第10章 酶的作用机制和酶的调节

第十章酶的作用机制和酶的调节
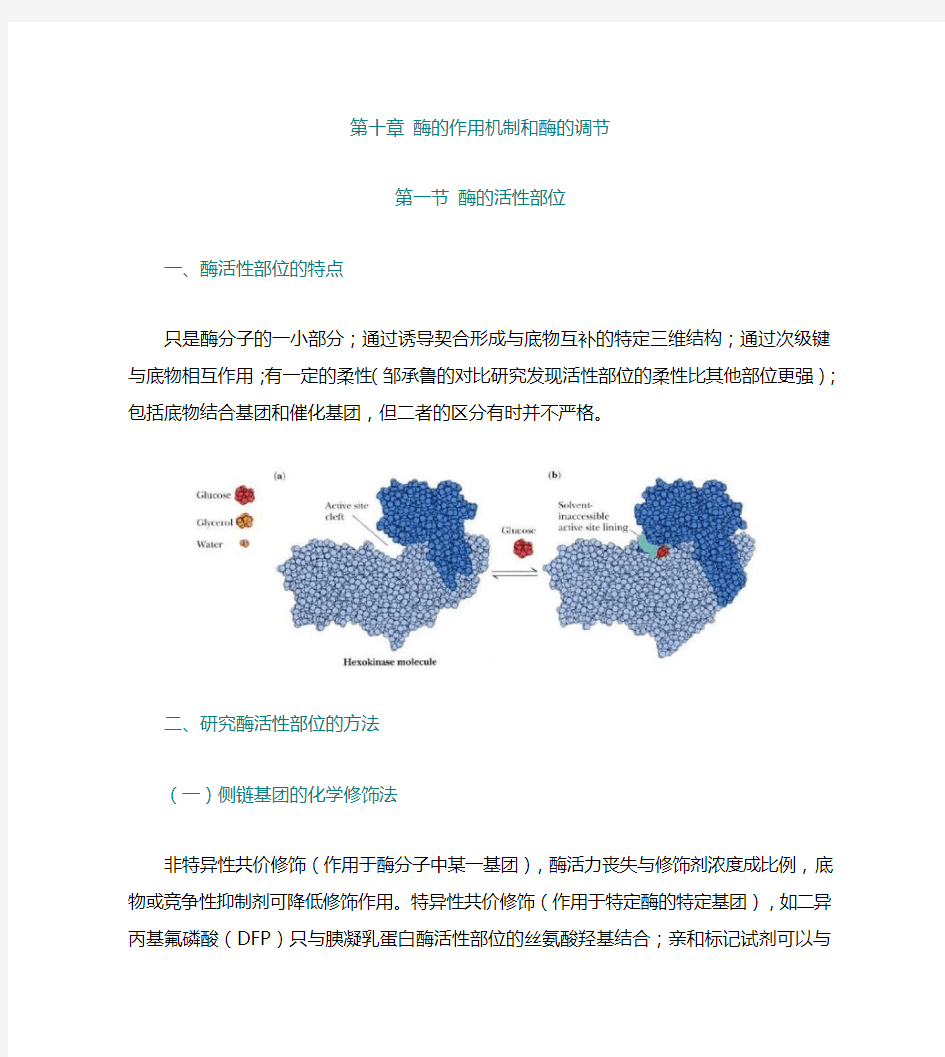
第一节酶的活性部位
一、酶活性部位的特点
只是酶分子的一小部分;通过诱导契合形成与底物互补的特定三维结构;通过次级键与底物相互作用;有一定的柔性(邹承鲁的对比研究发现活性部位的柔性比其他部位更强);包括底物结合基团和催化基团,但二者的区分有时并不严格。
二、研究酶活性部位的方法
(一)侧链基团的化学修饰法
非特异性共价修饰(作用于酶分子中某一基团),酶活力丧失与修饰剂浓度成比例,底物或竞争性抑制剂可降低修饰作用。特异性共价修饰(作用于特定酶的特定基团),如二异丙基氟磷酸(DFP)只与胰凝乳蛋白酶活性部位的丝氨酸羟基结合;亲和标记试剂可以与活性部位的特定基团共价定量结合,如对甲苯磺酰-L-苯丙氨酰氯甲基酮(TPCK)与胰凝乳蛋白酶活性部位丝氨酸羟基的结合。
(二)动力学参数测定法
(三)X-射线晶体结构分析法
(四)定点诱变法
第二节酶催化反应的独特性质
第三节影响酶催化效率的有关因素
一、底物和酶的邻近效应(approximation,proximity)与定向效应(orientation)
底物分子结合到酶的活性中心,使底物在酶活性中心的有效浓度大大增加,从而加快反应的速度。
一个有机化学模型。咪唑催化对硝基苯酯的水解,分子内反应比分子间反应快24 倍。
底物分子中参与反应的基团相互接近,并被严格定向定位,使酶促反应具有高效率称定向效应。邻羟基苯丙酸内脂的形成反应,两个甲基使羧基和羟基更好的定向,使反应速率提高2.5×1011。
二、底物的形变(distortion)和诱导契合(inducednt)
脯氨酸消旋酶的过渡态类似物与酶的亲和力远大于其底物,说明酶可以引起底物的形变。
酵母醛缩酶的过渡态类似物与酶的亲和力远大于其底物,说明酶可以引起底物的形变。
小牛小肠腺苷脱氨酶的过渡态类似物与酶的亲和力远大于其底物,说明酶可以引起底物的形变。
三、酸碱催化(acid—base catalysis)
专一性(狭义)酸碱催化
总酸碱催化或广义酸碱催化
表观速度常数依赖于pH 和溶液浓度。
Brφnsted 催化作用定律:
Logka=CA ? α(pKa)
pKa 和pKb 是总酸碱的解离常数,ka 和kb 是反应的速度常数。CA 和CB 为常数,与反应类型、温度、溶剂等因素有关。
pKa 小,则H+浓度高。
