利用染色体片段置换系定位水稻抗纹枯病QTLs

遗传学名词解释
遗传学名词解释 11、性状:生物体或其组成部分所表现的形态、生理或行为特征称为性状(character/trait) 13、相对性状:不同生物个体在单位性状上存在不同的表现,这种同一单位性状的相对差异 称为相对性状 14、显性(dominate)性状:在子一代中出现来的某一亲本的性状。 15、隐性 (recessive)性状:在子一代中未出现来的某一亲本的性状。 17、基因型(genotype):指生物个体基因组合,表示生物个体的遗传组成,又称遗传型; 18、表现型(phenotype):指生物个体的性状表现,简称表型。 19、纯合基因型:具有一对相同基因的基因型称为纯合基因型(homozygous genotype),如 CC和cc;这类生物个体称为纯合体(homozygote)。 ●显性纯合体(dominant homozygote), 如:CC. ●隐性纯合体(recessive homozygote), 如:cc. 21、基因的分离定律:一对等位基因在杂合体中各自保持其独立性,在配子形成时,彼此分 开,随机地进入不同的配子,在一般情况下:F1杂合体的配子分离比 为1:1,F2表型分离比是3:1,F2基因型分离比为1:2:1 22、测交(test cross)法:即把被测验的个体与隐性纯合亲本杂交,根据侧交子代(Ft)的 表现型和比例测知该个体的基因型。 23、独立分配定律:支配两对(或两对以上)不同性状的等位基因,在杂合状态时保持其独 立性。配子形成时,各等位基因彼此独立分离,不同对的基因自由组合。 24、系谱分析法:用图解表明一个家族中某种性状(或遗传疾病)发生的情况,进而判断该 性状(或遗传疾病)的遗传方式。 27、外显率(penetrance):指在特定环境中,某一基因型(常指杂合子)个体显示出预期表型 的频率(以百分比表示)。就是说同样的基因型在一定的环境中有的 个体表达了,而有的个体可能没有表达,这样外显率就小于100% ——不完全外显。外显率为100%——完全外显 28、表现度(expressivity):是指具有相同基因型的个体之间基因表达的变化程度。 29、共显性/并显性:一对等位基因的两个成员在杂合体中都表达的遗传现象。 30、镶嵌显性:由于等位基因的相互作用,双亲的性状在子代同一个体的不同部位表现的镶 嵌图式。 31、隐性致死基因:在杂合时不影响个体的生活力,但在纯合时有致死效应的基因。 32、显性致死基因(dominant lethal gene):在杂合状态下即表现致死作用的致死基因 33、复等位基因:在群体中占据某同源染色体同一座位的两个以上的决定同一性状的基因 34、基因互作:基因在决定同一生物性状表现时,所表现出来的相互作用。 35、互补基因:两对非等位的显性基因同时存在并影响生物的某同一性状时才使之表现该性 状,其中任一基因发生突变都会导致同一突变性状出现,这类基因称为互补基因。 37、叠加效应:不同基因对性状产生相同影响,只要两对等位基因中存在一个显性基因,表 现为一种性状;双隐性个体表现另一种性状;F2产生15:1的性状分离比例。 这类作用相同的非等位基因叫做叠加基因 38、上位效应:影响同一性状的两对非等位基因中的一对基因(显性或隐性)掩盖另一对显 性基因的作用时,所表现的遗传效应称为上位效应,其中的掩盖者称为上位 基因,被掩盖者称为下位基因。 39、显性上位:在上位效应中,起掩盖作用的是一个显性基因,使另一个显性基因的表型被 抑制,孟德尔F2表型比率被修饰为12:3:1
遗传学课后答案
第四章连锁遗传和性连锁 1.试述交换值、连锁强度和基因之间距离三者的关系。 答:交换值是指同源染色体的非姐妹染色单体间有关基因的染色体片段发生交换的频 率,或等于交换型配子占总配子数的百分率。交换值的幅度经常变动在0~50%之间。交换值越接近0%,说明连锁强度越大,两个连锁的非等位基因之间发生交换的孢母细胞 数越少。当交换值越接近50%,连锁强度越小,两个连锁的非等位基因之间发生交换的孢母细胞数越多。由于交换值具有相对的稳定性,所以通常以这个数值表示两个基因在同一染色体上的相对距离,或称遗传距离。交换值越大,连锁基因间的距离越远;交换值越小,连锁基因间的距离越近。 2.在大麦中,带壳(N)对裸粒(n)、散穗(L)对密穗(l)为显性。今以带壳、散穗与 裸粒、密穗的纯种杂交,F1表现如何?让F1与双隐纯合体测交,其后代为: 带壳、散穗201株裸粒、散穗18株 带壳、密穗20株裸粒、密穗203株 试问,这2对基因是否连锁?交换值是多少?要使F2出现纯合的裸粒散穗20株,至少应中多少株? 答:F1表现为带壳散穗(NnLl)。 测交后代不符合1:1:1:1的分离比例,亲本组合数目多,而重组类型数目少,所以这两对基因为不完全连锁。 交换值%=((18+20)/(201+18+20+203))×100%=8.6% F1的两种重组配子Nl和nL各为8.6%/2=4.3%,亲本型配子NL和nl各为(1-8.6%)/2=45.7%; 在F2群体中出现纯合类型nnLL基因型的比例为: 4.3%×4.3%=18.49/10000, 因此,根据方程18.49/10000=20/X计算出,X=10817,故要使F2出现纯合的裸粒散穗20株,至少应种10817株。 3.在杂合体ABy/abY,a和b之间的交换值为6%,b和y之间的交换值为10%。在没有 干扰的条件下,这个杂合体自交,能产生几种类型的配子;在符合系数为0.26时,配子的比例如何? 答:这个杂合体自交,能产生ABy、abY、aBy、AbY、ABY、aby、Aby、aBY8种类型 的配子。 在符合系数为0.26时,其实际双交换值为:0.26×0.06×0.1×100=0.156%,故其配子的比例为:ABy42.078:abY42.078:aBy2.922:AbY2.922:ABY4.922:aby4.922:Aby0.078:aBY0.078。 3.设某植物的3个基因t、h、f依次位于同一染色体上,已知t-h相距14cM,现有如下杂 交:+++/thf×thf/thf。问:①符合系数为1时,后代基因型为thf/thf的比例是多少?②符合系数为0时,后代基因型为thf/thf的比例是多少? 答:①1/8②1/2 5.a、b、c3个基因都位于同一染色体上,让其杂合体与纯隐性亲本测交,得到下列结果: +++74 ++c382 +b+3 +bc98 a++106
