓制剂(后者能够维持血流动力学的稳定)的发展,心肌再灌注治疗已经得到了很大程度的优化,但对于接受PPC1治疗的患者,仍然无明确有效的能够预防心肌再灌注损伤的治疗措施。对于STEMI患者,心肌梗死的面积与左室重塑、心力衰竭的发生以及PPCI的预后呈明显的负相关13-41 0Larose等⑸研究表明心肌梗死面积超过左心室总面积的23%的患者,不良心血管事件的发生率明显增加。在本文中,我们回顾临床上心肌保护方面的研究所面临的问题,并且重点阐述防治PPCI过程中出现心肌再灌注损伤的前景。 1心肌保护相关的研究所面临的问题 目前把对患者有益的新型保护心肌的治疗方法应用于临床当中有着极大的困难。大量研究发现心 *国家自然科学基金项目(编号:81600283),广西自然科学基金项目(编号:2016GXNSFBA380022) △通信作者。E-mail:403272754@https://www.360docs.net/doc/63445140.html, 【文献标志码】A 肌再灌注损伤后,有着许多信号通路的激活,这些研究中也阐明了这些通路是如何再灌注时起到心肌保护作用的[6_7]O同时这些研究也给出了一些药理学分子靶向治疗心肌再灌注损伤的机制。在过去的30-40年间,动物学实验中也证实了能够预防再灌注损伤及减少心梗面积的治疗方案(例如:抗氧化剂、钙通道阻滞剂、抗炎药、促红素、阿托伐他汀以及腺昔),但这些方案在临床工作中未得到令人满意的结果。最近也有许多研究提出一些新的降低心肌梗死面积的治疗方案(例如:低温治疗,调节线粒体功能,调控NO信号传导通路),但也都没有达到预期效果。导致这样结果的主要原因是动物试验中应用的动物心肌梗死模型与临床实际工作中的心肌梗死患者有很大的差别;另一种原因是临床试验设计不够充分Z。 随着PPCI治疗STEMI患者的临床疗效的持续提升,要证明一个新的保护心肌、减少心肌梗死面积的治疗方案是否有效则越来越困难。虽然目前心肌梗死患者的病死率较以前明显下降,但是心肌梗死患者心力衰竭发生率却越来越高。因此,仍需要探索新的、能够预防心肌再灌注损伤及减少心梗面积且可以维持左室收缩功能和预防心力衰竭的发生的治疗方案。 2减少心肌梗死面积的治疗方案 2.1缺血性后适应通过减少心肌再灌注、限制心 梗面积扩大的动物学试验中证实通过减少心肌再灌注引起几次短时期的心肌缺血具有预防心肌再灌注损伤及减少心肌梗死面积的功效,这种现象被称为心肌缺血预适应(ischemia preconditioning,IPC)0 Staat等学者将IPC应用于临床实践,并证实了该方法能够减少PPCI术后STEMI患者36%心肌梗死面
心肌缺血再灌注损伤的免疫学机制研究进展
心肌缺血再灌注损伤的免疫学机制研究 进展 夏霓,程翔(华中科技大学同济医学院附属协和医院 心内科,武汉 430022) 基金项目:国家自然科学基金项目(81170303和81222002);国家基础研究项目(973项目,2013CB531100);新 世纪优秀人才支持计划(NCET-09-0380)通讯作者:程翔 Email :nathancx@https://www.360docs.net/doc/63445140.html, 急性心肌梗死(acute myocardial infarction ,AMI )后,早期而有效的再灌注治疗是减小梗死面积并改善临床预后最有效的手段。然而,恢复缺血区的血流灌注会使再灌注前尚有活力的心肌细胞死亡,原有的缺血性损伤加重,导致梗死面积增大,被称为心肌缺血再灌注损伤(myocardial ischemia reperfusion injury ,MIRI )[1]。由于MIRI 的存在,AMI 后尽管得到最佳再灌注治疗仍有接近10%的患者发生死亡,而AMI 后心力衰竭的发生率也高达25%[2]。