62第六讲_Lecture_2_真核细胞的转录过程-讲义R
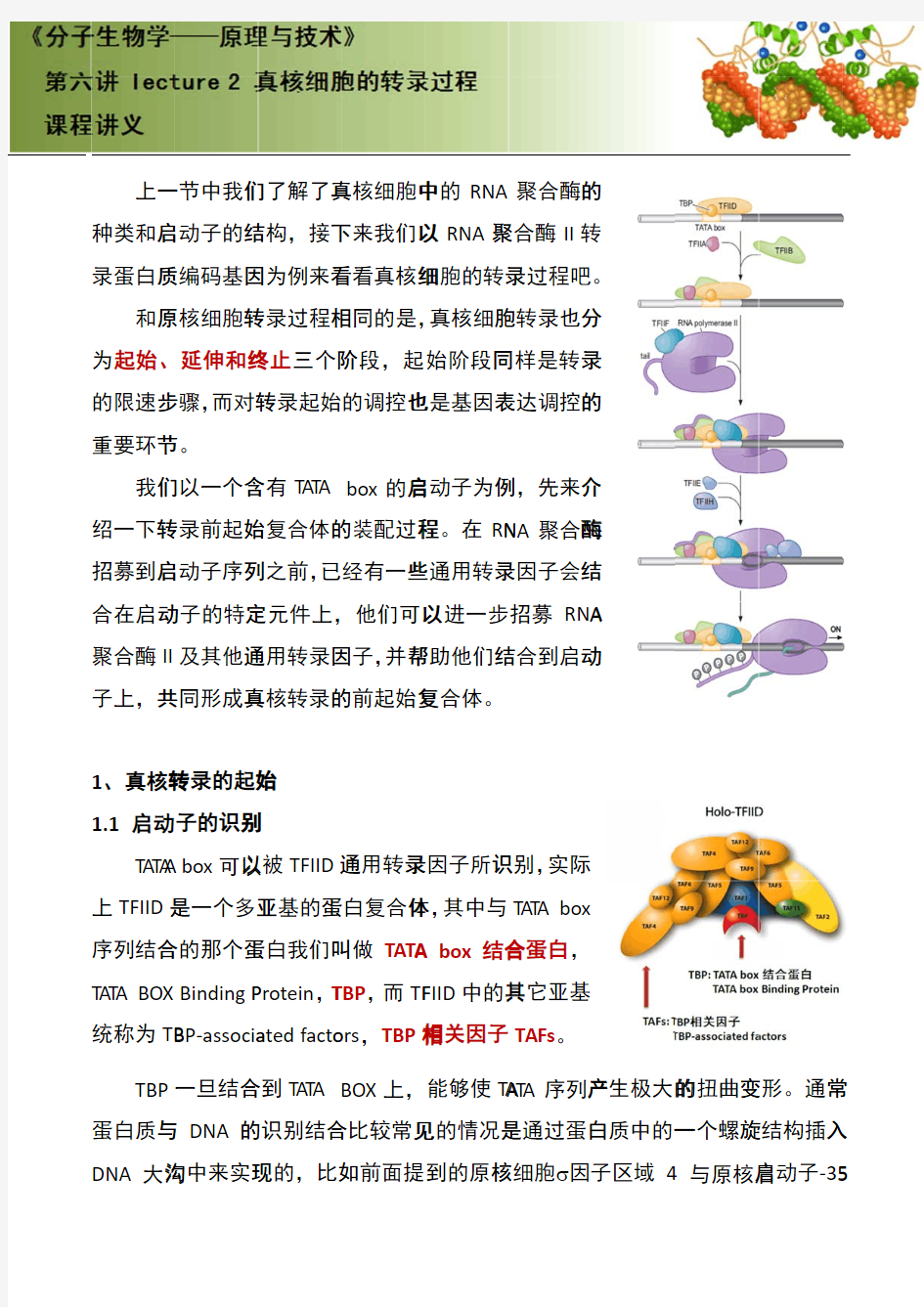
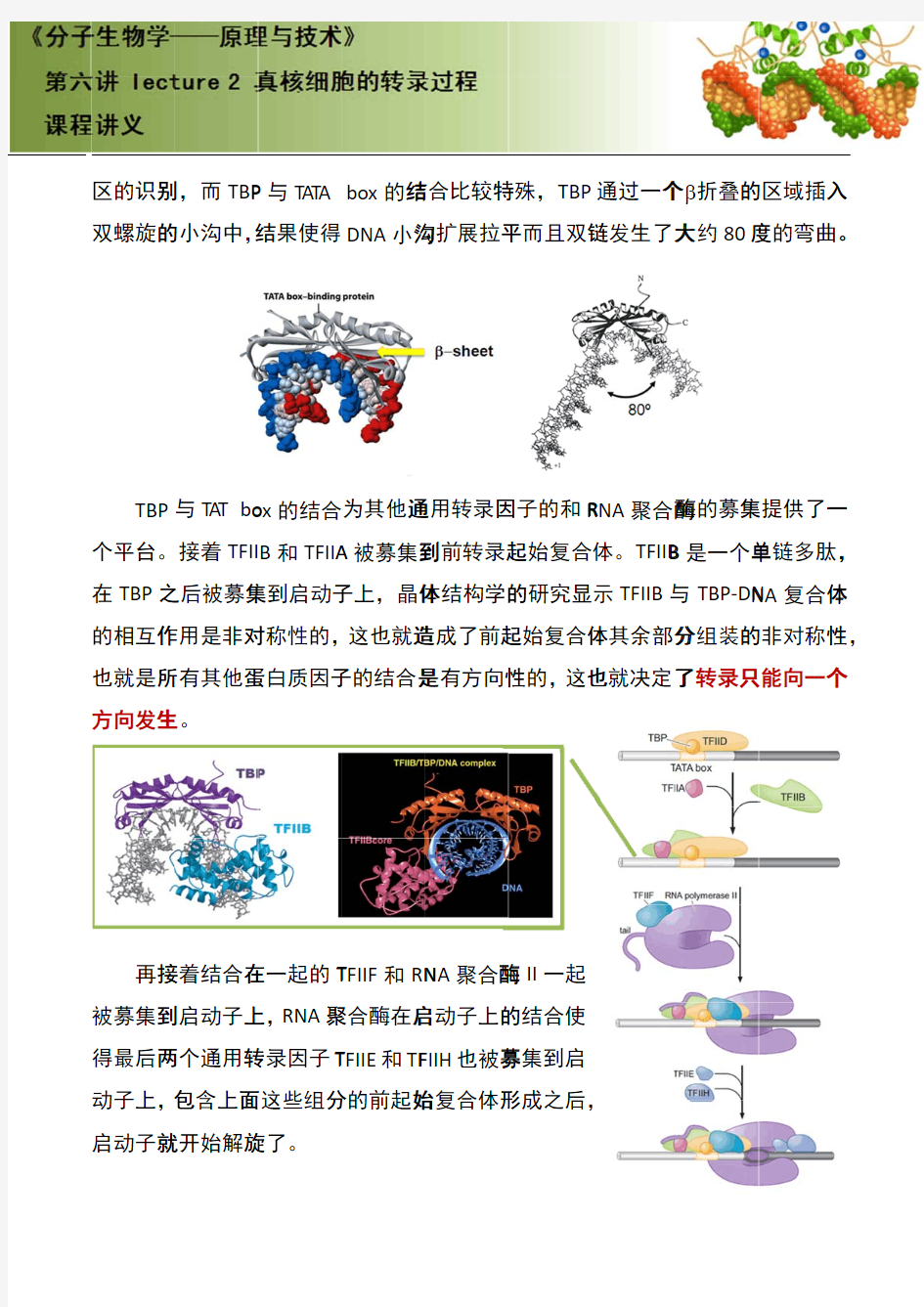
原核生物和真核生物中基因的转录
原核生物和真核生物中基因的转录、翻译和后修饰 摘要:原核生物和真核生物中基因的转录、翻译和后修饰,是各种功能蛋白质生物合成的一系列程序。本文通过介绍了原核生物和真核生物中基因的转录、翻译和后修饰的机制、原理、过程,从而了解真核生物和原核生物的基因表达和功能蛋白质合成上的差异。 关键词: 原核生物真核生物基因转录翻译后修饰 0引言: 21世纪,基因水平上的研究受到人们广泛的关注。原核生物和真核生物中基因的转录、翻译和后修饰是基础研究,人们也只有在此基础不断扩散深入研究其它基因水平问题。本文只简单介绍了一些关于基因转录、翻译和后修饰的一部分相关研究成果。 1 原核生物和真核生物中基因的转录: 基因转录是在由RNA聚合酶和辅助因子组成的转录复合物的催化下,从双链DNA分子中拷贝生物信息生成一条RNA链的过程。转录中,一个基因会被读取被复制为mRNA,就是说一特定的DNA片断作为模板,以DNA依赖的RNA合成酶作为催化剂的合成前体mRNA的过程。转录产物主要有三类RNA,即信使RNA (mRNA)、核糖体RNA(rRNA)和转移RNA(tRNA)。在基因转录过程中,RNA聚合酶起着非常重要的作用。RNA聚合酶可以催化所有四种核苷- 5′-三磷酸(ATP、GTP、UTP和CTP)聚合成与模板DNA互补的RNA。此反应需要Mg2+,反应中释放焦磷酸。[1]该酶在转录的各个过程中发挥了不同的作用。 1.1 基因转录的启动 RNA聚合酶正确识别DNA模板上的启动子并形成由酶、DNA和核苷三磷酸构成的三元起始复合物,转录便开始进行。启动子是DNA分子上可与RNA聚合酶特异结合,而使转录开始的一段DNA序列而本身不被转录。DNA模板上的启动区域常含有TATAATG顺序,称P盒。复合物中的核苷三磷酸一般为GTP,少数为ATP,因而原始转录产物的5′端通常为三磷酸鸟苷(pppG)或腺苷三磷酸(pppA)。真核DNA上的转录启动区域也有类似原核DNA的启动区结构,和在-30bp(即在酶和DNA结合点的上游30核苷酸处)附近也含有TATA结构,称TATA盒。[3]第一个核苷三磷酸与第二个核苷三磷酸缩合生成3′-5′磷酸二酯键后,则启动阶段结束,进入延伸阶段。 1.2 基因转录的延伸 σ亚基脱离酶分子,留下的核心酶与 DNA的结合变松,因而较容易继续往前移动。核心酶无模板专一性,能转录模板上的任何顺序,包括在转录后加工时待切除的居间顺序。脱离核心酶的σ亚基还可与另外的核心酶结合,参与另一
真核生物与原核生物转录与复制的区别
真核生物与原核生物转录 与复制的区别 This model paper was revised by the Standardization Office on December 10, 2020