染色体
一.名词解释(每题2分,共30分) B染色体:有些生物的细胞中出现的额外染色体 双受精:1个精核(N)与卵细胞(N)结合为合子(2N),将来发育成胚。另一精核(N)与两个极核(N+N)受精结合为胚乳核(3N),将来发育成胚乳的过程 体细胞联会:在一些动植物的体细胞有丝分裂中观察到一些非随机分布现象,其同源染色体在空间分布上有相互靠近的倾向,出现同源染色体紧密、平行配对的现象。 花粉直感:如果在3n胚乳的性状上由于精核的影响而直接表现父本的性状,这种现象称为胚乳直感或花粉直感。 遗传图谱:将一对同源染色体上的各个基因的位置确定下来,并绘制成图叫做遗传图谱 结构异染色质:除复制期外在细胞的所有时期都保持聚缩状态的染色质 假连锁:两对染色体上原来不连锁的基因,由于靠近易位断点,易位杂合体总是以相间分离方式产生可育的配子,因此就表现出假连锁现象。 染色体干扰:一个交叉的发生对第二个或以后的其他交叉会有一定的影响 复合易位:涉及染色体的三次以上的断裂,各染色体间发生复杂的相互易位。 端粒:末端特化的染色较深的部位,是染色体的天然末端,对染色体的稳定起着非常重要的作用 异源多倍体:指不同物种杂交产生的杂种后代经过染色体加倍形成的多倍体。 NOR(核仁组织区):是染色体上与核仁形成有关的区域。也叫次缢痕。 染色体消减:指双翅类、膜翅类等昆虫的胚在特定的卵核分裂中,部分染色体不进入核而丢失的现象 孪生现象:有丝分裂发生联会,产生交叉互换,使隐性基因重合 孤雄生殖:在正常的有性胚囊中,卵细胞未经受精而直接发育成胚的现象 单端单体:当某对染色体缺失但保留其1条染色体臂的端着丝粒染色体 细胞融合:指人工的或自然发生的细胞合并形成多核细胞的现象 部分同源染色体:来源相同,具有部分相同的基因和序列的染色体 染色体消减:指双翅类、膜翅类等昆虫的胚在特定的卵核分裂中,部分染色体不进入核而丢失的现象
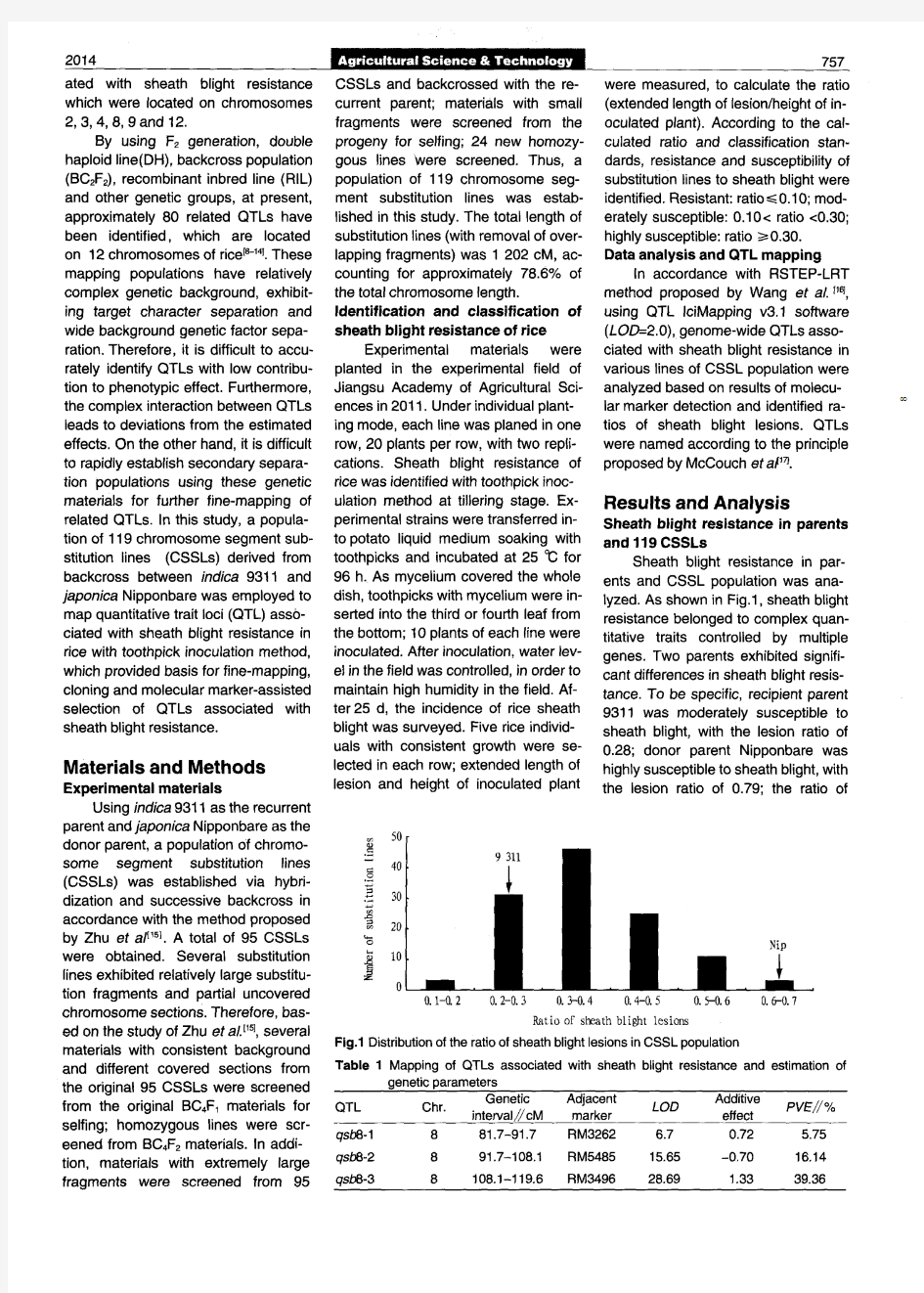
水稻染色体片段代换系群体的构建及应用研究进展
水稻染色体片段代换系群体的构建及应用研究进展 徐建军,梁国华 * (扬州大学,江苏省作物遗传生理国家重点实验室培育点/植物功能基因组学教育部重点实验室,江苏扬州225009) 摘要 定位和克隆水稻重要农艺性状QTL ,是水稻功能基因组学研究的重要方向,是分子标记辅助选择选育高产、优质、多抗水稻新品种的重要基础。染色体片段代换系是进行QTL 分析的理想材料。介绍了水稻染色体片段代换系群体的构建原理,综述了其构建及应用研究进展,并对其研究方向进行了展望。 关键词 水稻;染色体片段代换系;构建;应用;进展 中图分类号 S 511 文献标识码 A 文章编号 0517-6611(2011)04-01935-04 R esearch Progress of Constructi on a nd Applicatio n of R i ce (Or yza sati va L .)Chro mos o m e Seg men t Substituti on L i nes XU Jian -jun et al (Ji ang s u K ey Laboratory of CropG enetics and Physi o l ogy /K ey L aboratory o f theM i nistry ofEducati on for P lant Functi on -a lG enom i cs ,Y ang z hou Un i versit y ,Y angz hou ,Ji angs u 225009)Abstract The fi ne m apping and cl oni ng quantitati ve tra it loc i (QTL s)res ponsi b l e for traits of agrono m i c m i portance i n rice i s kno wn as the most general stategy i n pl ant genom ics and prov i des a f ounda ti on t o select new rice vari e ties whit h h i gh y i e l d qua lity ,resistance via marker -as -sisted selecti on (MA S)i n rice breedi ng prog ra m s .Chro moso m e segment substituti on li nes(CSSLs)are one o f t he most po w erful too ls for the det ecti on and prec i se mappi ng ofQTL s .I n t h i s paper ,t he pri nci p l e of deve l op i ng t he CSS L swas descr i bed ,t he research progress of consturcti on and applicati on was rev i ewed ,and the pros pectwas discussed .K ey words R i ce ;Chromoso m e seg m ent s ubstit uti on li nes(CSSLs);Constructi on ;Applicati on ;Progress 基金项目 国家重大基础研究发展规划项目(2005CB120807)。 作者简介 徐建军(1983-),男,湖南武冈人,博士研究生,研究方向: 水稻遗传育种和功能基因组学。*通讯作者。 收稿日期 2010-11-10 水稻是世界上重要的粮食作物之一,为世界1/2以上的人口提供食物和营养来源[1] 。