缺血的心肌恢复血流后,由于补体的激活和氧自由基的大量产生,白细胞被迅速募集到心肌,产生蛋白水解酶和氧自由基,从而导致损伤的发生发展,因此抗原非依赖性的天然免疫应答被认为是MIRI 的重要特征[3]。最近,作为主要介导适应性免疫应答的T 细胞和B 细胞被研究证实参与了各种器官的缺血再灌注损伤[4],并在MIRI 中日益受到关注。本文就MIRI 中的免疫炎症机制进展作一简要回顾。 尽管没有感染,缺血再灌注与对抗微生物入侵的免疫反应激活有很多共同之处。在缺血再灌注中的无菌炎性反应涉及危险相关分子模式(danger-associated molecular pattern ,DAMP )及其所激活的Toll 样受体(Toll-like receptor ,TLR ),天然免疫细胞的募集和激活以及适应性免疫系统的激活。1 天然免疫应答 1.1 TLR TLR 最早被认为识别病原相关分子模式(pathogen-associated molecular ,PAMP ),作为 抵抗病原微生物入侵的第一道防线;然而随后的研究表明,许多没有病原体的疾病TLR 也参与其中。组织损伤释放的内源性配体,即DAMP 可以识别TLR ,从而启动天然免疫应答[5]。在MIRI 中,不同阶段所激活的TLR 信号通路所起的作用也不尽相同。用TLR4[6]或TLR2[7]的配体预处理小鼠,能通过激活促生存的信号通路减轻随后的缺血再灌注损伤。Dong 等[8]的研究更证明TLR2–TIRAP-依赖性的信号通路介导缺血预适应。这些研究证实早期的TLR 信号通路激活在MIRI 中发挥保护作用。然而TLR 的持续激活则被证实有害,说明其在MIRI 中的双重作用。TLR2[9],TLR4[10]或MyD88[11]遗传缺陷的小鼠均表现为炎性反应和氧化应激受到抑制,梗死面积减小。TLR2的拮抗性抗体OPN-301[9]或脂多糖(LPS )与TLR4的竞争性抑制剂eritoran [12]均能减轻MIRI ,提示TLR2和TLR4可作为MIRI 的潜在性治疗靶点。1.2 补体 补体系统可通过3条途径被激活,分别为经典途径、凝集素途径和旁路途径。这3条通路最终都导致C3的降解,C5的激活,形成膜攻击复合体。C5a 是中性粒细胞的强趋化因子,通过上调CD11b/CD18(Mac-1)的表达,诱导中性粒细胞和内皮的牢固黏附以及随后的中性粒细胞穿内皮移行[13]。C5a 还能通过上调超氧化物增强中性粒细胞的氧化应激[14]。在心肌缺血性疾病中,对补体系统的研究最早开始于1971年的大鼠AMI 模型[15,16]。补体激活的3条途径都参与了MIRI 的疾病进程,而最近的研究表
心肌缺血再灌注损伤的发病机制.
心肌缺血再灌注损伤的发病机制 摘要21世纪是PCI的时代,PCI的发展与推广降低了ST段(STEMI)及非ST抬高(NSTEMI)性心肌梗死患者死亡率[1,2]、缩小了梗死的面积[3]、改善了左室的收缩功能[1,4],但是这种不断进步发展的PCI技术却不能显现出该技术当初刚用于临床时的降低心肌梗死患者的死亡率。因为研究人员们发现,某些患者就算开通了梗死的冠状动脉相关血管支配的心肌梗死面积却没有如人所愿的大大降低,心肌梗死的面积在开通冠脉后仍然在进展。因为研究人员发现缺血期的心肌在各种因素的作用下已经发生了损伤,心肌的再灌注有可能加重了缺血期心肌的损伤程度,对细胞或者细胞器造成了新的损伤,我们称之为再灌注损伤(reperfusion injury)。本文主要此种损伤的可能发生机制进行综述。 关键词心肌再灌注损伤心肌缺血心肌梗死炎症自由基线粒体渗透性转换孔 一、心肌再灌注损伤病理生理 心肌再灌注损伤(myocardial reperfusion injury)指的是缺血的心肌组织恢复血运后可能对心肌造成的进一步损伤[2,3]。但是缺血再灌注引起的心肌损伤的确切病理生理机制仍没有研究清楚。