不同点 真核生物和原核生物复制的不同点: 1.真核生物DNA的合成只是在细胞周期的S期进行,而原核生物则在整个细胞生长过程中都可进行DNA合成 2.原核生物DNA的复制是单起点的,而真核生物染色体的复制则为多起点的。真核生物中前导链的合成并不像原核生物那样是连续的,而是以半连续的方式,由一个复制起点控制一个复制子的合成,最后由连接酶将其连接成一条完整的新链。 3.真核生物DNA的合成所需的RNA引物及后随链上合成的冈崎片段的长度比原核生物要短。 4.原核生物中有DNA聚合酶Ⅰ、Ⅱ、Ⅲ三种聚合酶,并有DNA聚合酶Ⅲ同时控制两条链的合成。真核生物中有α、β、γ、ε、δ五种聚合酶。聚合酶α、δ是DNA 合成的主要酶,分别控制不连续的后随链以及前导链的生成。聚合酶β可能与DNA修复有关,聚合酶γ则是线粒体中发现的唯一一种DNA聚合酶. 5.染色体端体的复制不同。原核生物的染色体大多数为环状,而真核生物染色体为线状。末端有特殊DNA序列组成的结构成为端体。 真核生物和原核生物转录的不同点: 1.真核生物的转录在细胞核内进行,原核生物则在拟核区进行。 2.真核生物mRNA分子一般只编码一个基因,原核生物的一个mRNA分子通常含多个基因。
3.真核生物有三种不同的RNA聚合酶催化RNA合成,而在原核生物中只有一种RNA聚合酶催化所有RNA 的合成。 4.真核生物的RNA聚合酶不能独立转录RNA,三种聚合酶都必须在蛋白质转录因子的协助下才能进行RNA的转录,其RNA聚合酶对转录启动子的识别也比原核生物要复杂得多。原核生物的RNA聚合酶可以直接起始转录合成RNA。 真核生物和原核生物翻译的不同点: 的活化:起始氨基酸是,真核是从生成-tRNAi开始的。 翻译的起始:原核的起始tRNA是fMet-tRNA(fMet上角标),30s首先与mRNA模板相结合,再与fMet-tRNA(fMet上角标)结合,最后与50s结合。真核中起始tRNA是 Met-tRNA(Met上角标),40s小亚基首先与Met-tRNA(Met上角标)相结合,再与模板mRNA结合,最后与60s大亚基结合生成。 的延伸:没有区别 肽链的终止:原核含有三种释放因子RF1,RF2,RF3。真核只有eRF1和eRF3。 前体的加工蛋白质的折叠蛋白质的合成抑制这三步过程过于复杂,因具体物种而异 相同点 真核生物和原核生物复制的相同点: DNA复制 都是半保留复制、半不连续复制、双向复制,在复制中需要的原料、模板、引物都相同,都有前导链和滞后链,都分为起始、延伸、终止三个过程。
原核生物基因的转录的过程
原核生物基因的转录的 过程 Document number:NOCG-YUNOO-BUYTT-UU986-1986UT
原核生物基因的转录的过程 转录过程包括启动、延伸和终止。 启动RNA聚合酶正确识别DNA模板上的启动子并形成由酶、DNA 和核苷三磷酸(NTP)构成的三元起始复合物,转录即自此开始。DNA模板上的启动区域常含有TATAATG顺序,称普里布诺(Pribnow)盒或P盒。复合物中的核苷三磷酸一般为GTP,少数为ATP,因而原始转录产物的5′端通常为三磷酸鸟苷(pppG)或腺苷三磷酸(pppA)。真核DNA上的转录启动区域也有类似原核DNA的启动区结构,和在-30bp(即在酶和DNA结合点的上游30核苷酸处,常以—30表示,bp为碱基对的简写)附近也含有TATA结构,称霍格内斯(Hogness)盒或TATA盒。第一个核苷三磷酸与第二个核苷三磷酸缩合生成3′-5′磷酸二酯键后,则启动阶段结束,进入延伸阶段。 延伸σ亚基脱离酶分子,留下的核心酶与DNA的结合变松,因而较容易继续往前移动。核心酶无模板专一性,能转录模板上的任何顺序,包括在转录后加工时待切除的居间顺序。脱离核心酶的σ亚基还可与另外的核心酶结合,参与另一转录过程。随着转录不断延伸,DNA双链顺次地被打开,并接受新来的碱基配对,合成新的磷酸二酯键后,核心酶向前移去,已使用过的模板重新关闭起来,恢复原来的双链结构。一般合成的RNA链