水稻是我国最重要的粮食作物,水稻产量占我国粮食总产的1/2以上,对确保我国粮食安全和农业可持续发展具有举足轻重的意义。此外,水稻以其较小的基因组、成熟的遗传转化体系、高密度的连锁图谱、全基因组已经测序完成等优势,已成为单子叶植物遗传研究的模式植物[2] 。因此,大规模地开展控制水稻重要农艺性状基因的定位和克隆研究,不仅是水稻功能基因组学研究的重要内容,更是水稻产量提高、品质改良的重要途径。 水稻产量、品质、抗性等许多重要性状,都属于微效多基因控制的数量性状。QTL 的定位和克隆,是挖掘和利用新基因资源的一个非常有效的途径,可以使育种家直接选择和操作控制数量性状的基因型。目前,对水稻进行QTL 定位和克隆的常用的群体有F 2/F 3、BC l 、D H 、R I L 、N I Ls 和染色体片段代换系等。利用F 2/F 3、BC l 、DH 和R I L 等初级定位群体进行QTL 定位时,由于QTL 之间存在复杂的互作效应,对QTL 的效应估计不准确,对表型效应贡献少的QTL 常不能被准确鉴定出来,存在QTL 定位的准确性和精确性都不高的缺点。N I Ls 是目前应用较多的用来定位和克隆QTL 的群体,但是由于N I L s 的构建费时费力,其应用受到一定的限制[3] 。 Doi 等建议建立染色体片段代换系群体是解决上述问题、完成QTL 定位和克隆最有效的方法 [4] 。染色体片段代换 系是通过双亲杂交、回交和分子标记辅助选择选育的一整套供体代换片段覆盖受体全基因组的近等基因系。染色体片段代换系具有单一性、稳定性等特点。每一个染色体片段代换系只含有1个或几个来自供体亲本的染色体片段,利用其进行QTL 分析时,能够消除遗传背景的干扰,把复杂性状分 解为简单的孟德尔因子进行研究,提高QTL 鉴定的精确度和灵敏度;通过染色体片段代换系与轮回亲本杂交构建次级F 2 分离群体,可以对目标QTL 进行精细定位;染色体片段代换系属于永久性群体,可以提供大量的种子用于多点、多年、多重复的试验,可以研究QTL 之间的互作,以及QTL 与环境之间的互作。所以,建立染色体片段代换系对水稻功能基因组学的研究具有重要意义。 1 染色体片段代换系群体的构建原理 1.1 多态分子标记的发展 水稻染色体片段代换系是通过亲本杂交、多代回交和分子标记辅助选择建立的一系列近等基因系。在构建染色体片段代换系之前,首先,必须发展一批在水稻基因组上均匀分布的、达到一定覆盖密度的、在供体亲本与受体亲本之间有多态性的分子标记。 水稻是已经完成全基因组测序的物种,因此,基于已有的水稻遗传或物理图谱,发展基于PCR 的SS R 标记是最简单、实用的。SSR 标记以PCR 为基础,检测简单快速,多态性高,重现性好,标记信息可以直接从科技网站(h ttp ://www.gra m ene .or g /)上获取,为分子标记辅助选择,建立水稻染色体片段代换系奠定了基础。 1.2 分子标记辅助选择选育染色体片段代换系 首先,供体亲本与受体亲本杂交获得F 1;其次,以受体亲本作为轮回亲本,经过多代回交获得BC n F 1;再次,BC n F 1自交,利用分子标记辅助选择鉴定出含有1个或者几个受体亲本代换片段的单株;最后,利用这些单株自交获得染色体片段代换系。杂交的目的是导入供体亲本的染色体片段;回交的目的是重建受体背景,使代换系除了少数来自供体亲本的染色体片段外,遗传背景与受体亲本一致。理论上,经过2、3和4代回交,代换系来自受体亲本的遗传背景分别为75.00%、87.50%和93.75%。现阶段,不同的研究组在选育代换系时,进行分子标记辅助选择的代次有所不同。有的研究组从回交的低世代就开始进行分子标记辅助选择,有的则从高世代才开始,有的研究组在获得F 1之后经过多代自交再回交,然后才进行分子标记辅助选择。 安徽农业科学,J ournal of An hu iA gr.i S c.i 2011,39(4):1935-1938责任编辑 王淼 责任校对 李岩
利用单片段代换系研究水稻产量相关性状QTL加性及上位性效应
作物学报ACTA AGRONOMICA SINICA 2012, 38(11): 2007?2014 https://www.360docs.net/doc/9a18722656.html,/zwxb/ ISSN 0496-3490; CODEN TSHPA9E-mail: xbzw@https://www.360docs.net/doc/9a18722656.html, DOI: 10.3724/SP.J.1006.2012.02007 利用单片段代换系研究水稻产量相关性状QTL加性及上位性效应 赵芳明1张桂权2曾瑞珍2杨正林1凌英华1桑贤春1何光华1,* 1西南大学水稻研究所 / 转基因植物与安全控制重庆市重点实验室 / 南方山地农业教育部工程研究中心, 重庆 400716; 2华南农业大学广东省植物分子育种重点实验室, 广东广州 510642 摘要: 产量及其相关性状如单株有效穗数、千粒重、穗实粒数、穗总粒数和结实率等是水稻重要的农艺性状, 了 解产量及其相关性状QTL的加性及上位性效应对以分子标记聚合育种改良水稻产量具有重要意义。本文以16个单 片段代换系及15个双片段代换系分析了水稻产量相关性状QTL的加性及上位性效应。共检出影响产量及其相关性 状的13个QTL, 包括产量性状1个、单株有效穗数1个、千粒重4个、穗实粒数4个、穗总粒数2个和结实率1个, 分布于第2、第3、第4、第7和第10染色体上。此外, 检出12对双基因互作。结果显示, 2个正向(或负向)产量性 状QTL聚合, 往往会产生负向(或正向)的上位性效应, 能否产生更大(或更小)的目标性状, 取决于双片段遗传效应(加性效应与上位效应代数和)绝对值与单片段最大加性效应绝对值的差。本研究结果对实施高产分子标记聚合育种 方法有重要参考价值。 