其中一个很重要的因素就是目前所建立使用的心肌缺血-再灌注模型本身就是一个问题,因为我们知道心肌的缺血分很多种,其中最常见也是最凶险的一类便是ST段抬高型心肌梗死,现流行的线栓建模法被广泛应用,但心肌梗死的过程却没有线栓法阻断冠脉引起的心肌组织坏死及再灌注损伤如此简单,根据欧美国家的指南[4,5],将心肌梗死分为五型,从这五种分型可以发现简单的结扎、再通冠脉造成的心肌梗死模型也不过是其中分型的一型,其它四型或者更多的类型心肌在缺血及再灌注时发生的确切变化仍然没有研究清楚的,因为我们至今没有发现哪一种干预措施可以非常有效的缩小心肌梗死后心肌坏死的进展。但是自50多年前,Jennings等[6]第一次从犬的缺血后再灌注的心脏组织中发现
心肌缺血再灌注损伤介绍和实验设计
心肌缺血再灌注损伤介绍与实验设计 Ⅰ、心肌缺血再灌注损伤: 它就是指缺血心肌组织恢复血流灌注时,导致再灌注区心肌细胞及局部血管 网显著的病理生理变化,这些变化共同作用可促使进一步的组织损伤。那这里的 关键词就就是缺血心肌组织。那为什么会产生缺血的心肌组织呢?这就与临床上 的疾病有关了。一些心脏疾病,比如急性心肌梗死、冠心病等她们会使心脏发生 缺血的症状,其基本的生理过程就就是心肌缺血。 Ⅱ、心肌缺血的危害: 心肌缺血:指单位时间内的冠脉血流量减少,供给组织的氧量也减少,缺血必定存在缺氧表明缺血缺氧。心肌缺血比单纯性心肌缺氧无血流障碍要严重,因为前者除了缺氧的影响之外,缺血组织也不能获得足够的营养物质又不能及时清除 各种代谢产物带来的有害影响。 一、心肌缺血的原因主要分为两种情况:1就是冠脉血流量的绝对不足。这种情况就是由自身疾病产生的,主要包括冠状动脉阻塞,冠状动脉痉挛。2就是冠脉血流量的相对不足:包括供氧降低或耗氧增加,比如高原高空或通风不良的矿井吸入氧减少;肺通气或换气功能障碍,可致血氧含量降低红细胞数量与血红蛋白含量 减少等。 二、缺血对心肌的危害主要包括以下几个方面:1就是心肌收缩能力降低。2就是导致心肌舒张功能降低。3就是心肌组织的血流动力学发生改变,比如说血流的阻力增加等。4就是心肌电生理的变化,比如说静息点位降低,传导速度减慢;室颤阈降低等。5就是导致心肌形态学的改变。当然还有其她的危害,在这里就不一一列举了。 由于心肌缺血存在这么多的危害,临床上针对这一疾病采取了再灌注治疗方法,但随之而来的又就是另外一个临床问题:缺血再灌注损伤。 下面具体介绍一下心肌缺血再灌注损伤。心肌缺血再灌注损伤英文缩写为MIRI,最早由詹宁斯等于1960年提出,发现其临床表现为再灌注心律失常、心肌 顿抑、心肌能量代谢障碍等现象。随后又有学者在临床手术中也证实了这一观点,发现在冠脉搭桥术完成后,心肌坏死进一步加重的现象。接着布朗沃尔德教授在1985年提出了这样一个观点:心肌再灌注就是一把双刃剑,既可以损伤心肌也能
心脏缺血-再灌注损伤的变化
(一)心功能变化 1.缺血-再灌注性心律失常在心肌缺血-再灌注过程中出现的心律失常称为再灌注性心律失常(reperfusion,arrhythmia)其中以室性心律失常,如室性心动过速和心室颤动最为多见。缺血时间过短,心肌损伤不明显;缺血时间过长,心肌丧失电活动,二者均不易出现缺血-再灌注性心律失常。此外,心律失常的发生还与缺血心肌的数量、缺血的程度、再灌注血流的速度及电解质紊乱等因素有关。缺血-再灌注性心律失常的发生机制:自由基和钙超载造成,的心肌损伤及ATP减少使ATP敏感性钾通道激活等均可改变心肌电生理特性;增多的儿茶酚胺刺激;受体,可提高心肌的自律性;缺血-再灌注使纤颤阈降低;心肌电解质紊乱,均可导致心律失常的发生。 2.心肌舒缩功能降低缺血-再灌注导致的心肌可逆性或不可逆性损伤均造成心肌舒缩功能降低,表现为心输出量减少,心室内压最大变化速率(±dp/dtmax)降低,左室舒张末期压力(LVEDP)升高等。