对DNA模板具有高度的忠实性。RNA合成的速度,原核为25~50个核苷酸/秒,真核为45~100个核苷酸/秒。 终止转录的终止包括停止延伸及释放RNA聚合酶和合成的RNA。在原核生物基因或操纵子的末端通常有一段终止序列即终止子;RNA合成就在这里终止。原核细胞转录终止需要一种终止因子ρ(四个亚基构成的蛋白质)的帮助。真核生物DNA上也可能有转录终止的信号。已知真核DNA转录单元的3′端均含富有AT的序列〔如AATAA(A)或ATTAA(A)等〕,在相隔0~30bp之后又出现TTTT顺序(通常是3~5个T),这些结构可能与转录终止或者与3′端添加多聚A顺序有关。
真核生物转录特点
真核生物RNA的转录与原核生物RNA的转录过程在总体上基本相同,但是,其过程要复杂得多,主要有以下几点不同(图3-27)。 ⒈真核生物RNA的转录是在细胞核内进行的,而蛋白质的合成则是在细胞质内进行的。所以,RNA转录后首先必须从核内运输到细胞质内,才能指导蛋白质的合成。 ⒉真核生物一个mRNA分子一般只含有一个基因,原核生物的一个mRNA分子通常含有多个基因,而除少数较低等真核生物外,一个mRNA分子一般只含有一个基因,编码一条多态链。 ⒊真核生物RNA聚合酶较多在原核生物中只有一种RNA聚合酶,催化所有RNA的合成,而在真核生物中则有RNA聚合酶Ⅰ、RNA聚合酶Ⅱ和RNA聚合酶Ⅲ三种不同酶,分别催化不同种类型RNA的合成。三种RNA聚合酶都是由10个以上亚基组成的复合酶。RNA聚合酶Ⅰ存在于细胞核内,催化合成除5SrRNA 以外的所有rRNA的合成;RNA聚合酶Ⅱ催化合成mRNA前体,即不均一核RNA(hnRNA)的合成;RNA 聚合酶Ⅲ催化tRNA和小核RNA的合成。 ⒋真核生物RNA聚合酶不能独立转录RNA 。原核生物中RNA聚合酶可以直接起始转录合成RNA ,真核生物则不能。在真核生物中,三种RNA聚合酶都必须在蛋白质转录因子的协助下才能进行RNA的转录。另外,RNA聚合酶对转录启动子的识别,也比原核生物更加复杂,如对RNA聚合酶Ⅱ来说,至少有三个DNA的保守序列与其转录的起始有关,第一个称为TATA框(TATA box),具有共有序列TATAAAA,其位置在转录起始点的上游约为25个核苷酸处,它的作用可能与原核生物中的-10共有序列相似,与转录起始位置的确定有关。第二个共有序列称为CCAAT框(CCAAT box),具有共有序列GGAACCTCT,位于转录起始位置上游约为50-500个核苷酸处。如果该序列缺失会极大地降低生物的活体转录水平。第三个区域一般称为增强子(enhancer),其位置可以在转录起始位置的上游,也可以在下游或者在基因之内。它虽不直接与转录复合体结合,但可以显著提高转录效率。
原核与真核生物mRNA的特征比较
原核与真核生物mRNA的特征比较原核生物中: ?mRNA的转录和翻译发生在同一个细胞空间,?这两个过程几乎是同步进行的。 真核细胞中: 真核细胞mRNA的合成和功能表达发生在不同的空间和时间范畴内。 ?mRNA以较大分子量的前体RNA出现在核内,?只有成熟的、相对分子质量明显变小并经化学修饰的mRNA才能进入细胞质,参与蛋白质的合成。