α和β是Brφnsted 系数,范围在0 到1 之间,接近于1 为专一性酸碱催化,接近于0 表示酸碱催化不起作用。
咪唑基的pK 值接近生理pH 值,H+的传递速度快,是酸碱催化中最重要的基团。
四、共价催化
酶与底物形成共价键的3 个例子,其共同特点是酶的亲核中心X 进攻底物的亲电中心。
五、金属离子催化
金属酶含紧密结合的金属离子。金属激活酶含松散结合的金属离子。
金属离子的作用主要有:
与结合底物为反应定向;
参与氧化还原反应;
稳定或屏蔽负电荷;
活化水分子;
与过渡态底物形成螯合物。
六、多元催化和协同效应
酶的催化过程通常是多种机制同时作用,相互协同,因而有很高的催化效率。
七、活性部位微环境的影响
第四节酶催化反应机制的实例一、溶菌酶(1ysozyme)
二、胰核糖核酸酶A(pancreatic ribonuclease A,RNase A)
四、丝氨酸蛋白酶(serine proteases)
胰凝乳蛋白酶、胰蛋白酶、弹性蛋白酶氨基酸序列的比较,黑点表示相同的氨基酸,活性部位的氨基酸用符号标出。
胰凝乳蛋白酶和水蛭蛋白酶抑制剂(蓝色带状)的结构,His57(蓝色),Asp102(红色),Ser195(黄色)在空间上十分靠近。
胰凝乳蛋白酶催化的裂解反应的动力学。
乙酸基以共价键与酶的Ser195 结合,因而释放较晚。酰基-酶中间物快速形成,随后缓慢释放产物。
胰凝乳蛋白酶催化反应的机制:
五、天冬氨酸蛋白酶(asparticproteases)
若为共价催化,会有两种中间物之一,两种中间物均未能找到,说明这一类酶为非共价催化。
(a)HIV-1 protease,a dimer,and (b) pepsin (a monomer).Pepsin′s N-terminal half is shown in red;C-terminal half is shown in blue.
HIV-1 protease complexed with the inhibitor CrixivanR(red) the flaps(residues 46-55 from each subunit)covering the active site are shown in green and the active site aspartate residues involved in catalysis are shown inwhite.