1982年Braunwald 和Kloner首先用心肌顿抑(myocardialstunning)一词来描述心肌并未因缺血发生不可逆损伤,但在再灌注血流已恢复或基本恢复正常后一定时间内心肌出现的可逆性收缩功能降低的现象,以与心肌坏死、持续缺血或其他非缺血性因素引起的心功能障碍相区别。随着临床上溶栓治疗、心绞痛缓解、PTCA、冠状动脉搭桥术及心脏移植的开展,短期可逆性心肌缺血所导致的较长时间的心功能抑制愈来愈受到关注,其诊断及治疗直接影响到患者的预后。目前认为,心肌顿抑是缺血-再灌注损伤的表现形式之一,自由基爆发性生成和钙超载是心肌顿抑的主要发病机制。(二)心肌代谢变化缺血期心肌ATP及磷酸肌酸含量降低,ADP、AMP 及其降解产物含量升高。如缺血损伤轻,心肌高能磷酸化合物含量可较快恢复正常。如缺血时间较长,再灌注后心肌高能磷酸化合物含量不仅不回升,反而进一步降低。其机制:再灌注时自由基和钙超载等对线粒体的损伤使心肌能量合成减少;加之再灌注血流的冲洗,ADP、AMP等物质含量比缺血期降低,造成合成高能磷酸化合物的底物不足。(三)心肌超微结构的变化缺血-再灌注损伤时,心肌超微结构变化较单纯缺血时进一步加重,表现为细胞膜破坏,线粒体肿胀、嵴断裂、溶解、空泡形成,由于Ca2+蓄积,基质内致密颗粒增多,肌原纤维断裂、节段性溶解和出现收缩带。缺血-再灌注还可造成不可逆性损伤,出现心肌出血、坏死。
缺血后适应对心肌缺血/再灌注损伤的影响及其进展
中国循证心血管医学杂志2018年4月第10卷第4期 Chin J Evid Based Cardiovasc Med,April,2018,Vol.10,No.4 ? 506 ?? 综述 ? 缺血后适应对心肌缺血/再灌注损伤的影响及其进展 张文1,杨巍1 作者单位:1 150001 哈尔滨,哈尔滨医科大学附属第一医院心内科 通讯作者:杨巍,E-mail:yangwei@https://www.360docs.net/doc/63445140.html, doi:10.3969/j.issn.1674-4055.2018.04.34 1 心肌缺血/再灌注损伤 对于急性ST段抬高型心肌梗死(STEMI)患者,采用经皮冠状动脉介入治疗(PCI)开通罪犯血管及时恢复血流灌注,能降低梗死面积、保护心室功能,但在1年内仍有7%的死亡率和22%的心力衰竭(心衰)发生率,当高风险的STEMI发生心源性休克时,1年内的死亡率会更高,可达12%[1,2]。虽然心肌再灌注是挽救濒死心肌的有效方法,但是,在此过程的早期阶段又造成了心肌的额外损伤,比如扰乱离子稳态、活性氧的过量产生、激活炎症反应、引起线粒体功能紊乱和钙超载等,从而降低了心肌再灌注的获益,这一现象称为——心肌缺血/再灌注(I/R)损伤[3-6]。大多数学者认为即使单纯的PCI恢复心肌灌注后可以降低心肌梗死面积,但再灌注损伤仍可占总梗死面积的50%[5]。而且,随着科技的发展,虽然新型抗血小板、抗凝药物的应用改善再灌注血流,但对于PCI术后的患者再灌注损伤无明显作用。对于患者,其心肌梗死面积与心肌左室重塑、心衰及PCI术后的预后紧密联系。Larose 等表示当心肌梗死面积≥左室面积的23%时,易发生心衰等恶性心血管事件[7]。如何降低心肌I/R损伤并改善预后成为研究热点。通过大量的动物试验,发现了许多具有预防心肌再灌注损伤的措施,如抗氧化剂、镁剂、钙通道抑制剂、抗炎药物、阿托伐他汀、极化液、腺苷等,但这些方法从实验转化到临床应用并获利于患者却非常困难。2 缺血后适应及其临床应用 有研究首次报道在再灌注开始时迅速进行几个短暂的缺血与再灌注的循环可以降低狗的心肌梗死面积[8] 。之后这项发现迅速被用于多种实验模型,发现通过短时间的心肌缺血打断再灌注过程来调节心肌缺血环境,增加心肌对缺血/再灌注损伤的耐受能力,可显著地保护心肌细胞避免缺血/再灌注损伤,该现象称为缺血后适应(IPostC) [9,10]。 