第5章受精的机制
发育生物学-复习资料-重点总结

绪论1、发育生物学:是应用现代生物学的技术研究生物发育机制的科学。
它主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老和死亡,即生物个体发育中生命现象发展的机制。
2、〔填空〕发育生物学模式动物:果蝇、线虫、非洲爪蟾、斑马鱼、鸡和小鼠。
第一篇发育生物学基本原理第一章细胞命运的决定1、细胞分化:从单个的全能细胞受精卵开始产生各种分化类型细胞的发育过程称细胞分化。
2、细胞定型可分为“特化”和“决定”两个阶段:当一个细胞或者组织放在中性环境如培养皿中培养可以自主分化时,可以说这个细胞或组织发育命运已经特化;当一个细胞或组织放在胚胎另一个部位培养可以自主分化时,可以说这个细胞或组织发育命运已经决定。
〔特化的发育命运是可逆的,决定的发育命运是不可逆的。
把已特化细胞或组织移植到胚胎不同部位,会分化成不同组织,把已决定细胞或组织移植到胚胎不同部位,只会分化成同一种组织。
〕3、〔简答〕胚胎细胞发育命运的定型主要有两种作用方式:第一种通过胞质隔离实现,第二种通过胚胎诱导实现。
〔1〕通过胞质隔离指定细胞发育命运是指卵裂时,受精卵内特定的细胞质别离到特定的裂球中,裂球中所含有的特定胞质可以决定它发育成哪一类细胞,而与邻近细胞没有关系。
细胞发育命运的这种定型方式称为“自主特化”,细胞发育命运完全由内部细胞质组分决定。
这种以细胞自主特化为特点的胚胎发育模式称为“镶嵌型发育”,因为整体胚胎好似是由能自我分化的各部分组合而成,也称自主型发育。
〔2〕通过胚胎诱导指定细胞发育命运是指胚胎发育过程中,相邻细胞或组织之间通过互相作用,决定其中一方或双方细胞的分化方向。
相互作用开始前,细胞可能具有不止一种分化潜能,但是和邻近细胞或组织的相互作用逐渐限制它们的发育命运,使之只能朝一定的方向分化。
细胞发育命运的这种定型方式成为“有条件特化”或“渐进特化”或“依赖型特化”,因为细胞发育命运取决于与其邻近的细胞或组织。
这种以细胞有条件特化为特点的胚胎发育模式称为“调整型发育”,也称有条件发育或依赖型发育。
动物繁殖学课件第五章受精、妊娠和分娩

3.发育所需营养物质主要来自母体输卵管和子宫
4.胚胎发育在透明带内进行
合子发育,以形态特征大体可分一下几个阶段:
1.桑椹胚 2.囊胚 3.原肠胚
第三节 妊娠识别与胚泡附植
一、妊娠的识别和建立
受精卵在输卵管中完成卵裂等部分早期胚胎发育阶段,便进 入子宫内并开始和子宫渐进性地建立联系。这种联系是以内分 泌活动为基础的。孕体(conceptus)是指胎儿、胎水、胎膜构 成的综合体。在妊娠初期,孕体产生的激素传给母体,母体对 此产生相应的反应,识别胎儿的存在,并在二者之间建立起密 切的联系,这一过程即为妊娠识别。 孕体和母体之间产生了信息传递和反应后,双方的联系和互 相作用已通过激素的媒介和其它生理因素而固定下来,从而胚胎 在子宫中生长发育,这叫做妊娠的建立(确立)。
2.卵子的运行
卵子本身并无运动能力,被伞部接纳的卵子,借助纤毛 颤动,沿着伞部的皱襞,通过漏斗口进入壶腹部。
3.卵子的运行速度
家畜卵子沿着输卵管运行的速度没有确切的材料。 卵子在输卵管内运行的时间包括: a.从伞部到达壶狭连接部的时间。 b.在壶狭连接部停留的时间。 c.通过壶狭连接部进入宫管连接部的时间。 卵子在输卵管运行的时间:牛90h,马98h,猪50h,羊72h。
(五)精子的运行速度