关键词: 水稻; 单片段代换系; 产量相关性状QTL; 加性效应; 上位性效应 Epistatic and Additive Effects of QTLs for Yield-Related Traits Using Single Segment Substitution Lines of Rice (Oryza sativa L.) ZHAO Fang-Ming1, ZHANG Gui-Quan2, ZENG Rui-Zhen2, YANG Zheng-Lin1, LING Ying-Hua1, SANG Xian-Chun1, and HE Guang-Hua1,* 1 Rice Research Institute, Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops / En-gineering Research Center of South Upland Agriculture, Ministry of Education, Chongqing 400716, China; 2 Guangdong Key Laboratory of Plant Molecular Breeding, South China Agricultural University, Guangzhou 510642, China Abstract: Yield-related traits such as panicle number per plant, thousand grain weight, number of grains per panicle, number of spikelets per panicle and seed setting rate are important agronomic traits in rice. Understanding additive and epistatic effects of QTL for yield-related traits are important to increase rice yield using method of pyramiding breeding with molecular marker as-sisted selection. In this paper, additive and epistatic effects of QTLs for rice yield-related traits were analyzed using 16 single segment substitution lines (SSSL) and 15 double segment substitution lines (DSSL). A total of thirteen QTLs for yield-related traits were identified on the chromosomes 2, 3, 4, 7, and 10 respectively, containing one for grain yield per plant (GY), one for panicle number (PN), four for thousand-grain weight (TGW), four for number of grains per panicle (NGP), two for number of spikelets per panicle (NSP) and one for seed-setting rate (SSR). Furthermore, twelve pairs of digenic interactions were detected for yield-related traits. The results showed that pyramiding two QTLs with positive effects (or two QTLs with negative effects) often results in negative epistatic effects (or positive epistatic effects) in DSSL. Whether larger or smaller value of yield-related traits is produced lies on the difference between the absolute value of genetic effect (algebraic sum of additive and epistatic effects) in the DSSL and the largest value of additive effect in the SSSL. These results are important to improve yield by pyramiding fa-vorable QTLs for yield-related traits. Keywords: Rice; Single segment substitution lines; QTLs for yield-related trait; Additive effects; Epistatic effect 单产的提高始终是水稻育种的追求目标, 而产量性状是数量性状, 遗传基础复杂, 易受环境和遗 本研究由西南大学基本科研业务费专项资金(XDJK2010B011)和重庆市自然科学基金项目(CSTC, 2010BB1131)资助。 *通讯作者(Corresponding author): 何光华, E-mail: hegh@https://www.360docs.net/doc/9a18722656.html, 第一作者联系方式: E-mail: zhaofangming2004@https://www.360docs.net/doc/9a18722656.html, Received(收稿日期): 2012-03-18; Accepted(接受日期): 2012-07-05; Published online(网络出版日期): 2012-09-10. URL: https://www.360docs.net/doc/9a18722656.html,/kcms/detail/11.1809.S.20120910.1328.007.html
例谈染色体结构变异交叉互换基因突变三者之间的辨析.