mRNA的组成: ?编码区(coding region):从起始密码子AUG开始经一连串编码氨基酸的密码子直至终止密码子。 ?5’端上游非编码区(5’UTR):位于AUG之前不翻译的区域。 ?3’端下游非编码区(3’UTR):位于终止密码子之后不翻译的区域。
原核生物mRNA的特征 ?半衰期短。 ?许多原核生物mRNA以多顺反子的形式存在。 ?原核生物mRNA的5’端无帽子结构,3’端没有或只有较短的多聚(A)结构。
原核生物mRNA的特征 1.半衰期短 ?原核生物中,mRNA的 转录和翻译是在同一个 细胞空间里同步进行 的,蛋白质合成往往在 mRNA刚开始转录时就 被引发了。 ?大多数细菌mRNA在转 录开始1分钟后就开始降 解。mRNA降解的速度大 概只有转录或翻译速度的 一半。
原核生物mRNA的特征 2. 许多以多顺反子的形式存在: 原核细胞的mRNA(包括病毒)有时可以同时编码几个多肽。 Prokaryotic mRNA (polycistrionic)
单顺反子mRNA (monocistronic mRNA):只编码一个蛋白质的mRNA。 多顺反子mRNA(polycistronic mRNA):编码多个蛋白质的mRNA。
真核生物与原核生物转录与复制的区别
不同点 真核生物和原核生物复制的不同点: 1.真核生物DNA的合成只是在细胞周期的S期进行,而原核生物则在整个细胞生长过程中都可进行DNA合成 2.原核生物DNA复制是单起点的,而真核生物染色体的复制为多起点的。真核生物中前导链的合成并不像原核生物那样是连续的,而是以半连续的方式,由一个复制起点控制一个复制子的合成,最后由连接酶将其连接成一条完整的新链。 3.真核生物DNA的合成所需的RNA引物及后随链上合成的冈崎片段的长度比原核生物要短。 4.原核生物中有DNA聚合酶Ⅰ、Ⅱ、Ⅲ三种聚合酶,并有DNA聚合酶Ⅲ同时控制两条链的合成。真核生物中有α、β、γ、ε、δ五种聚合酶。聚合酶α、δ是DNA合成的主要酶,分别控制不连续的后随链以及前导链的生成。聚合酶β可能与DNA修复有关,聚合酶γ则是线粒体中发现的唯一一种DNA聚合酶. 5.染色体端粒的复制不同。原核生物的染色体大多数为环状,而真核生物染色体为线状。末端有特殊DNA序列组成的结构成为端粒。 真核生物和原核生物转录的不同点: 1.真核生物的转录在细胞核内进行,原核生物则在拟核区进行。 2.真核生物mRNA分子一般只编码一个基因,原核生物的一个mRNA分子通常含多个基因。 3.真核生物有三种不同的RNA聚合酶催化RNA合成,而在原核生物中只有一种RNA聚合酶催化所有RNA 的合成。 4.真核生物的RNA聚合酶不能独立转录RNA,三种聚合酶都必须在蛋白质转录因子的协助下才能进行RNA的转录,其RNA聚合酶对转录启动子的识别也比原核生物要复杂得多。原核生物的RNA聚合酶可以直接起始转录合成RNA。真核生物和原核生物翻译的不同点: 氨基酸的活化:原核起始氨基酸是甲酰甲硫氨酸,真核是从生成甲硫氨酰-tRNAi开始的。翻译的起始:原核的起始tRNA是tRNA fMet,30s小亚基首先与mRNA模板相结合,再与tRNA fMet结合,最后与50s大亚基结合。真核中起始tRNA是tRNA Met,40s小亚基首先与tRNA Met相结合,再与模板mRNA结合,最后与60s大亚基结合生成起始复合物。 肽链的延伸:没有区别 肽链的终止:原核含有三种释放因子RF1,RF2,RF3。真核只有eRF1和eRF3。 蛋白质前体的加工蛋白质的折叠蛋白质的合成抑制这三步过程过于复杂,因具体物种而异。 相同点 真核生物和原核生物复制的相同点: DNA复制 都是半保留复制、半不连续复制、双向复制,在复制中需要的原料、模板、引物都相同,都有前导链和滞后链,都分为起始、延伸、终止三个过程。 RNA转录:
原核生物基因的转录的过程
原核生物基因的转录的过程 转录过程包括启动、延伸和终止。 启动RNA聚合酶正确识别DNA模板上的启动子并形成由酶、DNA和核苷三磷酸(NTP)构成的三元起始复合物,转录即自此开始。DNA模板上的启动区域常含有TATAATG顺序,称普里布诺(Pribnow)盒或P盒。复合物中的核苷三磷酸一般为GTP,少数为ATP,因而原始转录产物的5′端通常为三磷酸鸟苷(pppG)或腺苷三磷酸(pppA)。真核DNA上的转录启动区域也有类似原核DNA的启动区结构,和在-30bp(即在酶和DNA结合点的上游30核苷酸处,常以—30表示,bp为碱基对的简写)附近也含有TATA结构,称霍格内斯(Hogness)盒或TATA盒。第一个核苷三磷酸与第二个核苷三磷酸缩合生成3′-5′磷酸二酯键后,则启动阶段结束,进入延伸阶段。 延伸σ亚基脱离酶分子,留下的核心酶与DNA的结合变松,因而较容易继续往前移动。核心酶无模板专一性,能转录模板上的任何顺序,包括在转录后加工时待切除的居间顺序。脱离核心酶的σ亚基还可与另外的核心酶结合,参与另一转录过程。随着转录不断延伸,DNA双链顺次地被打开,并接受新来的碱基配对,合成新的磷酸二酯键后,核心酶向前移去,已使用过的模板重新关闭起来,恢复原来的双链结构。一般合成的RNA链对DNA模板具有高度的忠实性。RNA合成的速度,原核为25~50个核苷酸/秒,真核为45~100个核苷酸/秒。 终止转录的终止包括停止延伸及释放RNA聚合酶和合成的RNA。在原核生物基因或操纵子的末端通常有一段终止序列即终止子;RNA合成就在这里终止。原核细胞转录终止需要一种终止因子ρ(四个亚基构成的蛋白质)的帮助。真核生物DNA上也可能有转录终止的信号。已知真核DNA转录单元的3′端均含富有AT的序列〔如AATAA(A)或ATTAA(A)等〕,在相隔0~30bp之后又出现TTTT顺序(通常是3~5个T),这些结构可能与转录终止或者与3′端添加多聚A顺序有关。
11-生物化学习题与解析--RNA的生物合成过程
11-生物化学习题与解析--RNA 的生物合成过程
RNA的生物合成过程 一、选择题 (一) A 型题 1 .下列关于转录的叙述正确的是 A .转录过程需 RNA 引物 B .转录生成的 RNA 都是翻译模板 C .真核生物转录是在胞浆中进行的 D . DNA 双链一股单链是转录模板 E . DNA 双链同时作为转录模板 2 . DNA 上某段编码链碱基顺序为 5 ' -ACTAGTCAG- 3 ' ,转录后 mRNA 上相应的碱基顺序为 A . 5 ' -TGATCAGTC-3 ' B . 5 ' -UGAUCAGUC-3 ' C . 5 ' -CUGACUAGU-3 ' D . 5 ' -CTGACTAGT-3 ' E . 5 ' -CAGCUGACU-3 ' 3 .不对称转录是 A .双向复制后的转录 B .以 DNA 为模板双向进行转录 C .同一单链 DNA ,转录时可以交替作为编码链和模板链 D .同一单链 DNA ,转录时只转录外显子部分 E .没有规律的转录 4 .真核生物的转录特点是 A .发生在细胞质内,因为转录产物主要供蛋白质合成用 B .