精子自射精部位到达受精部位的时间比精子自身运动 的时间要短。牛、羊在交配后15分钟左右,即可在壶腹 部发现精子。猪15~30分钟,马为24分钟左右。
(六)卵子的运行
1.卵子的接纳 母畜接近排卵时,输卵管伞充分开放、充血,并靠输卵 管系膜肌肉的活动使输卵管伞紧贴于卵巢表面,加之卵 巢固有韧带收缩而引起的围绕自身纵轴的旋转运动,使 伞的表面更接近卵巢的开口部。
2.精子在子宫中运行
发育生物学试题库

发育生物学试题库(发育生物学教学组)目录:第一章章节知识点与重点 (1)第二章发育生物学试题总汇 (6)第三章试题参考答案 (14)第一章章节知识点与重点绪论1.发育和发育生物学2.发育的功能3.发育生物学的基础4.动物发育的主要特点5.胚胎发育的类型(嵌合型、调整型)6.研究发育生物学的主要方法第一章细胞命运的决定1.细胞分化2.细胞定型及其时相(特化、决定)3.细胞定型的两种方式与其特点(自主特化、有条件特化)4.胚胎发育的两种方式与其特点(镶嵌型发育依赖型发育)5.形态决定子6.胞质定域(海胆、软体动物、线虫)7.形态决定子的性质8.细胞命运渐进特化的系列实验9.双梯度模型10.诱导11.胚胎诱导第二章细胞分化的分子机制1.细胞表型分类2.差异基因表达的源由3.了解基因表达各水平的一般调控机制第三章转录后的调控1.RNA加工水平调控2.翻译和翻译后水平调控第四章发育中的信号传导1.信号传导2.了解参与早期胚胎发育的细胞外信号传导途径第五章受精的机制1.受精2.受精的主要过程及相关知识3.向化性4.顶体反应5.皮质反应第六章卵裂1.卵裂特点(课堂作业)2.卵裂方式3.两栖类、哺乳类、鱼类、昆虫的卵裂过程及特点4.(果蝇)卵裂的调控机制第七章原肠作用1.了解原肠作用的方式:2.海胆、文昌鱼、鱼类、两栖类、鸟类、哺乳类的原肠作用基本过程与特点第八章神经胚和三胚层分化1.三个胚层的发育命运第九章胚胎细胞相互作用-诱导1.胚胎诱导和自动神经化、自动中胚层化2.胚胎诱导、异源诱导者3.初级诱导和次级诱导、三(多)级诱导4.邻近组织相互作用的两种类型5.间质与上皮(腺上皮)的相互作用及机制第十章胚轴形成1.体形模式2.图式形成3.果蝇形体模式建立过程中沿前后轴不同层次基因的表达4.果蝇前后轴建立的分子机制5.果蝇背腹轴形成的分子机制第十一章脊椎动物胚轴的形成1.什么是胚轴2.两栖类胚轴形成过程及分子机制3.了解鸟类、鱼类、哺乳类动物胚轴形成过程及分子机制第十二章脊椎动物中枢神经系统和体节形成机制1.脊椎动物中枢神经系统的前后轴形成2.脊椎动物中枢神经系统的背腹轴形成3.脊椎动物体节分化特征第十三章神经系统的发育1.神经系统的组织发生神经系统的组成来源(神经管、神经嵴、外胚层板)中枢神经系统的组织发生(脊髓、大脑、小脑、核团)神经系统发生过程中的组织与调控(位置、数目)2.神经系统的功能建立3.神经突起(树突和轴突)4.局部有序投射5.突触第十四章附肢的发育和再生1.附肢的起源2.附肢的早期发育附肢发育中外胚层与中胚层的相互作用附肢发育中轴性建立3.附肢再生(再生过程、再生调节)第十五章眼的发育1.视泡发育、分化2.晶状体发育、分化3.晶状体再生4.角膜发育第十六章变态1.变态2.昆虫变态的激素调控3.两栖类变态的激素调控第十七章性腺发育和性别决定1.哺乳动物的性腺发育2.哺乳动物的性别决定3.果蝇的性别决定4.雌雄同体、环境性别决定第十八章生殖细胞的发生1.精子发生:特点,过程2.卵子发生:特点,过程第十九章干细胞1.干细胞2.干细胞分类3.了解干细胞的应用第二十章动物发育的环境调控1.发育与环境关系2.环境对正常发育的调控3.环境对正常发育的干扰4.遗传与环境之间的相互作用第二章发育生物学试题样题总汇一、填空题(每空1分)1.发育生物学研究的主要内容是和,其主要任务是研究生命体发育的及其机制。