例谈染色体结构变异\交叉互换\基因突变三者之间 的辨析 生物的可遗传变异主要有三种情况构成,分别是染色体变异、基因重组、基因突变。其中染色体变异中的染色体结构变异、基因重组中的交叉互换、基因突变三者既有相似性,又有本质区别,现根据几个实例对这三种情况进行辨析。 1染色体结构变异与基因突变的辨析 【例1】(多选)已知某物种的一条染色体上依次排列着M、N、O、p、q五个基因,如图1所示,在细胞有丝分裂后出现了如图A-D变化所示的染色体,其中属于染色体结构变异的是() 解析:在此题中A图染色体片段缩短,基因数量明显减少,属于染色体结构变异中的缺失;B图中0基因数量增多属于染色体结构变异中的增添;C图中基因的数量种类都没有发生变化,但是各个基因在染色体上的位置发生了明显的变化,所以应属于染色体结构变异中的基因倒位。D图中基因的数量没有发生变化,各个基因的位置没有变,但是原图中N、o、p三个基因分别变成了n、o、P,出现了原染色体上没有的基因,应是基因突变或交叉互换。本题题干中已明确说明是发生在有丝分裂过程中,因此排除了交叉互换的可能。应为基因突变。 参考答案:ABC。 小结:在此题中要辨析染色体结构变异和基因突变之间的差异,主要是抓住两者的本质差异:染色体上有无新基因的产生。 注意:由于染色体结构变异中还有一种情况是易位,因此在利用此策略时还要注意染色体上产生的新基因是否是细胞内其他染色体上已经存在的。如果产生的新基因是在其他染色体上也有的,那么也不排除发生易位的可能。2染色体结构变异与交叉互换的辨析 【例2】图2是果蝇细胞的染色体组成,以下说法正确的是() A1与4之间的交换属于基因重组 B染色体1、2、4、5属于一个染色体组 C控制果蝇红眼或白眼的基因位于6号染色体上 D果蝇单倍体基因组可以由1、3、7、5、6组成
野生稻染色体片段代换系构建及其效应分析
材料创制与性状评价 Germplasm Innovation and Trait Evaluation 野生稻染色体片段代换系构建及其效应分析 张晨昕 邱先进 董华林余四斌* 华中农业大学作物遗传改良重点实验室,植物科学技术学院,武汉,430070*通讯作者,ysb@https://www.360docs.net/doc/9a18722656.html, 摘 要 通过回交程序结合分子标记辅助选择构建了一套染色体片段来源于马来西亚普通野生稻的珍汕 97B 染色体片段代换系。该套染色体片段代换系由105份材料构成,每系含有一个或少数几个导入片段,所 有导入片段相互衔接覆盖野生稻全基因组。染色体片段代换系的平均背景回复率为94.6%,平均导入片段长度为41.7cM 。利用该群体以及相同亲本的高世代BC 3F 3群体,共定位到40个QTL 影响抽穗期、株高、SPAD 值、有效穗数和穗长等农艺性状。该套野生稻染色体片段代换系为发掘和利用野生资源中的优良基因提供重要材料基础。 关键词 野生稻,染色体片段代换系,QTL Development and Characterization of Chromosome Segment Substitution Lines Using O.rufipogon as Donor Zhang Chenxin Qiu Xianjin Dong Hualin Yu Sibin * National Key Laboratory of Crop Genetic Improvement,College of Plant Science and Technology,Huazhong Agricultural University,Wuhan,430070*Corresponding author,ysb@https://www.360docs.net/doc/9a18722656.html, DOI:10.3969/mpb.008.001113 Abstract A set of chromosome segment substitution lines (CSSLs)has been developed by successive backcross be tween the common wild rice (O.rufipogon )from Malaysia as donor and Zhenshan97B as recurrent parent with molec-ular marker-assisted selection.This CSSLs population consists of one hundred and five lines.Each line carries one or a few segments.Mutual overlapping chromosomal segments have a complete coverage of the wild rice genome.The average genome proportion of recurrent parent is 94.6%.The average length of introgression segment is https://www.360docs.net/doc/9a18722656.html,ing BC 3F 3and CSSLs population,40QTLs were detected for five agronomic traits including heading date,plant height,SPAD value,panicle number per plant and panicle length.The developed CSSLs will provide an important source for identification of favorable gene from the wild rice and molecular breeding in rice.Keywords Wild rice,Chromosome segment substitution lines,QTL 分子植物育种,2010年,第8卷,第6期,第1113-1119页Molecular Plant Breeding,2010,Vol.8,No.6,1113-1119 基金项目:本研究由国家农业本研究由国家农业部948项目(2006-G1)和转基因生物新品种培育重大专项课题共同资助 水稻是最重要的粮食作物之一。