转录产物有 poly ( A )尾, DNA 模板上有相应的 poly ( dT )序列 C .转录的终止过程需ρ( Rho )因子参与 D .转录起始需要形成 PIC (转录起始前复合物) E .需要α因子辨认起点 5 .下列关于转录编码链的叙述正确的是 A .能转录生成 mRNA 的 DNA 单链 B .能转录生成 tRNA 的 DNA 单链 C .同一 DNA 单链不同片段可作模板链或编码链 D .是基因调节的成份 E .是 RNA 链 6 . Pribnow box 序列是 A . AAUAAA B . TAAGG C C . TTGACA D . TATAAT E . AATAAA 7 .真核生物的 TATA 盒是 A .参与转录起始 B .翻译的起始点 C . RNA 聚合酶核心酶结合位点 D .σ因子结合位点 E .复制的起始点 8 .原核生物 DNA 指导的 RNA 聚合酶由数个亚基组成,其核心酶的组成是 A .α 2 ββ ' ( ω ) B .α 2 β ( σ ) C .α 2 ββ ' σ ( ω ) D .α 2 β ' ( ω ) E .αββ ' 9 .原核生物识别转录起始点的是 A .ρ因子 B .核心酶 C . RNA 聚合酶的α亚基 D .σ亚基 E . RNA 聚合酶的β 亚基 10 .ρ因子的功能是 A .参与转录的启动过程 B .参与转录的全过程 C .加速 RNA 的合成 D .参与转录的终止过程 E .可改变 RNA 聚合酶的活性 11 .在转录延长阶段, RNA 聚合酶与 DNA 模板的结合是 A .全酶与模板结合 B .核心酶与模板特定位点结合 C .结合松弛而有利于 RNA 聚合酶向前移动
生物化学习题与解析RNA的生物合成过程
R N A的生物合成过程 一、选择题 (一)A型题 1.下列关于转录的叙述正确的是 A.转录过程需RNA引物B.转录生成的RNA都是翻译模板 C.真核生物转录是在胞浆中进行的D.DNA双链一股单链是转录模板 E.DNA双链同时作为转录模板 2.DNA上某段编码链碱基顺序为5'-ACTAGTCAG-3',转录后mRNA上相应的碱基顺序为 A.5'-TGATCAGTC-3'B.5'-UGAUCAGUC-3' C.5'-CUGACUAGU-3'D.5'-CTGACTAGT-3' E.5'-CAGCUGACU-3' 3.不对称转录是 A.双向复制后的转录B.以DNA为模板双向进行转录 C.同一单链DNA,转录时可以交替作为编码链和模板链 D.同一单链DNA,转录时只转录外显子部分 E.没有规律的转录 4.真核生物的转录特点是 A.发生在细胞质内,因为转录产物主要供蛋白质合成用 B.转录产物有poly(A)尾,DNA模板上有相应的poly(dT)序列 C.转录的终止过程需ρ(Rho)因子参与 D.转录起始需要形成PIC(转录起始前复合物) E.需要α因子辨认起点 5.下列关于转录编码链的叙述正确的是 A.能转录生成mRNA的DNA单链B.能转录生成tRNA的DNA单链 C.同一DNA单链不同片段可作模板链或编码链 D.是基因调节的成份E.是RNA链 6.Pribnowbox序列是A.AAUAAAB.TAAGGCC.TTGACAD.TATAATE.AATAAA 7.真核生物的TATA盒是 A.参与转录起始B.翻译的起始点C.RNA聚合酶核心酶结合位点 D.σ因子结合位点E.复制的起始点 8.原核生物DNA指导的RNA聚合酶由数个亚基组成,其核心酶的组成是 A.α2ββ'(ω)B.α2β(σ)C.α2ββ'σ(ω)D.α2β'(ω)E.αββ' 9.原核生物识别转录起始点的是 A.ρ因子B.核心酶C.RNA聚合酶的α亚基 D.σ亚基E.RNA聚合酶的β亚基 10.ρ因子的功能是 A.参与转录的启动过程B.参与转录的全过程C.加速RNA的合成 D.参与转录的终止过程E.可改变RNA聚合酶的活性 11.在转录延长阶段,RNA聚合酶与DNA模板的结合是