发育生物学1—7章 课后习题答案

《发育生物学》课后习题答案绪论1、发育生物学的定义,研究对象和研究任务?答:定义:是应用现代生物学的技术研究生物发育机制的科学。
研究对象:主要研究多细胞生物体从生殖细胞的发生、受精、胚胎发育、生长到衰老死亡,即生物个体发育中生命现象发展的机制。
同时还研究生物种群系统发生的机制。
2、多细胞个体发育的两大功能?答:1.产生细胞多样性并使各种细胞在本世代有机体中有严格的时空特异性;2.保证世代交替和生命的连续。
3、书中所讲爪蟾个体发育中的一系列概念?答:受精:精子和卵子融合的过程称为受精。
卵裂:受精后受精卵立即开始一系列迅速的有丝分裂,分裂成许多小细胞即分裂球,这个过程称为卵裂。
囊胚:卵裂后期,由分裂球聚集构成的圆球形囊泡状胚胎称为囊胚。
图式形成:胚胎细胞形成不同组织,器官和构成有序空间结构的过程胚轴:指从胚胎前端到后端之间的前后轴和背侧到腹侧之间的背腹轴4、模式生物的共性特征?答:a.其生理特征能够代表生物界的某一大类群;b.容易获得并易于在实验室内饲养繁殖;c.容易进行试验操作,特别是遗传学分析。
5、所讲每种发育生物学模式生物的特点,优势及其应用?答:a.两粞类——非洲爪蟾取卵方便,可常年取卵,卵母细胞体积大、数量多,易于显微操作。
应用:最早使用的模式生物,卵子和胚胎对早期发育生物学的发展有举足轻重的作用。
b.鱼类——斑马鱼受精卵较大,发育前期无色素表达,性成熟周期短、遗传背景清楚。
优势:a,世代周期短;b,胚胎透明,易于观察。
应用:大规模遗传突变筛选。
c.鸟类——鸡胚胎发育过程与哺乳动物更加接近,且鸡胚在体外发育相对于哺乳动物更容易进行试验研究。
应用:研究肢、体节等器官发育机制。
d.哺乳动物——小鼠特点及优势:繁殖快、饲养管理费用低,胚胎发育过程与人接近,遗传学背景较清楚。
应用:作为很多人类疾病的动物模型。
e.无脊椎动物果蝇:繁殖迅速,染色体巨大且易于进行基因定位。
酵母:单细胞动物,容易控制其生长,能方便的控制单倍体和二倍体间的相互转换,与哺乳动物编码蛋白的基因有高度同源性。
第五章 受精生物学大全

4. 受精发生 在第二次减 数分裂完成 后,例如: 海胆和腔肠 动物。
4
5 不同动物受精发生于卵子发育的不同阶段(引自Longo FJ,1997 )
二、受精方式
(一)体内受精和体外授精
1. 体外受精:排在水里 水生动物,如许多鱼类和部分两栖类等 2. 体内受精:进入生殖道抵达受精部位 高等动物如爬行类、鸟类、哺乳类、某些软体 动物、昆虫以及某些鱼类和少数两栖类
五、精子在获能中产生的变化
3. 超激活运动
获能过程中,精子运动能力逐渐加强,运动 方式发生改变。 • 许多哺乳动物的精子在发生顶体反应之前 运动能力加快,称为超激活
(superactivation)。
31
超激活现象
• 当金黄地鼠精子开始接触获能液时,开始时呈 活跃的线性向前运动,随后进入相对缓和的运 动状态,此时大多数精子以头对头相聚集,尾 部颤动。2h后则又互相离开,开始前所未有的 活跃运动,起初线性向前,很快出现猛烈的特 殊运动,尾部呈现强烈的鞭打样摆动,头的运 动轨迹呈不规则8字形。 • 小鼠等多种啮齿类动物和金黄地鼠类似,但许 多哺乳动物和人精子获能时不发生凝集。
2.5亿?