全世界50%以上的人口都以它为主要食物来源。栽培稻受到长期的定向选育导致品种遗传基础变窄,其产量潜力及抵抗各种逆境等能力受到影响。野生稻具有丰富的遗传多样性,长期处于野生状态,经历生存竞争和自然选择,积累了大量的栽培稻品种所缺乏的有利基因(钟代彬等,2000)。例如,上世纪70年代,我国科学家从普通野生稻O.rufipogon 中找到了雄性不育细胞质源,实现了杂交水稻的三系配套(Lin and Yuan, 1980)。最近,研究者利用分子标记技术还发现普通野生稻中存在增加粒重和产量的基因(Xiao et al.,1998;Xie et al.,2006;2008). 本研究通过回交程序结合分子标记辅助选择将O.rufipogon (IRGC105491)染色体片段导入到珍汕97B (ZS97B)中,构建覆盖全基因组且相互重叠的染色体片段代换系,对部分农艺性状进行数量性状位点(quantitative trait loci,QTL)定位,为发掘和利用野生资源中的优良基因提供材料基础。
第九章 染色体畸变
第九章染色体畸变 第一节染色体结构的变异 一、染色体结构变异的机理 二、染色体结构变异的类型 三、染色体结构变异的应用 1927年穆勒(H.J.Muller)用X射线处理果蝇精子,发现:电离辐射 染色体结构变异。 自然辐射、温度、营养、生理环境等因素的异常变化,都可能使染色体发生断裂。 人为地用某些物理因素(如UV、x-ray、γ-ray、中子等)或化学试剂处理生物体、细胞,染色体断裂的频率还会大大提高。 结构变异的形成:断裂-愈合假说 自发或诱发情况下,染色体可能断裂,形成粘性末端(cohesive end),具有与另一个断裂末端愈合的能力,而不能稳定存在。 A.不愈合:保持断头,无着丝粒的染色体片段丢失,产生结构变异; B.重建:断裂末端自身愈合,恢复原状; C.非重建性愈合:错接,产生结构变异。 (一)缺失 (二)重复 (三)倒位 (四)易位 1、缺失的类型: (1)顶端缺失(末端缺失):丢失染色体某臂的外端。 (2)中间缺失:丢失染色体某臂内部的一段。 (3)整臂缺失:染色体也可能丢失整个一个臂,变成顶端着丝粒染色体和一个无着丝粒的染色体断片。 2、缺失的鉴定 一般采用缺失杂合体在减数分裂时染色体配对的图象来区别。 (1)中间缺失 (2)顶端缺失 (3)断片 (1)中间缺失: 缺失环:如果是中间缺失,且缺失区段较长,那么,缺失杂合体的一对染色体在粗线期可以联会,但正常染色体上多余的区段常呈环状突起。 中间缺失染色体没有断头外露,比较稳定,因而常见的缺失染色体多是中间缺失的。 (3)断片: 在最初发生缺失的细胞内,可见到遗弃在细胞质里无着丝粒的 断片。但随着细胞多次分裂,断片即消失。 3、缺失的遗传学效应 (1)出现异常或致死:丢失了基因,代谢过程可能部分地中断,使个体生活力降低、表型变化或不能成活。 发生缺失的配子常败育,特别是花粉,几乎不能存活,胚囊忍受缺失的能力略高于花粉,因此缺失多数是
【遗传学】第五章 染色体和连锁群
第五章染色体和连锁群 本章重点 一、连锁遗传: 二对性状杂交有四种表现型,亲型多、重组型少; 杂种产生配子数不等,亲型相等、重组型相等。 二、连锁和交换机理: 粗线期交换、双线期交叉,非姐妹染色体交换。 三、交换值及其测定: 重组配子数/总配子数; 测交法测定,也可用F2 材料进行估计。 四、基因定位和连锁遗传图: 确定位置、距离,基因位于染色体上; 二点测验、三点测验; 连锁群、连锁遗传图。 学时:9 1900年孟德尔遗传规律重新发现以后,生物界广泛重视,进行了大量试验。其中有些属于两对性状的遗传结果不符合独立分配规律→摩尔根以果蝇为材料进行深入细致研究→提出连锁遗传规律→创立基因论→认为基因成直线排列在染色体上,进一步发展为细胞遗传学。 第一节连锁和交换 一、连锁 (一)连锁现象的发现 1906年,贝特生(Bateson W.)和贝拉特(Punnett R. C.)在香豌豆的二对性状杂交试验中→首先发现性状连锁遗传现象。 第一个试验: P 紫花、长花粉粒(PPLL)×红花、圆花粉粒(ppll) ↓ F1紫、长PpLl ↓? F2紫、长紫、圆红、长红、圆 P_L_P_ll ppL_ppll总数 实际个体数4831390393 13386952 按9:3:3:1推算3910.51303.5 1303.5434.56952 上述结果进行X2检验时,X2=3371.58,说明实计数与预计数差异极其显著,不可能由随机误差造成,应作重复实验。 以上结果表明F2 :