第五节酶活性的调节控制
一、别构调控(allos teri regulation)
(一)别构酶的性质
别构酶均为寡聚酶,除活性部位外,还有可以同效应物(调节物)结合的调节部位。别构酶的调控方式有四类:
正协同效应:酶的一个亚基与底物结合后,其他亚基与底物的结合能力加强,v-[S]曲线为S 形。负协同效应:酶的一个亚基与底物结合后,其他亚基与底物的结合能力减弱,v-[S]曲线为平坦的双曲线。
可用Rs ([S]90%V/[S]10%V)来定量地区分三种酶:Rs 等于81 为米氏酶,大于81 则有正协同效应,小于81 为负协同效应。
更常用的是Hill 系数法,以log(v/(Vm-v))对log[S]作图,曲线的最大斜率为Hill 系数,米氏酶等于1,正协同酶大于1,负协同小于1。
别构抑制作用使酶的反应速度降低,正协同效应加强。
别构激活作用使酶的反应速度加快,正协同效应减弱。
目前认为齐变模型不适合于负协同效应。
(二)天冬氨酸转氨甲酰酶的性质天冬氨酸为正协同效应剂;
ATP 为别构激活剂;
CTP 为别构抑制剂。
第10章 酶的作用机制和酶的调节
第10章酶的作用机制和酶的调节 教学目的:掌握酶的活性部位结构与功能、酶活性的别构调节、酶原激活,了解酶高效性原因 教学重点:酶活性部位的结构与功能及酶的活性的别构调节 教学难点:酶活性的别构调节 教学方法:多媒体 教学内容: 一、酶的活性部位及确定方法 (一)酶活性部位概念及特点 1、酶的活性中心(活性部位):指酶分子中的表面有一个必需基团比较集中、并构成一定空间结构的微小区域。酶活性中心的基团,按其功能可分为结合基团和催化基团。 活性中心的基团都是维持酶活性的必需基团, 2、酶活性部位的共同点: (1)酶活性部位仅占酶体积的很小一部分,通常只占整个酶分子体积的1~2%,酶分子是大分子物质,由很多氨基酸构成,而活性部位仅由几个氨基酸残基组成 催化部位一般由2~3个氨基酸残基组成。 结合部位氨基酸残基数目,不同的酶有所不同。可能是一个,也可能是多个。 (2)酶的活性部位具有三维结构,构成酶活性中心的基团,可位于同一条肽链上,也可位于不同的肽链上,在一级结构上可能相距甚远,但在空间结构上位置必须相互靠近;酶的空间结构受物理或化学因素影响时,酶的活性部位可能会遭破坏,酶会失活。(3)活性中心的结合基团与底物专一性结合,这需要活性部位的基团精确排列。活性部位具有一定的柔韧性,活性部位的结构并不是与底物的结构正好互补。 在酶与底物结合过程中,酶活性中心的构象在底物的诱导下可发生形变,然后嵌合互补形成中间产物,而底物在酶活性中心的诱导下也可发生形变,变的易与酶结合,有时是两者的构象同时发生变化后才互补契合(诱导契合学说)。 (4)酶活性部位位于酶分子表面的一个裂缝内,底物分子或底物分子的一部分结合到裂缝中,裂缝内的非极性基团较多,形成一个疏水环境,提高与底物的结合能力,也有极性的氨基酸残基,以便与底物结合并催化底物发生反应。 (5)底物通过较弱的次级键与酶结合。 组成酶活性中心的氨基酸残基,常见的有:组氨酸、赖氨酸、天冬氨酸、谷氨酸、丝氨酸、半胱氨酸和酪氨酸 3、研究酶活性部位的方法 (1)共价修饰 (2)亲和标记法 (3)切除法 (4)X射线晶体结构分析法 二、酶促反应机制 (一)基元催化的分子机制:酶的催化作用包括若干基元催化。 1、酸碱催化 2、共价催化 3、金属离子催化 (二)酶具有高催化能力的原因 1、底物与酶的邻近效应与定向效应 2 、底物分子中的敏感键发生“张力”与“形变”
10第十章 酶的作用机制和酶的调节