部位 子宫 子宫 阴道 子宫 阴道和子宫 阴道 阴道 子宫 阴道
在壶腹部精子数 (个) 100 500 250-500 18-1600 25-50 少量 600-700 1000 200
20个?
10
各种动物精子在输卵管中维持活动能力和受精能力的时间
动物 活动能力 受精能力(h) 动物 (h) 小鼠 13 6 羊 活动能力 受精能力(h) (h) 48 24-48
21
二、获能条件
1. 体内获能
• 没有严格的器官特异性,可以在妇女腹腔中进 行。 • 没有种属特异性,因为一种动物精子能够在其 他种类动物的雌性生殖道内获能。 • 雌性生殖道中输卵管特异糖蛋白(OGP)对精 子获能起直接作用 • 多种激素和因子对获能有促进作用:PAF、孕 酮、肝素和载脂蛋白
第五章-受精

第五章受精受精是单倍体的精子和卵子相互结合和融合而形成双倍体合子的过程,是有性生殖生物个体发育的起点。
它一方面保证了双亲的遗传作用,另一方面恢复了染色体双倍体数目。
同时受精可以把个体发生过程中产生的变异通过生殖细胞遗传下去,保证了物种的多样性,在生物进化上具有重要意义。
第一节受精研究历史的简单回顾1875年和1876年,Hertwig和Fol分别在各自独立的研究中首先在海胆中发现了受精现象。
Hertwig发现了海胆精子入卵后雌雄两原核融合的现象,而Fol发现了精子接近和穿入卵子以及受精膜形成的过程,至此结束了胚胎学上的“精源学说”和“卵源学说”。
1883年,van Beneden发表了马副蛔虫受精的细胞生物学研究论文,肯定了在遗传上父母贡献均等的理论,并使精卵作用的研究更为深入。
他在马副蛔虫受精卵的第一次有丝分裂纺锤体上看到四条染色体,其中两条来自父方,两条来自母方,提出父母的染色体通过精卵的融合传给子代。
此后,H.T. Boveri在马副蛔虫的研究工作进一步巩固了上述理论,把染色体看作是遗传信息的载体。
二十世纪以来,有关受精的研究转向探讨受精后生理学变化和两性配子结合的机制。
在早期的研究中发现受精后细胞膜通透性变化、耗氧量增加等现象。
1912年以后,F.R. Lillie根据在沙蚕和海胆获得的一系列研究结果,提出了精卵相互作用的受精素假说。
根据这一假说,卵子分泌称为受精素(fertilin)的物质,使精子运动能力加强,并向卵子聚集。
四十年代前后,A. Tyler 就受精素的生物学、化学和免疫学特征展开了一系列工作,并进一步强调卵子在成熟过程中分泌的物质对受精有重要意义。
与此同时,M. Hartmann在海胆的研究工作中证实,不仅卵子能分泌受精素(雌配素),精子也能排出抗受精素(雄配素),二者相互作用的程度决定着受精的成功与否。
不久以后,在两栖类上发现卵外胶膜在精卵相互作用中发挥重要功能,为两栖类受精所必需。
05第五章妊娠Pregnancy
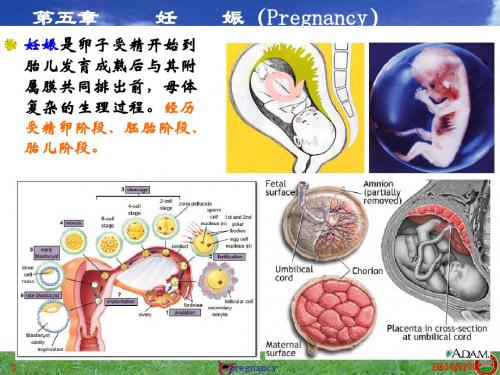
Pregnant :RPL increases after day 7, h P4 ;PGF increases day 11;rodent placental lactogen protects CL from PGF
2019/7/18
pregnancy
17
妊娠信号:
猪:雌激素(11-13d); 绵羊:滋养层素(15d)(oTP-1/IFN-τ); 牛:bTP-1、早孕因子(EPF,early pregnancy factor, 保护胎儿不受母体免疫系统的损害,小鼠在交配后 4~6h就出现在外周血液中); 灵长类:hCG(7-10d)。
