叶绿体形态、结构、分布及发生
叶绿体的基因组结构和进化

叶绿体的基因组结构和进化叶绿体是一种特殊的细胞器,其内部含有自己的基因组,可以独立进行基因表达和复制。
在细胞中,叶绿体的主要功能是进行光合作用,为细胞提供能量和有机物。
叶绿体的基因组结构和进化是生命科学领域中的一个热门研究方向,本文将从多个方面来探讨这个话题。
一、叶绿体基因组结构叶绿体的基因组一般分为两个部分:大环和小环。
大环是一个双链DNA分子,大小约为120kb,编码有多个重要的基因和调控序列。
小环是一个单链DNA分子,大小约为20kb,编码有一些核糖体RNA和一些参与蛋白合成的tRNA。
除此之外,叶绿体中还有一些裸人DNA,大小约为1-3kb,其中包含着一些无功能的序列。
总的来说,叶绿体中的基因数目较少,大约只有100来个,而且大多数基因都存在于大环中。
在叶绿体基因组中,编码有数个叶绿素类蛋白的基因,它们是光合作用中的关键部件。
此外,还有编码激酶、磷酸酯酶和转运蛋白等多个种类的基因。
叶绿体中有很多的反式重复序列(IRs),这些序列重复出现并分别位于大环的两端,起到了保护基因组完整性以及支持基因重排序的作用。
随着对叶绿体的研究的深入,人们逐渐发现,在叶绿体基因组中,还存在着不同程度的多样化。
这种多样化程度不只是在种类之间不同,也在同一个像的不同个体之间有所不同。
二、叶绿体基因组的来源叶绿体基因组的来源,一直是基因组学领域中的一项热点研究。
根据现有的研究成果,叶绿体基因组最可能的来源是一个单细胞的光合自养生物。
根据目前已知的信息,叶绿体最近的共同祖先应该出现在40亿年前左右。
它的基因组不断演化和分裂,最终形成了现在存在的繁多的叶绿体。
在叶绿体基因组的进化过程中,发生了很多的事件。
比如基因重排序,基因重复、剪接和互补等。
有些事件可能发生在叶绿体发生分裂的过程中,有些可能与外界环境的变化相关。
三、叶绿体基因组的重要性叶绿体基因组是很多植物学、生态学和生命科学的研究方向。
这是因为,叶绿体基因组可以用作分类鉴定标记或群体遗传学的研究对象,它可以被用来还原物种起源和进化历程,解析生物形态和生态适应性等问题。
观察叶绿体的形态和分布

一、实验原理:1、叶肉细胞中的叶绿体,散布于细胞质基质中,呈绿色,扁平的椭球形或球形。
可以在高倍显微镜下观察到它的形态和分布。
2、线粒体普遍存在于植物细胞和动物细胞中。
线粒体的形态多样,有短棒状、圆球形,线形,哑铃形。
3、键那绿(Janus green B)染液:是专一性,染线粒体的活细胞染液。
可以使活细胞中的线粒体呈蓝绿色。
而细胞质接近无色。
线粒体能在键那绿染液中维持活性数小时。
4、通过染色,可以在高倍显微镜下观察到生活状态的线粒体的形态和分布。
二、目的要求:1、初步掌握高倍显微镜的使用方法。
2、观察叶绿体的形态和分布。
3、观察线粒体的形态和分布。
三、材料用具:⑴材料:藓类的叶(或菠菜叶、黑藻叶等)。
四、试剂: ⑴新配制的质量分数为1%的键那绿染液(将0.5 g键那绿溶剂于5mL生理盐水中,加温到30~40℃使其充分溶解).五、用具:①显微镜②载波片③盖玻片④镊子⑤滴管⑥消毒牙签⑦铅笔四、方法步骤:(一)制作鲜类叶片临时装片:1、取材:⑴用镊子取一片藓类的小叶,把根部去掉。
⑵或者取菠菜叶稍带些叶肉的下表皮。
2、制片:⑴往载玻片中央滴一滴清水。