原核生物基因的转录的过程
原核生物基因的转录的过程转录过程包括启动、延伸和终止。 启动RNA聚合酶正确识别DNA模板上的启动子并形成由酶、DNA和核苷三磷酸(NTP)构成的三元起始复合物,转录即自此开始。DNA模板上的启动区域常含有TATAATG顺序,称普里布诺(Pribnow)盒或P盒。复合物中的核苷三磷酸一般为GTP,少数为ATP,因而原始转录产物的5′端通常为三磷酸鸟苷(pppG)或腺苷三磷酸(pppA)。真核DNA上的转录启动区域也有类似原核DNA的启动区结构,和在-30bp(即在酶和DNA结合点的上游30核苷酸处,常以—30表示,bp为碱基对的简写)附近也含有TATA结构,称霍格内斯(Hogness)盒或TATA盒。第一个核苷三磷酸与第二个核苷三磷酸缩合生成3′-5′磷酸二酯键后,则启动阶段结束,进入延伸阶段。 延伸σ亚基脱离酶分子,留下的核心酶与DNA的结合变松,因而较容易继续往前移动。核心酶无模板专一性,能转录模板上的任何顺序,包括在转录后加工时待切除的居间顺序。脱离核心酶的σ亚基还可与另外的核心酶结合,参与另一转录过程。随着转录不断延伸,DNA 双链顺次地被打开,并接受新来的碱基配对,合成新的磷酸二酯键后,核心酶向前移去,已使用过的模板重新关闭起来,恢复原来的双链结构。一般合成的RNA链对DNA模板具有高度的忠实性。RNA合成的速度,原核为25~50个核苷酸/秒,真核为45~100个核苷酸/秒。
终止转录的终止包括停止延伸及释放RNA聚合酶和合成的RNA。在原核生物基因或操纵子的末端通常有一段终止序列即终止子;RNA合成就在这里终止。原核细胞转录终止需要一种终止因子ρ(四个亚基构成的蛋白质)的帮助。真核生物DNA上也可能有转录终止的信号。已知真核DNA转录单元的3′端均含富有AT的序列〔如AATAA(A)或ATTAA(A)等〕,在相隔0~30bp之后又出现TTTT顺序(通常是3~5个T),这些结构可能与转录终止或者与3′端添加多聚A顺序有关。
原核生物基因的转录的过程复习过程
精品文档 精品文档原核生物基因的转录的过程 转录过程包括启动、延伸和终止。 启动RNA聚合酶正确识别DNA模板上的启动子并形成由酶、DNA和核苷三磷酸(NTP)构成的三元起始复合物,转录即自此开始。DNA模板上的启动区域常含有TATAATG顺序,称普里布诺(Pribnow)盒或P盒。复合物中的核苷三磷酸一般为GTP,少数为ATP,因而原始转录产物的5′端通常为三磷酸鸟苷(pppG)或腺苷三磷酸(pppA)。真核DNA上的转录启动区域也有类似原核DNA的启动区结构,和在-30bp(即在酶和DNA结合点的上游30核苷酸处,常以—30表示,bp为碱基对的简写)附近也含有TATA结构,称霍格内斯(Hogness)盒或TATA盒。第一个核苷三磷酸与第二个核苷三磷酸缩合生成3′-5′磷酸二酯键后,则启动阶段结束,进入延伸阶段。 延伸σ亚基脱离酶分子,留下的核心酶与DNA的结合变松,因而较容易继续往前移动。