Estrous cycle - 4 days
Mating induces prolactin release :Maintenance of CL ; Psuedopregnancy
Not Pregnant :Prolactin decreases after day 7 ;PGF increases day 11 ;Estrus by day 17
2019/7/18
pregnancy
12
猪的妊娠识别
猪的胚泡能产生雌激素,使母体产生妊娠识别。
猪的妊娠识别机制是,受精后的囊胚或胚泡于11d~ 12d开始迅速生长,大约在12d~30d期间,滋养层的外 胚层利用母体中的前体合成雌二醇和雌酮。然后在子 宫内结合成为硫酸雌酮。
雌激素发生局部作用,使子宫内膜合成PGF2α减少,同 时也阻止分泌至子宫腔内的PGF2α释入子宫静脉,以致 不能进入全身血循环和卵巢,黄体就不会受到影响而
马属动物的妊娠识别机制比灵长类动物更加复杂,一直到 怀孕35d天左右才能检测到CG,说明马属动物的孕体能改 变子宫静脉中PGE2/PGF2α的比例,在怀孕35d之前由PGE2 发挥促黄体化功能。
第五章授精与妊娠

精子通过子宫颈示意图
子宫颈粘液扫描电镜像片 左:排卵时子宫颈粘液扫描电镜像片;右:黄体期子宫颈粘液扫描电镜像片 注意:两图子宫颈粘液“网格”的相对大小,左图中精子的相对大小
精子通过子 宫颈进入子 宫示意图
上:精子通 过子宫颈进 入子宫诸因 素
下:精子与 子宫颈上皮 细胞的相对 大小
精子在子宫颈中运输扫描电镜像片 左:精子在子宫颈上皮细胞上的排列图 右:精子与子宫颈上皮细胞的相对大小
囊胚初形成时,外面仍Байду номын сангаас透明带,进 入子宫角后,囊胚腔内液体增多,使 它很快增大,并突破透明带。此后, 囊胚就变成透明的泡状称为胚泡。
(三)原肠配的发育
胚泡的进一步发育,出现内胚层,此时称 为原肠期。随着胚泡的发育,胚结上面的 滋养层细胞溶解退化,胚结裸露出来,称 为胚盘,随后,胚盘的向着囊胚腔的部分 以分层的方式逐渐形成一个新的细胞层, 向胚泡的内壁扩展,成为完整的一层--内胚 层。其外面的细胞层成为外胚层。以后在 内胚层和滋养层之间出现中胚层。
•精子顶体反应示意图 A:精子顶体反应前;B:精子顶体反应 中 C:精子顶体反应后
精子顶体反应示意图 1、精子顶体反应前
2、精子顶体反应后 注意精子与卵黄膜的 结合位点 3、精子顶体反应
a. 精子顶体反应前 b. 精子顶体反应中 c. 精子顶体反应后期, 注意脱落的精子膜和 顶体外膜 d. 精子顶体反应后
三、受精
1、顶体反应(AR,acrosome reaction)出现 顶体囊泡化 包裹精子顶体膜破裂,释放出内容物的过程。这 是精卵融合必须经历的反应之一。也是获能后的 必然结果。有人称顶体的变化为精子结构的获能 。顶体反应与物种及周围环境有关。
2、受精过程 受精始于精、卵相遇,两性原核合并形成合子时 结束。 受精是指精卵完成生理成熟后,两者互相相遇, 精子主动向卵子内部转入,从而出现一系列的复 杂的细胞学和细胞化学的变化过程,此变化才能 使两者充分结合成新的接合子。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第五章受精的机制动物受精研究内容:精子获能和顶体反应、卵子的激活精卵融合,受精后生化变化动物受精方式受精:是指两性生殖细胞融合并形成具备双亲遗传潜能的新个体的过程。