⑵用镊子夹住所取的以上小叶或下表皮,放入清水滴中。
⑶盖上盖玻片。
⑷注意:临时装片中的叶片不能放干了,要随时保持有水状态。
3、制作人口腔上皮细胞临时装片:⑴在洁净的载波片中央滴一滴键那绿染液。
⑵用消毒的牙签在自己漱静的口腔内壁上轻轻地刮几下,⑶把牙签上附有碎屑的异端,放在染液中涂几下,⑷盖上盖玻片。
(二)用显微镜观察叶绿体:1、用低倍显微镜观察:⑴对光:①转动显微镜的转换器,让一个高倍物镜对准载物台中央的通光孔。
②转动遮光器使一个大光圈对准通光孔。
③左眼向目镜里看,④同时右手转动反光镜,(光线弱时,用凹面反光镜。
光线强时,用平面反光镜。
)⑤当视野中看到一个明亮的光圈时,光就对好了。
⑵低倍显微镜观察:①将制好的叶的临时装片,放在载物台上。
让要看的物体在通光孔的正中央。
新课标高中生物人教版必修第一册第二册生物世界〖叶绿体和线粒体的结构与功能〗

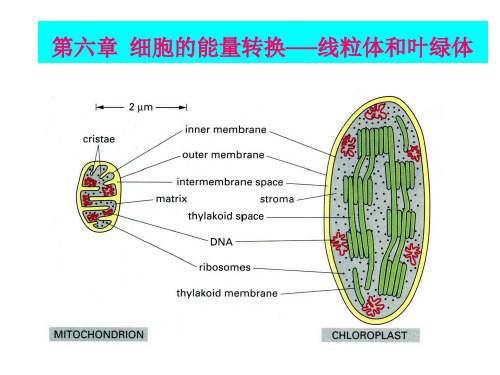
叶绿体和线粒体的结构与功能(1)叶绿体的结构与功能叶绿体是植物进行光合作用的细胞器。
具有叶绿体的植物除高等植物外,还有真核藻类。
叶绿体的形状因物种的不同而有所差异。
藻类的叶绿体形态差异较大,可以是板状、带状、杯状、囊状、星状等。
高等植物的叶绿体一般形状比较固定,多为扁平的椭球形,平均直径为4~6 μm,厚2~3 μm。
叶绿体由双层膜包被,每层膜厚6~8 nm,外膜与内膜之间有10~2021m宽的膜间隙。
两层膜均由单位膜(由脂质双层及嵌合蛋白质构成的一层生物膜)组成,具有选择透过性。
叶绿体膜内的基础物质称为基质。
基质中悬浮着复杂的膜片层系统,其基本单位是由单位膜封闭形成的扁平小囊,称为类囊体(也称片层)。
类囊体有规律地垛叠在一起形成好似一摞硬币的结构被称为基粒类囊体。
贯穿在两个或两个以上基粒之间没有发生垛叠的类囊体,称为基质类囊体。
相邻基粒由基质类囊体链接在一起,使类囊体腔之间彼此相通,因而,一个叶绿体内的全部类囊体实际上是一个连续的封闭的三维结构。
类囊体膜上有多种蛋白复合体,包括光合电子传递体和光合色素蛋白质复合体,是光合作用中进行光反应的结构。
类囊体膜上的光合色素负责在光合作用中吸收光能,这种膜片层系统极大地增加了光合作用中的受光面积,提升了光合作用的效率。
叶绿体的基质中有可溶性的蛋白(酶)以及其他活跃的代谢物质,其中包括光合作用中催化碳固定的酶系统,因此,光合作用中二氧化碳的固定、还原是在叶绿体基质中完成的。
基质中还存在叶绿体自身的DNA、RNA和核糖体,能够自主进行遗传物质的传递以及蛋白质的合成。
(2)线粒体的结构与功能与叶绿体相比,线粒体要小一些,直径05~1 μm,长1~2 μm,通常呈椭球状或圆柱体。
线粒体也由内外两层单位膜包裹,内外膜之间有腔。
外膜平整光滑,内膜内折形成嵴。
内膜上分布有许多规律排列的带柄的球状小体,即ATP合酶,它利用电子传递过程中形成的质子跨膜电化学势梯度合成ATP。
第六章 线粒体和叶绿体
1) 内膜结构
单层膜,比外膜厚; 通透性较外膜小; 向内折叠成嵴,表面积极大增加,代谢效率 极大提高;
嵴的形状与数量和细胞种类与生理状况有关
2) 线粒体分类(根据嵴的形态结构)
(a)板层形 (b)管形 (a),(b)为基本形 (c)羽冠形 (d)网膜形 (e)绒毛形 (f)平行形 (g)同心圆形
返回
第二节 叶绿体与光合作用