第十章酶的作用机制和酶的调节 目的和要求:理解、掌握酶活性部位的相关概念和特点;掌握酶催化高效性的相关机理;了解几种酶的催化机制,理解结构和功能的适应性;了解酶活性的调节方式,掌握酶活性的别构调节、可逆共价调节和酶原激活调节方式及生物代谢中的作用。 一、酶的活性部位 ㈠酶的活性部位的特点 1、概念:三维结构上比较接近的少数特异的氨基酸残基参与底物的结合与催化作用,这一与酶活力直 接相关的区域称酶的活性部位。 结合部位:专一性 催化部位:催化能力,对需要辅酶的酶分子,辅酶或其一部分就是活性中心的组成部分;组成酶活性部位的氨基酸数目对不同酶而言存在差异,占整个酶氨基酸残基小部分 酶活性部位的基团:亲核性基团,丝氨酸的羟基,半胱氨酸的巯基和组氨酸的咪唑基。酸碱性基团:天冬氨酸和谷氨酸的羧基,赖氨酸的氨基,酪氨酸的酚羟基,组氨酸的咪唑基和半胱氨酸的巯基等。 2、特点 ⑴活性部位在酶分子的总体中只占相当小的部分(1%~2%) ⑵酶的活性部位是一个三维实体 ⑶酶的活性部位并不是和底物的形状互补的 ⑷酶的活性部位是位于酶分子表面的一个裂隙内 ⑸底物通过次级键结合到酶上 ⑹酶活性部位具有柔性 ㈡研究酶活性部位的方法 1、酶分子基团的侧链化学修饰 ⑴非特异性共价修饰:活力丧失程度与修饰剂浓度有正比关系;底物或可逆的抑制剂可保护共价修饰 剂的修饰作用。 ⑵特异性共价修饰:分离标记肽段,可判断活性部位的氨基酸残基,如二异丙基氟磷酸(DFP)专一 性与胰凝乳蛋白酶活性部位丝氨酸残基的羟基结合。 ⑶亲和标记:利用底物类似物和酶活性部位的特殊亲和力将酶加以修饰标记来研究酶活性部位的方法。 修饰剂的特点:①结构与底物类似,能专一性引入到酶活性部位;②具活泼化学基团,能与活性部位某一氨基酸共价结合,相应的试剂称“活性部位指示剂”。 胰凝乳蛋白酶和胰蛋白酶,TPE是酶的底物,TPCK是酶的亲和试剂,当酶与TPCK温浴后,酶活性丧失,这种结合具有空间结构的需求,同时也阻止其他试剂如DFP结合。对酶活性中心的组氨酸咪唑环进行修饰。 ⑷自杀性底物标记:底物与酶结合并被催化所生成的产物可以与活性部位的基团专一性结合从而抑制 酶的催化活性。 2、动力学参数测定法:通过动力学方法求得相关参数,作出相应判断。 3、X-射线晶体衍射法:如溶菌酶和胰蛋白酶活性中心的测定
酶作用机理和调节【生物化学】
酶作用机理和调节 一、选择题 ⒈关于酶活性中心的描述,哪一项正确?() A、所有的酶都有活性中心; B、所有酶的活性中心都含有辅酶; C、酶的必须基团都位于酶的活性中心内; D、所有的抑制剂都是由于作用于酶的活性中心; E、所有酶的活性中心都含有金属离子 ⒉酶分子中使底物转变为产物的基团是指:() A、结合基团; B、催化基团; C、疏水基团; D、酸性基团; E、碱性基团
⒊酶原的激活是由于:() A、氢键断裂,改变酶分子构象; B、酶蛋白和辅助因子结合; C、酶蛋白进行化学修饰; D、亚基解聚或亚基聚合; E、切割肽键,酶分子构象改变 ⒋同工酶是指() A、辅酶相同的酶; B、活性中心的必需基团相同的酶; C、功能相同而分子结构不同的酶; D、功能和性质都相同的酶; E、功能不同而酶分子结构相似的酶 ⒌有关别构酶的结构特点,哪一项不正确?() A、有多个亚基; B、有与底物结合的部位; C、有与调节物结合的部位; D、催化部位和别
构部位都位于同一亚基上;E、催化部位与别构部位既可以处于同一亚基也可以处于不同亚基上。 ⒍属于酶的可逆性共价修饰,哪项是正确的? A、别构调节; B、竞争性抑制; C、酶原激活; D、酶蛋白和辅基结合; E、酶的丝氨酸羟基磷酸化 ⒎溶菌酶在催化反应时,下列因素中除哪个外,均与酶的高效率有关?() A、底物形变; B、广义酸碱共同催化; C、临近效应与轨道定向; D、共价催化; E、无法确定 ⒏对具有正协同效应的酶,其反应速度为最大反应速度0.9时底物浓度([S]0.9)与最大反应