受精作用是发育的开端,一个新的生命从此开始。
受精的功能1、将父母的基因传递给子代2、激发卵细胞中确保发育正常进展的一系列反应。
受精过程中几个重要的事件:精子与卵子的接触与识别、精子入卵、精子与卵子遗传物质的融合以及卵子的激活和发育开始。
受精既可以发生在体外,也可以在体内发生。
1. 成熟精子和卵子的结构2. 精子和卵子的接触和识别3. 精子入卵的调控4. 卵子代谢的激活5. 遗传物质的融合一、成熟精子和卵子的结精子头部由顶体囊泡(acrosomal vesicle)和精核构成。
顶体位于精核前端,由高尔基体演化而来。
顶体中含有多种水解酶,主要作用是溶解卵子的外膜。
有些动物的顶体中还有与精卵识别有关的分子。
整个精核是一致密结构,几乎看不到染色质丝和核仁,精子中所有的基因都不表达。
不同的动物以不同的形式推动精子游动。
大多数动物的精子依靠尾部或鞭毛波浪式的摆动,推动精子在水或雌性动物生殖道中移动。
鞭毛具有特殊的结构,轴纤丝(axoneme)是鞭毛中起推动作用的主要部分,具有典型的“9+2”结构。
哺乳动物精子鞭毛的横切面,示中央的轴纤丝(central axoneme)和外围的纤丝( external fibers)。
精子的运动装置——鞭毛的超微结构,示微管排列的“9+2”结构。
卵质外是质膜(plasma membrane),质膜外是卵黄膜(vitelline envelope)。
质膜在受精时可以调控特定的离子在卵子内外的流动,且能与精子质膜融合。
卵黄膜能识别同一物种的精子,对受精的物种特异性有非常重要的作用。
在哺乳动物中特称为透明带(zona pellucida),紧靠着透明带的一层滤泡细胞称为放射冠(corona radiata)。
皮层(cortex)是质膜下一层约5um厚的胶状胞质,比内部的胞质硬,含有高浓度的肌动蛋白分子,受精时,聚合成微丝,延伸到细胞表面形成微绒毛(microvilli),帮助精子进入卵子。
皮层内有皮层颗粒(cortical granule),含消化酶、粘多糖、黏性糖蛋白和透明蛋白,阻止多精入卵并可以为卵裂球提供支持。
精子和卵子的接触和识别受精过程中,精卵的识别具有种属特异性,即给定物种的卵子对同源精子的识别与结合具有绝对的特异性。
造成这种特异性的原因在于雌雄配子表面具有某些结构互补的特异分子,通过这些特异分子之间的相互作用,保证了雌雄配子的正确识别。
精子和卵子的相互作用主要分为5个步骤:1. 精子的趋化性( chemotaxis )2. 精子的顶体反应,释放水解酶。
3. 精子与卵子外围的卵黄膜(透明带)结合4. 精子穿过卵外的结构5. 精卵细胞质膜的融合1、精子的趋化性精子的趋化性(chemotaxis)是指精子根据化学浓度梯度直接向卵子运动的现象。
现已在许多动物中发现,其卵母细胞完成第二次减数分裂后,可以分泌具有物种特异性的的趋化因子如海胆的精子激活肽resact,构成卵周特有的微环境。
2、精子获能哺乳动物的精子需要在雌性生殖道中停留一个特定的时期,以获得对卵子受精的能力,这一过程称为精子获能( capacitation )。
获能期间,精子的细胞膜发生了一系列变化,包括内膜分子重排、精子表面某些成分移除,但分子机制还不很清楚。
哺乳动物精子的获能,牛精子在到达输卵管壶腹部之前与输卵管上皮细胞的接触。
3. 顶体反应顶体反应是指受精前精子在同卵子接触时,精子顶体产生的一系列变化。
具有顶体结构的无脊椎动物或脊椎动物中,只有发生顶体反应的精子才能进入卵子并与卵子融合,也只有精子与卵子接触时才发生顶体反应。