●一.叶绿体(Chloroplast)的形态结构
二.叶绿体的结构与化学组成
●三.叶绿体的功能—光合作用
(photosynthesis)
一、叶绿体(Chloroplast)的形态结构
●叶绿体与线粒体形态结构比较
叶绿体内膜并不向内折叠成嵴;内膜不含电
子传递链;除了膜间隙、基质外,还有类囊体; 捕光系统、电子传递链和ATP合成酶都位于类囊体 膜上。 ●叶绿体超微结构
1964年由Paul Boyer最先提出来的。此学
说认为电子沿电子传递链传递使线粒体 内膜蛋白质组分发生了构象变化,形成 一种高能形式。这种高能形式通过ATP 的合成而恢复其原来的构象。未能找到 有力的证据。
(3)化学渗透学说 1961 由英国生物化学家Peter Mitchell最先提出来
的。此学说认为电子传递释放出的自由能和ATP合成
+
个H 泵出内膜。
c 内膜对H+不能自由通过,泵出
膜外侧的H+不能自由返回膜内侧,
因而造成H+浓度的跨膜浓度梯度。
此梯度包含的能量可驱使ADP和Pi 生成ATP。
d.氢通过ATP合成酶上 的特殊的途径,返回基 质。H+浓度梯度所释放
的自由能,偶联ADP和
细胞生物学翟中和第四版-06-线粒体与叶绿体

(一)ATP合酶(ATP synthase)
ATP合酶分子结构模式图
ATP 合酶的分子由球形的头部和 基部组成,头部朝向线粒体基质, 规则性地排布在内膜下并以基部 与内膜相连。
ATP合酶的头部被称为偶联因子1 (F1),由5 种类型的9个亚基组 成,形成一个“橘瓣”状结构,其 中只有β 亚基具有催化ATP合成或 水解的活性。 F1 的功能是催化ATP合成,γ 亚 基的一个结构域构成穿过F1的中 央轴。ε 亚基协助γ 亚基附着到 ATP合酶的基部结构F0上。γ 与ε 亚基结合形成“转子”,旋转于 α 3β 3的中央,调节3个β 亚基催 化位点的开放和关闭。
第二节 叶绿体
• 光合作用是地球上有机体生存和发展的根本源泉。 • 绿色植物年产干物质达1014公斤。
一、叶绿体的基本形态及动态特征
(一)叶绿体的形态、分布及数目
• 在高等植物的叶肉细胞中,叶 绿体呈凸透镜或铁饼状,直径
为5~10 μm,厚2~4 μm。
• 分布在细胞质膜与液泡间薄层 的细胞质中,呈平层排列。 • 通常情况下,高等植物的叶肉 细胞含20~200 个叶绿体。
一、线粒体的融合与分裂
• 动、植物细胞中均可观察到频繁的线粒体融合与 分裂现象,这被认为是线粒体形态调控的基本方 式,也是线粒体数目调控的基础。 • 细胞中所有的线粒体构成一个不连续的动态整体。
线粒体融合与分裂的分子基础:
• 融合与分裂依赖于特定的基因和蛋白质的调控。融合所必需
的基因最早发现于果蝇,取名Fzo(fuzzy onion 模糊洋葱头) Fzo基因编码一跨膜的GTPase(鸟苷三磷酸酶),定位在线粒 体外膜上,介导线粒体的融合。
(一)叶绿体的形态、分布及数目
• 叶绿体通过位移避开强光 的行为称为躲避响应
第六章线粒体和叶绿体-细胞生物学

细胞色素氧 化酶复合体
> 13多肽链 c 2 血红素 O2 2 Cu
电子传递 路线
NADH Flavin Q 2 血红素、Ⅳ组成,催化琥珀酸的 脱氢氧化
氧化磷酸化的作用机理
• (一)质子动力势(P91)
能量以电化学质子浓度梯度 (Electrochemical Proton Gradient)的形式储存