Staat等将这种方法迅速用于临床,并表明缺血后适应,即在罪犯血管用四个1 min缺血和1 min灌注的循环,在STEMI 患者PCI术后可以使心肌梗死面积降低36%[11]。另外,有研究表明在冠状动脉完全闭塞(TIMI 0级)的STEMI患者中,IPostC处理后患者获益最大[12] ,但是其保护机制尚不清楚,考虑到临床手术的可操作性及临床效果的显著性,对IPostC 减轻I/R损伤的机制还需进一步研究。 3 缺血后适应对心肌缺血/再灌注损伤的作用及其机制3.1 IPostC通过促进自噬降低心肌I/R损伤 自噬是吞噬自体胞浆内老化或损伤的蛋白及细胞器并使其包被进入囊泡, 与溶酶体融合形成自噬溶酶体,降解其所包裹内容物的过程,从而完成细胞自体的新陈代谢和一些细胞器的更新[13]。一些研究表明自噬在心肌I/R损伤中发挥重要作用。当在可调控的范围内促进自噬可以在I/R损伤中补偿线粒体损伤和建立蛋白质稳态[14]。Hao等通过动物模型发现,IPostC和I/R 组相比较,心肌梗死面积降低,心肌结构紊乱、细胞间质水肿等减轻,保留了心肌的正常结构,而且线粒体碎片的量及体积的减小程度都显著改善,而当给予I/R心肌IPostC 加上自噬抑制剂处理时,IPostC的这种心肌保护作用就消失了,出现心肌细胞坏死、细胞核溶解和显著的心肌结构紊乱[15]。由此可见自噬可以被IPostC所调控,并且参与了IPostC的心肌保护机制。另外,还有报道称IPostC可以通过促进自噬减轻氧化应激从而抑制再灌注损伤[16],其具体机制还需进一步研究。 3.2 IPostC通过调节神经元型一氧化氮合酶途径降低心肌I/R 损伤 心肌I/R损伤机制中氧化应激和钙超载是主要途径[2],主要通过产生过量内源性的活性氧和氮氧化物发挥作用[17]。虽然一氧化氮(NO)在再灌注心肌保护的方法如缺血预适应和IPostC中都发挥着重要作用,但实际上NO在心肌I/R损伤中是一把双刃剑[18]。NO在经典的环磷酸鸟苷环化酶(cGMP)介导的信号通路中发挥作用,但最近的研究表明NO调节的心肌功能也受到氮氧化物酶系统的限制[19]。内皮型一氧化氮合酶(eNOS)位于细胞膜小凹,调节细胞膜L型钙通道,神经元型一氧化氮合酶(nNOS)位于肌浆网和线粒体中,通过维持钙循环和亚硝基——氧化还原反应来调节肌浆网和线粒体的功能[20-22]。有报道称nNOS大量表达可以保护小鼠心肌避免I/R 损伤,但是,同时也增加了小鼠室性心律失常和心肌梗死后的死亡率[23]。Hu等在离体小鼠心脏中发现,IPostC组与I/R组相较而言,IPostC显著促进了左室收缩力的恢复,降低了左室舒张末压水平和血浆中乳酸脱氢酶(LDH)水平,而nNOS 抑制剂却能消除IPostC的心肌保护能力[24],提示IPostC通过调节nNOS途径发挥心肌保护作用。 3.3 IPostC通过下调钙敏感受体降低心肌I/R损伤 钙敏感受体(CaSR)调节机体多个组织和器官的钙代谢平衡[25]。在2003年首次报道了CaSR存在于小鼠心肌组织[26]。CaSR 在心肌组织中为G蛋白偶联受体,可以通过激活磷脂酶C (PLC)促进3-磷酸磷脂酰肌醇(IP3)的产生,进而促进肌浆网中钙离子释放入线粒体内,通过启动线粒体和肌浆网的凋亡通路诱发心肌凋亡[27]。曾有报道称在心肌I/R损伤过程中,CaSR过度表达,引起钙超载促进心肌细胞凋亡,而IPostC可以下调CaSR的表达起到心肌保护作用[28,29]。随后实验中发现,IPostC通过下调CaSR通路的心肌保护作用离不开ATP敏感性钾通道(K +-ATP),是多种心肌保护方法如缺血预适应、远程预处理等过程中最后的效应受体,而K +-ATP通道的开放则是CaSR下调后的下游效应[29,30]。在心 【中图分类号】R542.2 【文献标志码】 A 【文章编号】1674-4055(2018)04-0506-02