顶体反应是受精的先决条件顶体中含有顶体酶系统,包括:透明质酸酶,放射冠分散酶和顶体素。
顶体内的结合素可识别特异的糖基序列,以保证精子与卵的种特异性结合。
诱发顶体反应的条件?:卵丘细胞,透明带,离子:Ca 2+,Na+,k+海胆的顶体反应精子与卵子胶膜结合后,可引起顶体反应。
顶体反应包括两个主要的事件:顶体膜与精子质膜发生融合以及顶体突起(acrosomal process)的形成海胆精子的顶体反应过程哺乳动物的顶体反应哺乳动物的顶体是一个帽状结构,覆盖于精核的前端。
顶体反应时,顶体帽部分的质膜与顶体外膜在多处发生融合,使顶体内的物质从融合处释放出来。
精子顶体酶及其穿透作用通过顶体反应释放的物质中含有大量的水解酶,因此顶体这一结构被认为具有类似溶酶体的功能。
精子顶体酶的一个功能是在卵子外围打一个洞,另外顶体反应还与精卵结合有关。
4,精卵识别的特异性海胆的精卵识别海胆精子发生顶体反应后释放顶体酶,使卵细胞外的胶膜降解,精子穿越胶膜,其突起与卵黄膜相互识别,与之融合,然后与卵细胞膜融合,导致精核进入卵细胞中。
海胆精子顶体突起与卵子微绒毛的接触海胆的精卵识别是由特异性结合蛋白(Bindin)所介导的。
Bindin定位在精子的顶体突起上,具有种属特异性。
卵子卵黄膜上存在Bindin的受体,也被分离纯化出来。
海胆精子顶体突起上Bindin的定位哺乳动物的精卵识别哺乳动物精卵的特异性识别发生在卵细胞的透明带(zona pellucida)部分。
小鼠透明带中含有ZP3 糖蛋白,它与ZP1、ZP2以网状的骨架结构存在于透明带中。
ZP3能结合精子,并引发顶体反应。
精子细胞膜上有三种受体:1. sp56(56kDa,半乳糖结合蛋白)---可与ZP3分子上的半乳糖端部相结合。
如果ZP3的一个半乳糖基发生丢失或改变,精子将无法与卵子结合。
2. 半乳糖基转移酶 (GalTase)—可与ZP3分子上的N-乙酰葡糖胺结合,使精子G蛋白激活并诱导顶体反应。
3. P95 (ZP受体激酶)—(95kDa)一种跨膜蛋白,其外侧部分可与ZP3分子特异结合,而内侧部分具有酪氨酸激酶的功能。
该酶被激活后,导致顶体反应。
二次结合在顶体反应期间,与ZP3结合的顶体前端发生胞吐作用,精子必须与透明带结合才可以完成穿透作用,这种结合称为二次结合(secondary binding)。
二次结合是通过顶体内膜上的特殊蛋白与透明带中的ZP2糖蛋白结合的。
三、精子入卵的调控1、雌雄配子的融合精子通过与卵黄膜或透明带的相互作用,发生顶体反应,使和精子结合的卵黄膜或透明带被顶体反应释放的水解酶溶解,并在该位置进行精卵细胞膜的融合。
雌雄配子的融合大多被限定在特定的区域内。
2、多精受精阻断的机制与皮层反应许多精子都可以到达卵子的表面并与之吸附,但是通常只有一个精子能完成受精,称为单精入卵。
多个精子入卵受精称为多精入卵,将导致死亡或不正常的发育。
生命发展了多种机制防止多倍的染色体组融合,最普通的办法是阻碍多精入卵。
海胆阻碍多精入卵的作用方式海胆卵子细胞膜去极化引起的快速的阻碍作用;卵子皮层颗粒的胞吐作用产生的一种较慢的阻碍作用;卵子细胞质降解额外精子的核酸或排出包含有额外精子核酸的细胞质。
(1)快速阻碍多精入卵卵膜中存在离子通道,卵膜的快速阻碍多精入卵作用是通过改变自身膜电位形成的。
精子进入卵细胞触发细胞膜静息电位迅速去极化,引起膜外精子与卵细胞识别和融合的障碍。
如人为维持原有的膜电位,可诱导多精受精现象发生;如改变正常的初始膜电位,则会阻止卵细胞的受精。