电子传递过程中, H+ 被有关的酶复合体从线粒体基质中 排抽到膜间间隙 由此造成: 1. 在内膜两侧产生pH 梯度(pH gradient ,DpH ),基质 内的pH 高于膜间间隙(细胞溶胶)的pH • 在内膜两侧产生电压梯度( voltage gradient ,DV), 基质内为负电荷,基质外为正电荷 DpH 和 DV 共同形成电化学质子梯度 “Electrochemical Proton Gradient” 电化学质子梯度产生 “质子动力” (proton-motive force)
心磷脂(Cardiolipin)
3.膜间间隙 (Inter-membrane space): 是内外膜之间的腔隙,宽约6-8nm。 含有利用ATP来磷酸化其它核苷酸的酶 标志酶为腺苷酸激酶 4.基质 (Matrix), 含有: 线粒体 DNA (mt-DNA) 线粒体核糖体, 线粒体 tRNAs 数百种酶, 包括催化以下反应的酶系统: -丙酮酸和脂肪酸的氧化 (产生乙酰辅酶A [acetyl CoA] -柠檬酸循环 (产生 NADH) -线粒体基因的表达(转录、翻译等) 标志酶为苹果酸脱氢酶
第六章 细胞的能量转换—线粒体与叶绿体
主要内容:
第一节 线粒体与氧化磷酸化 第二节 叶绿体与光合作用 第三节 线粒体与叶绿体是半自主性细胞器及其起源
叶绿体的实验报告结果(3篇)

第1篇一、实验目的1. 通过高倍显微镜观察叶绿体的形态、分布及运动状态。
2. 分析叶绿体的生理功能及其与植物光合作用的关系。
二、实验原理叶绿体是植物细胞中进行光合作用的细胞器,其主要功能是吸收光能并将其转化为化学能。
叶绿体呈椭球形,含有叶绿素等色素,具有绿色外观。
在不同光照条件下,叶绿体会发生运动,以适应光能的吸收。
三、实验材料与仪器1. 实验材料:新鲜叶片(如菠菜、水稻等)2. 实验仪器:高倍显微镜、载玻片、盖玻片、滴管、镊子、刀片、培养皿、铅笔、显微镜载物台等四、实验方法1. 制作临时装片:取新鲜叶片,用刀片将叶片切成薄片,置于载玻片上,滴加适量的生理盐水,盖上盖玻片。
2. 观察叶绿体形态:在高倍显微镜下观察叶绿体的形态,记录叶绿体的形状、大小、颜色等特征。
3. 观察叶绿体分布:观察叶绿体在细胞内的分布情况,记录叶绿体的排列方式、密度等。
4. 观察叶绿体运动:在不同光照条件下,观察叶绿体的运动状态,记录叶绿体的运动方向、速度等。
5. 分析叶绿体的生理功能:结合实验现象,分析叶绿体的生理功能及其与植物光合作用的关系。
五、实验结果与分析1. 叶绿体形态:观察到的叶绿体呈椭球形,大小不一,颜色为绿色。
部分叶绿体内部可见明显的淀粉粒。
2. 叶绿体分布:叶绿体在细胞内呈散在分布,分布密度不均。
靠近细胞壁的叶绿体分布较为密集,而细胞核附近的叶绿体分布较为稀疏。
3. 叶绿体运动:在不同光照条件下,叶绿体表现出明显的运动。
在强光下,叶绿体以侧面朝向光源,运动速度较快;在弱光下,叶绿体以正面朝向光源,运动速度较慢。
4. 叶绿体的生理功能:叶绿体是植物进行光合作用的主要细胞器,其主要功能是吸收光能并将其转化为化学能。
叶绿体内含有叶绿素等色素,能够吸收光能。
在不同光照条件下,叶绿体的运动有助于提高光能的吸收效率,从而提高植物的光合作用效率。
六、结论1. 通过高倍显微镜观察,我们成功观察到了叶绿体的形态、分布及运动状态。