核心酶无模板专一性,能转录模板上的任何顺序,包括在转录后加工时待切除的居间顺序。脱离核心酶的σ亚基还可与另外的核心酶结合,参与另一转录过程。随着转录不断延伸,DNA双链顺次地被打开,并接受新来的碱基配对,合成新的磷酸二酯键后,核心酶向前移去,已使用过的模板重新关闭起来,恢复原来的双链结构。一般合成的RNA链对DNA模板具有高度的忠实性。RNA合成的速度,原核为25~50个核苷酸/秒,真核为45~100个核苷酸/秒。 终止转录的终止包括停止延伸及释放RNA聚合酶和合成的RNA。在原核生物基因或操纵子的末端通常有一段终止序列即终止子;RNA合成就在这里终止。原核细胞转录终止需要一种终止因子ρ(四个亚基构成的蛋白质)的帮助。真核生物DNA上也可能有转录终止的信号。已知真核DNA转录单元的3′端均含富有AT的序列〔如AATAA(A)或ATTAA(A)等〕,在相隔0~30bp之后又出现TTTT顺序(通常是3~5个T),这些结构可能与转录终止或者与3′端添加多聚A顺序有关。
原核生物和真核生物基因表达调控复制、转录、翻译特点的比较
1.相同点:转录起始是基因表达调控的关键环节 ①结构基因均有调控序列; ②表达过程都具有复杂性,表现为多环节; ③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性; 2.不同点: ①原核基因的表达调控主要包括转录和翻译水平。真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。 ②原核基因表达调控主要为负调控,真核主要为正调控。 ③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。 ④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。 ⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。原核生物基因以操纵子的形式存在。转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。 真核生物基因表达的调控环节较多: 在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。 在转录水平主要通过反式作用因子调控转录因子与TATA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。 在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。 在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。 真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。 真核生物和原核生物复制的不同点: ①真核生物DNA的合成只是在细胞周期的S期进行,而原核生物则在整个细胞生长过程中都可进行DNA合成 ②原核生物DNA的复制是单起点的,而真核生物染色体的复制则为多起点的。真核生物中前导链的合成并不像原核生物那样是连续的,而是以半连续的方式,由一个复制起点控制一个复制子的合成,最后由连接酶将其连接成一条完整的新链。 ③真核生物DNA的合成所需的RNA引物及后随链上合成的冈崎片段的长度比原核生物要短。