(2)皮层颗粒反应多精受精快速阻碍机制中膜电位的变化时间非常短暂(1 min左右),不足以永久实现阻碍多精入卵。
结合到卵黄膜的精子是通过皮层的小泡破裂,发生皮层反应被移除的;否则将导致多精入卵。
皮层由皮层颗粒组成,通常分布在没有微绒毛结构的卵膜下方。
当精子进入卵子时,皮层颗粒与卵膜发生融合,颗粒内容物被释放到卵膜和卵黄膜之间的区域——卵黄周隙(perivitelline space),这些释放物中有几种蛋白质在皮层颗粒反应中发挥了重要的作用。
皮质颗粒内含物中含有的蛋白质:1). 蛋白水解酶可以使卵黄膜与质膜间的联系分离;剪除卵膜上bindin的受体及与之结合的精子。
2). 粘多糖:进入卵周隙吸水膨胀,使卵黄层向外隆起,形成受精膜(fertilization envelope )举起。
3). 过氧化物酶:皮层颗粒分泌的过氧化物酶通过交联相邻蛋白质的酪氨酸残基使受精膜变硬。
受精膜最先在精子入卵的位置形成,并向外扩张至整个卵细胞,从而阻止多精入卵。
4). 透明质素(hyalin):在卵外形成透明质层(hyalin layer),它与卵裂中对分裂球的支持作用有关。
卵细胞质膜突起的微绒毛深入到其基部。
哺乳动物不形成受精膜,但皮质颗粒中释放的酶对透明带中的精子受体分子进行修饰,使之丧失与精子结合的能力,因此,称为透明带反应。
半乳糖基转移酶 (GalTase)—可与ZP3分子上的N-乙酰葡糖胺结合,使精子G蛋白激活并诱导顶体反应。
卵激活时皮质颗粒释放出来的N-乙酰葡糖酶能对ZP3上的GalTase结合位点进行修饰,由此阻断透明带外围的精子与受精卵结合。
)3、钙的作用皮层颗粒反应的作用机制与顶体反应基本相似,Ca2+作为细胞内信使发挥了及其重要的作用。
研究证明,启动皮层颗粒反应的钙主要储存在卵细胞的内质网中,而不是卵外钙离子的内流。
卵细胞内的信号分子三磷酸肌醇IP3激活Ca+的释放,而IP3的产生是与GTP结合蛋白或酪氨酸激酶相关的。
钙离子的作用:传导神经信号:促进神经递质分泌让心脏跳动:使细胞内外发生电位差传递御敌信号:启动了钙离子介导的信号通路,促使免疫细胞分化和生长调节酶的活性:钙调节蛋白与钙离子形成复合物调控生殖细胞的成熟和受精细胞内质网正常释放钙离子的机制:内质网释放钙离子后,内部的负电荷会增加,从而阻止钙离子继续释放,随后外部带正电荷的物质会进入内质网,并中和负电荷,使钙离子的释放保持正常内质网的膜上有“TRIC”的通道。
带正电荷的钾离子可以经由这种通道进入内质网,从而保证内质网能够正常释放钙离子。
四、卵子的激活机制未受精的卵子是惰性的,细胞的呼吸活动、RNA的转录和蛋白质的合成处于或几乎处于零水平。
只有受精的刺激才能唤醒其代谢的活跃进行。
这一活化过程分为两个阶段:一、早期反应(应答):指从精卵接触到发生皮质反应的数秒钟内所发生的事件。
二、晚期反应(应答):在受精开始后数分钟内发生的事件。
五、遗传物质的融合1. 雌雄原核的形成及融合单倍体的精子核进入卵细胞后,解凝聚形成雄原核,卵细胞核则形成雌原核,精核形成雄性原核的过程受到卵子的严格控制。
精子入卵时线粒体与鞭毛在卵细胞内降解,因此线粒体基因一般认为是母源性的;但中心体是父源性的。
海胆雌雄原核的融合海胆雄原核形成后旋转180o,其中心体位于雌、雄原核之间,装配成星体,连接并牵动雄原核与雌原核相互靠近,最后融合形成合子核(zygote nucleus)哺乳动物雌雄原核的融合哺乳动物精子入卵后,在卵母细胞胞质中谷胱甘肽的作用下精子染色体解凝聚。