观察叶绿体结构实验报告(3篇)

第1篇一、实验目的1. 了解叶绿体的基本结构及其在植物细胞中的作用。
2. 通过显微镜观察叶绿体的形态和分布。
3. 掌握显微镜的使用方法,提高实验操作技能。
二、实验原理叶绿体是植物细胞中进行光合作用的细胞器,其主要功能是吸收光能,将水和二氧化碳转化为有机物质,并释放氧气。
叶绿体内部含有叶绿素,是进行光合作用的重要色素。
本实验通过显微镜观察叶绿体的形态和分布,分析其结构特点。
三、实验材料与仪器1. 实验材料:新鲜菠菜叶片2. 仪器:显微镜、载玻片、盖玻片、盖玻片夹、镊子、滴管、显微镜镜头、目镜、显微镜支架四、实验步骤1. 取新鲜菠菜叶片,用镊子撕取一片叶片,放入载玻片中。
2. 用滴管滴加适量的生理盐水,使叶片保持湿润。
3. 盖上盖玻片,轻轻压平,避免气泡产生。
4. 将载玻片放置在显微镜载物台上,调整镜头,观察叶绿体。
5. 仔细观察叶绿体的形态、大小、分布及与细胞质的关系。
6. 记录观察到的叶绿体结构特点。
五、实验结果与分析1. 叶绿体呈扁平椭球形,绿色,分布在细胞质中。
2. 叶绿体周围有较厚的细胞质,称为叶绿体基质。
3. 叶绿体内含有大量叶绿素,使其呈现绿色。
4. 叶绿体表面有少量的类囊体,是光合作用的主要场所。
5. 叶绿体在细胞中的分布不均匀,靠近细胞壁的叶绿体数量较多。
六、实验结论通过本次实验,我们观察到了叶绿体的基本结构,包括形态、大小、分布及与细胞质的关系。
叶绿体是植物细胞中进行光合作用的重要细胞器,其内部含有叶绿素,是进行光合作用的重要色素。
叶绿体的分布不均匀,靠近细胞壁的叶绿体数量较多,这可能与光合作用的强度有关。
七、实验讨论1. 叶绿体在植物细胞中的分布不均匀,这是否与光合作用的强度有关?2. 叶绿体在植物生长发育过程中,其形态和数量是否会发生改变?3. 叶绿体的形态和分布与光合作用的效率有何关系?八、实验总结本次实验通过观察叶绿体的结构,使我们了解了叶绿体的基本形态和分布,以及其在植物细胞中的作用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
.
四.叶绿体的超微结构
双层膜 类囊体 叶绿体基质
.
1. 叶绿体膜
外膜:厚6-8nm, 通透性大,含孔蛋白,相对分子质量达104的分子亦可通过
内膜:厚6-8nm, 通透性小,不含孔蛋白,仅允许O2、CO2、H2O分子自由 通过,但有许多转运蛋白选择性转运大分子进出叶绿体
膜间隙:内、外膜之间的腔隙
.
光照强度对叶绿体分布及位置影响的示意图
二.叶绿体的分化与去分化
.
二.叶绿体的分化与去分化
在种子萌发过程中,子叶、叶鞘和真叶细胞中的原质体相继分化叶绿体,这 种分化依赖于光照 分化表现:形态上,体积的增大,内膜系统的形成,叶绿素的累积
生化和分子生物学,叶绿体功能所必需的酶、蛋白质、大分子的 合成、运输及定位
时,这两种细胞器还具有自身遗传物质,编码一小部分必需的RNA和蛋白质
.
1.叶绿体的半自主性
叶绿体DNA呈环状,分子大小因物种不同存在较大差异,在200-2500kb之 间
叶绿体内含有4种RNA, 30中(烟草)或31种(地钱)或31(地钱)tRNA以 及100多种多肽
核质互作:在真核细胞中,细胞核与线粒体、叶绿体之间在遗传信息和基因 表达调控等层次上建立的分子协作机制
.