旗开得胜速度为0.1时的底物浓度([S]0.1)二者的比值[S]0.9/[S]0.1应该为() A、>81; B、=81; C、<81; D、无法确定 ⒐以Hill系数判断,则具负协同效应的别构酶() A、n>1; B、n=1; C、n<1; D、n≥1; E、n≤1
酶的作用机制和酶的调节
酶的作用机制和酶的调节 重点综述 1. 酶作用机制:有专一性机理(锁与钥匙学说和诱导契和假说)和高效性的机理,以后者出现偏多,而且考查的题型上也是多样化(填写、选择、判断、问答等)。 (1)酶作用机理的两种学说,可以只作一般性的了解。 (2)酶作用高效性的机理要重点掌握。体现在以下5个方面:①靠近与定向;②变形与扭曲;③共价催化;④酸碱催化;⑤酶活性部位的低介电区。 在这一部分中,还要了解某些酶的作用原理: ①溶菌酶:活性部位有Clu3,和ASP52典型的酸碱催化。 ②胰凝乳蛋白酶:活性部位有ASPl02、His57和Serl95组成的电荷拉力网。 ③羧肽酶A:含金属离子zn2+的酶。 2. 酶的调节:酶调节的类型(共价调节,化学修饰,酶原激活,酶含量在分子水平的调节)。 几个概念也很重要:别构酶,调节酶等。 (一)名词解释 1.变构酶(allosteric enzyme);2.同工酶(isozyme);3.活性中心(active center);4. 酶原的激活(activation of zymogen); 5. 别构效应(allosteric effect); 6. 正协同效应(positive cooperative effect) (二)选择题(在备选答案中选出1个或多个正确答案) 1. 酶原激活的实质是 A. 激活剂与酶结合使酶激活 B. 酶蛋白的变构效应 C. 酶原分子一级结构发生改变从而形成或暴露出酶的活性中心 D. 酶原分子的空间构象发生了变化而一级结构不变 E. 以上都不对 2. 同工酶的特点是 A. 催化相同的反应,但分子结构和理化性质不同的一类酶 B. 催化相同反应,分子组成相同,但辅酶不同的一类酶 C. 催化同一底物起不同反应的酶的总称 D. 多酶体系中酶组分的统称 E. 催化作用,分子组成及理化性质相同,但组织分布不同的酶 3. 乳酸脱氢酶(LDH)是一个由两种不同的亚基组成的四聚体。假定这些亚基随机结合成四聚体,这种酶有多少种同工酶? A. 两种 B. 三种 C. 四种 D. 五种 E. 六种 4.下列关于酶活性中心的叙述哪些是正确的 A.是由一条多肽链中若干相邻的氨基酸残基以线状排列而成 B.对于整个酶分子来说,只是酶的一小部分 C.仅通过共价键与作用物结合
生物化学(第三版)第十章 酶的作用机制和酶的调节课后习题详细解答_ 复习重点
第十章酶的作用机制和酶的调节 提要 酶的活性部位对于不需要辅酶的酶来说,就是指酶分子中在三维结构上比较靠近的几个氨基酸残基负责与底物的结合与催化作用的部位,对于需要辅酶的酶来说,辅酶分子或辅酶分子上的某一部分结构,往往也是酶活性部位的组成部分。酶活性部位有6个共同特点。研究酶活性部位的方法有:酶分子侧链基团的化学修饰法,动力学参数测定法,X射线晶体结构分析法和定点诱变法,这些方法可互相配合以判断某个酶的活性部位。 酶是催化效率很高的生物催化剂,这是由酶分子的特殊结构所决定的。经研究与酶催化效率的有关因素有7个,即底物和酶的邻近效应与定向效应,底物的形变与诱导契合,酸碱催化,共价催化,金属离子催化,多元催化和协同效应,活性部位微环境的影响。