①. 同样出现四种表现型; ②. 不符合9:3:3:1; ③. 亲本组合数偏多,重新组合数偏少(与理论数相比)。 第二个试验: P紫花、圆花粉粒(PPll)×红花、长花粉粒(ppLL) ↓ F1 紫、长PpLl ↓? F2 紫、长紫、圆红、长红、圆 P_L_P_ll ppL_ppll总数 实际个体数22695971419 按9:3:3:1推算235.878.578.526.2419 结果与第一个试验情况相同。 X2=32.40,证明它仍然显著不符合孟德尔规律。 从这两个实验的数据可以看出一种共同的倾向:即与自由组合定律所预期结果相比较,F2中性状的亲本组合类型远远多于重组组合的类型,这等于说,在F1杂种形成配子时两对基因可能发生的4种类型中,有更多保持亲代原来组合的倾向,而且这种倾向与显隐性无关。 连锁遗传:原来亲本所具有的两个性状,在F2连系在一起遗传的现象。 相引组:甲乙两个显性性状,连系在一起遗传、而甲乙两个隐性性状连系在一起的杂交组合。如:PL/pl。 相斥组:甲显性性状和乙隐性性状连系在一起遗传,而乙显性性状和甲隐性性状连系在一起的杂交组合。如:Pl/pL。 (二)连锁遗传的解释 试验结果是否受分离规律支配? 第一个试验: 紫花:红花(4831+390):(1338+393)=5221:1731 ≈3:1 长花粉:短花粉(4831+393):(1338+390)=5224:1728 ≈3:1 第二个试验: 紫花:红花(226+95):(97+1)=321:98 ≈3:1 长花粉:短花粉(226+97):(95+1)=323:96 ≈3:1 *以上结果都受分离规律支配,但不符合独立分配规律。 *F2不符合9:3:3:1,则说明F1产生的四种配子不等。 可用测交法加以验证,∵测交后代的表现型种类以及比例可反映出F1配子的种类和比例。 利用测交法验证连锁遗传现象: 特点:连锁遗传的表现为:
遗传学课后答案
第四章连锁遗传与性连锁 1.试述交换值、连锁强度与基因之间距离三者的关系。 答:交换值就是指同源染色体的非姐妹染色单体间有关基因的染色体片段发生交换的频率,或等于交换型配子占总配子数的百分率。交换值的幅度经常变动在0~50%之间。交换值越接近0%,说明连锁强度越大,两个连锁的非等位基因之间发生交换的孢母细胞数越少。当交换值越接近50%,连锁强度越小,两个连锁的非等位基因之间发生交换的孢母细胞数越多。由于交换值具有相对的稳定性,所以通常以这个数值表示两个基因在同一染色体上的相对距离,或称遗传距离。交换值越大,连锁基因间的距离越远;交换值越小,连锁基因间的距离越近。 2.在大麦中,带壳(N)对裸粒(n)、散穗(L)对密穗(l)为显性。今以带壳、散穗与裸粒、密穗的 纯种杂交,F1表现如何?让F1与双隐纯合体测交,其后代为: 带壳、散穗201株裸粒、散穗18株 带壳、密穗20株裸粒、密穗203株 试问,这2对基因就是否连锁?交换值就是多少?要使F2出现纯合的裸粒散穗20株,至少应中多少株? 答:F1表现为带壳散穗(NnLl)。 测交后代不符合1:1:1:1的分离比例,亲本组合数目多,而重组类型数目少, 所以这两对基因为不完全连锁。 交换值% =((18+20)/(201+18+20+203))×100%=8、6% F1的两种重组配子Nl与nL各为8、6% / 2=4、3%,亲本型配子NL与nl各为(1-8、6%) /2=45、7%; 在F2群体中出现纯合类型nnLL基因型的比例为: 4、3%×4、3%=18、49/10000, 因此,根据方程18、49/10000=20/X计算出,X=10817,故要使F2出现纯合的裸粒散穗20株,至少应种10817株。 3、在杂合体ABy/abY,a与b之间的交换值为6%,b与y之间的交换值为10%。在没有干 扰的条件下,这个杂合体自交,能产生几种类型的配子;在符合系数为0、26时,配子的比例如何? 答:这个杂合体自交,能产生ABy、abY、aBy、AbY、ABY、aby、Aby、aBY 8种类型的配子。 在符合系数为0、26时,其实际双交换值为:0、26×0、06×0、1×100=0、156%,故其配子的比例为:ABy42、078:abY42、078:aBy2、922:AbY2、922:ABY4、922:aby4、922:Aby0、078:aBY0、078。 3.设某植物的3个基因t、h、f依次位于同一染色体上,已知t-h相距14cM,现有如下杂 交:+++/thf×thf/thf。问:①符合系数为1时,后代基因型为thf/thf的比例就是多少?②符合系数为0时,后代基因型为thf/thf的比例就是多少? 答:①1/8 ②1/2 5、a、b、c 3个基因都位于同一染色体上,让其杂合体与纯隐性亲本测交,得到下列结果: +++ 74 ++c 382 +b+ 3 +bc 98 a++ 106
第八章染色体的结构变异
第八章染色体的结构变异 第一节染色体结构变异的根源 自然界中每一种生物的染色体数目和结构都是相对稳定的并且一般的情况之下,以整倍的方式复制自身,从而使每条染色体及上面负载的基因能稳定的传递,即保证了物种的稳定性。然而染色体结构的稳定是相对的变则是绝对的,在自然及人工诱变的情况下,染色体会发生与亲本相比较具有明显不同的缺失,重复,倒位,易位四种结构的变化,其中缺失,重复,倒位属于同源染色体之间的变化,易位属于非同源染色体之间的变化。那么这种变异的原因是什么呢? 一个完整无损的染色体不能同另外一个染色体结合的不受另外的染色体是完整的还是损伤的。实验又证明,只有新的此面才有重新粘合的能力,已经游离的染色体片断及颗粒一般是不再有粘合能力的。如果某一个染色体由于外因和内因的作用,断成二个或二个以上的新片段,其中必有一个片段有着丝点,染色体折断的这些片段可能按原来的直线方向和顺序再次的接合起来,这叫做重建。也可能再次接合时改变了原来的直线顺序或者同其中的染色体的断片接起来,这叫改组。由于染色体发生了断裂,断裂面连接时又发生不同方式的错接即发生了改组。如果按照改组了的染色体结构准确的复制,并在后代的细胞中得到保存,这样一个新型的染色体就完型了,从而原来的染色体的结构就改变了。所以改组是结构变异的根源,而断裂是结构变异的前奏,这在细胞学中被称为“先断后接”的假说,这是染色体结构变异的机理。