分化障碍形成白化苗
特定情况下,叶绿体的分化是可逆的,即可发生去分化 如:叶肉细胞经组织培养形成愈伤组织细胞时,叶绿体去分化再次形成原质体
.
.
叶绿体分化及分化异常的表现
三.叶绿体的分裂
.
三.叶绿体的分裂
部位:生长中的幼叶内 前叶绿体:幼叶中的叶绿体,体积为成熟叶绿体的1/10-1/5,基质内置形成
核质冲突:核质互作相关的细胞核或线粒体、叶绿体基因单方面发生突变, 引起细胞中的分子协作机制出现严重障碍时,细胞或真核生物个体通常会表
. 现出一些异常的表型,这种表型背后的分子机制即为核质冲突
.
DAPI对细胞进行染色
2. 叶绿体的起源
内共生起源学说认为(Mereschkowsky):叶绿体来源于行光能自养的蓝细菌与宝箱 相当 基因组与细菌基因组具有明显的相似性 具备独立,完整的蛋白质合成系统 分裂方式与细菌相似 与线粒体的膜一起与其他的膜存在明显不同
高等植物叶肉细胞含20-200个叶绿体 稳定性:高等植物叶肉细胞内叶绿体体积和数目的稳定 动态性:叶绿体定位(叶绿体在细胞内位置和分布受到的动态调控) 包括叶绿体的移动及移动后在新的最适位置上的“锚定” 躲避响应:叶绿体通过位移避开强光的行为 积聚响应:在光照较弱的情况下,叶绿体汇集到细胞的受光面的行为
叶绿体内膜与类囊体之间的液态胶体物质 主要成分:可溶性蛋白质和其他代谢活跃物质 如Rubisco,叶绿体DNA,核糖体, 核糖体,脂滴,植物铁蛋白和淀粉粒等
物质
.
.
五. 叶绿体的半自主性及其起源
半自主性 起源
.
1.叶绿体的半自主性
1962年,Ris和Plant在衣藻叶绿体中发现DNA状物质 叶绿体DNA(cpDNA)或称质体DNA(ptDNA) 半自主性:线粒体和叶绿体的功能依赖数以千记的和基因编码的蛋白质,同
少数基质类囊体,尚未或正在开始形成基粒类囊体。 分裂机制:分裂环(外环和内环)缢缩 叶绿体分裂装置:所有与叶绿体分裂相关的蛋白组成的分裂功能单位(未知)
.
.Leabharlann ARC5:与叶绿体分裂必需的 dynamin相关蛋白
PDV1/PDV2:外膜跨膜蛋白
.
高等植物叶绿体分裂过程中相关蛋白的定位
四.叶绿体的超微结构
.
2. 类囊体
定义:叶绿体内部由内膜衍生而来的封闭的扁平膜囊,主要成分是蛋白质和 脂类(60:40),不饱和脂肪酸含量高,膜流动性强
形成基粒与基质片层(或基质类囊体) 类囊体膜的主要蛋白有:光系统Ⅱ(PSⅡ),Cytb6f复合物,光系统Ⅰ
(PSⅠ)及CF0-CF1ATP合酶复合物
.
3. 叶绿体基质
动态连接(通过内外膜延伸形成管状突出——基质小管,实现叶绿 体间相互联系 基质小管的融合与分断有助于实现实时的物质或信息交换,还可能具备其他重要 生理功能
.
CHUP1: chloroplast unusual positional 1, 一种微丝结合蛋白, 编码该蛋白的基因突 变后叶绿体呈现定位 异常
叶绿体形态、结构、分布及发生
2013级非师1班2组 李红艳 程鸿敏 陈昆鹏 陈杨 董哲旭 贺蒋勇 金超凡 刘晨 罗茜 谭新苗 肖开元 袁莉 曾俊
岚 张雪
.
一.形态、分布及数目
.
光学显微镜下观察到的叶绿体
绿色 凸透镜或铁饼状 分布在细胞质膜与液泡间薄层的细胞质中,呈平层排列
.
一.形态、分布及数目