染色体的畸变在自然界中是广泛的存在的常常又是自然发生的,常因营养,温度,生理等方面的异常变化而引起染色体的断损,鸭跖单的培养液中因缺Ca,缺Mg就会导致染色体的断裂。洋葱的种子贮存的越久,幼苗的根端细胞内断损的染色体越多。如果人为的用各种理化因素处理,那和结构变异的频率就会大大增加。一个染色体无论发生怎样的断裂,必须具有一个着丝点者才能稳定。如果染色体具有双着丝点,在细胞分裂时,由于受到两个着丝点分别向两极的拉力,从而在两个着丝点之间断裂,造成再次的结构变异。无着丝点的片断由于在细胞分裂的后期得不到纺锤丝的牵引,被遗弃在新生的子核之外,最终丢失。 第二节缺失 一、缺失的定义及种类 1.定义:一个正常染色体的某一区段丢失,因而也随之丢失该区段上所负载的基因的这种变化,叫做染色体的缺失。 2.种类:缺失分为下面几种 A.顶端缺失:染色体缺失的区段可能在某臂的外端称为末端或顶端缺失。这种情况少见。如:a.bcde→a.bcd e B.中间缺失:染色体缺失的区段可能在某臂的内段,这种缺失多见。如:a.bcde→a.be cd 缺失之后会怎样呢? 染色体如果发生顶端的缺失就会在该臂上留有新的断头,断头 难以愈合,不能形成稳定的结构,因而可能发生下面两种可能:
染色体步移技术
螺旋讲堂2010年第4期总第23期 染色体步移 内容概览: 一、染色体步移技术概述 二、常见的染色体步移技术介绍 1、结合基因组文库的染色体步移技术 1) 物理剪切法构建亚克隆文库2) 限制性内切酶法构建亚克隆文库2、基于PCR 技术的染色体步移技术 1) 连接成环PCR 2)外源接头介导PCR (1)连接载体的PCR (2)连接单链接头的PCR (3)连接双链接头PCR 3)半随机引物PCR 策略 (1)新Alu-PCR (2)DW-ACP TM PCR (3)热不对称交错PCR (TAIL-PCR ) 三、染色体步移技术之TAIL-PCR 1、TAIL-PCR 的原理 2、TAIL-PCR 的优点以及技术难点 3、TAIL-PCR 的操作流程 4、TAIL-PCR 中的注意事项 5、常见问题分析 生物人的网上家园 plum 螺旋网HelixNet 染色体步移技术是一种常用的克隆已知片段旁侧序列的技术。本文中主要总结了近年来染色体步移技术的发展情况,介绍了结合基因组文库的染色体步移技术和 基于PCR 的染色体步移技术,并且比较了他们之间的优缺点,最后着重与大家一起 讨论一下TAIL-PCR 的相关内容,希望能对大家有所帮助。
一、染色体步移技术概述 染色体步移(Chromosome walking)又称为基因组步移(Genome walking)是指由生物基因组或基因组文库中的已知序列出发,逐步探知其旁邻的未知序列或与已知序列呈线性关系的目标序列的方法。对于已经完成基因组测序的模式生物种(如人、小鼠、线虫、水稻、拟南芥等),可直接从数据库中轻松的找到已知序列的侧翼序列。但是目前为止,自然界中大多数生物基因组DNA序列仍未知,要想知道一个已知区域两侧的DNA序列,染色体步移无疑是一种非常有效的方法。因此,染色体步移技术在现代分子生物学研究中较为重要,本文主要就染色体步移的相关方法,不同方法之间的比较,着重以TAIL-PCR的原理,实验操作及注意事项等进行讨论。 染色体步移技术的主要应用可归结为以下5个方面: (1)鉴定T—DNA或转座子的插入位点,鉴定转基因技术所导致的外源基因的插入位点; (2)根据基因的已知片段、EST或插入的转座子序列克隆目的基因,分离基因的启动子及调控元件; (3)用于人工染色体PAC、YAC和BAC的片段搭接; (4)构建图位克隆中的重叠群; (5)转化标记辅助育种中的STS或SCAR标记等。 二、常见的染色体步移技术介绍 目前,分离侧翼序列的染色体步移方法主要有两种,一是结合基因组文库为主要手段的染色体步移技术,构建基因组文库进行染色体步移尽管步骤比较繁琐,但是适于长距离步移,可以获得代表某一特定染色体的较长连续区段的重叠基因组克隆群。随着亚克隆文库条件构建条件的优化及测序技术的进步,这种方法也将更加快捷,准确。另一个是基于PCR扩增为主要手段的染色体步移技术。基于PCR扩增为主要手段的染色体步移技术步移距离相对较短,但是操作比较简单,尤其适合于已知一段核苷酸序列的情况下进行的染色体步移。在此基础上人们相继发明了十几种侧翼序列克隆的方法,依据其技术原理,可以将这些方法分为3类:连接成环PCR、外源接头介导PCR和半随机引物PCR。 1、结合基因组文库的染色体步移技术 基因组文库是指生物体全部DNA,经过合适的核酸内切酶消化或机械切割以后,克隆到适当的载体分子中,构成重组体分子群体,然后转化给诸如大肠杆菌这样的寄主菌株进行复制繁殖,如此构建的理论上含有生物体整个基因组全部遗传信息的克隆集合体,称作基因组文库,它是进行基因组学研究的重要技术平台,在基因组物理图谱的构建以及基因的图位克隆中发挥了重要作用。基因组文库的种类主要有入噬菌体文库、酵母人工染色体文库(yeast artificial chromosome,YAC)、P1噬菌体文库和细菌人工染色体文库(bacterial artificial chromosome,BAC)等。其中,BAC文库因其插入片段大、嵌合率低、遗传稳定性好、 易于操作等优点而备受青睐,近年来,小麦、棉花、水稻等越来越多的物种构建了BAC文库。 在基因克隆中,该方法用于鉴定一系列彼此重叠的DNA限制片段,分离序列及表达产物未知的基因,特别 是发育相关的基因。利用基因组文库克隆目的基因,首先要有一个根据目的基因建立起来的遗传分离群体, 找到与目的基因紧密连锁的分子标记,然后用遗传作图将目的基因定位在染色体的特定位置,找到与目的 基因紧密连锁的分子标记。构建含有插人大片段DNA的基因组文库(BAC或YAC),以与目的基因连锁的 分子标记为探针筛选基因组文库,鉴定出分子标记所在的大片段克隆,再以该克隆为染色体步移的起点, 用外侧克隆末端作为探针筛选基因组文库,分离新的重叠克隆用获得的阳性克隆,多次重复上述步骤,逐 渐逼近目的基因,构建目的基因区域的重叠群。随后通过亚克隆文库获得含有目的基因的小片段克隆,最 后通过遗传转化和功能互补验证最终确定目的基因的碱基序列。 目前构建亚克隆文库比较常用的DNA片段化方法主要有两种:物理剪切法和限制性内切酶法。