但这些因素不是同时在一个酶中其作用,也不是一种因素在所有的酶中起作用,对于某一种酶来说,可能分别主要受一种或几种因素的影响。 研究酶催化的反应机制,始终是酶学研究的一个重点,通过大量的研究工作,已经对一些酶的作用机制有深入了解,该章对溶解酶、胰核糖核酸酶A、羧肽酶A、丝氨酸蛋白酶、天冬氨酸蛋白酶等的催化作用机制进行了详尽的讨论。 酶活性是受各种因素调节控制的,除了在第8章中已介绍的几种因素外,主要还有①别构调节,例如ATCase。②酶原的激活,如消化系统蛋白酶原的激活及凝血系统酶原的激活。③可逆共价修饰调控,如蛋白质的磷酸化,一系列蛋白激酶的作用。通过以上作用,使酶能在准确的时间和正确的地点表现出它们的活性。别构酶一般都是寡聚酶,有催化部位和调节部位,别构酶往往催化多酶体系的第一步反应,受反应序列的终产物抑制,终产物与别构酶的调节部位相结合,由此调节多酶体系的反应速率。别构酶有协同效应,[S]对υ的动力学曲线呈S形曲线(正协同)或表现双曲线(负协同),两者均不符合米氏方程。ATCase作为别构酶的典型代表,已经测定了其三维结构,详细研究了别构机制和催化作用机制。为了解释别构酶协同效应的机制,有两种分子模型受到人们重视,即协同模型和序变模型。酶原经过蛋白水解酶专一作用释放出肽段,构象发生变化,形成酶的活性部位,变成有活性的酶,这个活化过程,是生物体的一种调控机制。可逆地共价修饰调控作用是通过共价调节酶进行的,通过其他酶对其多肽链某些基团进行可逆地共价修饰,使处于活性与非活性的互变状态,从而调节酶活性。共价修饰的基团主要是磷酸化、腺苷酰化、尿苷酰化及ADP-核糖基化等。 同工酶是指催化相同的化学反应,但其蛋白质分子结构、理化性质和免疫性能等方面不同的一组酶。同工酶是研究代谢调节、分子遗传、生物进化、个体发育、细胞分化和癌变的有力工具,在酶学、生物学及医学研究中占有重要地位。LDH同工酶研究的比较清楚,是由良种不同亚基组成的四聚体,有5种同工酶,在不同组织中含量不同,反映了同工酶的组织特异性。 习题 1.阐明酶活性部位的概念。可使用那些主要方法研究酶的活性部位? 答:酶的活性部位对于不需要辅酶的酶来说,就是指酶分子在三维结构上比较靠近的几个氨基酸残基负责与底物的结合与催化作用的部分;对于需要辅酶的灭来说,辅酶分子或辅酶分子上的某一部分结构,往往也是酶活性部位的组成部分。 研究酶活性部位的方法有:酶分子侧链基团的化学修饰法、动力学参数测定法、X射线晶体结构分析法和定点诱变法。 2.简要阐明胰Rnase A的活性部位如何确定?
第10章 酶的作用机制
第十章酶的作用机制 P384 (一)酶的活性部位 (1)酶活性部位:酶的特殊催化能力只局限在大分子的一定区域,活性部位又称活性中心。 酶分子中与酶活力直接相关的区域称为活性中心,分为: ①结合部位:负责与底物结合,决定酶的专一性。 ②催化部位:负责催化底物键的断裂或形成,决定酶的催化能力。 对需要辅酶的酶,辅酶分子或辅酶分子某一部分结构往往是酶活性部位组成部分。 (2)酶活性部位特点: 1. 活性部位只占酶分子中相当小的部分,通常1~2%。P384 表10-1列举一些酶活性部位的氨基酸残基。如溶菌酶一共129个氨基酸残基,活性部位为Asp52和Glu35;胰凝乳蛋白酶241个残基,活性部位为His57,Asp102,Ser 195。 2. 活性部位为三维实体。活性部位氨基酸残基在一级结构上可能相距甚远,甚至不在一条肽链上,但在空间结构上相互靠近。因此空间结构破坏酶即失活。活性中心以外部分可为酶活性中心提供三维结构。 3. 酶与底物的结构互补是指在酶和底物结合过程中,相互构象发生一定变化后才互补。如P385 图10-1。 4. 活性部位位于酶分子表面的一个裂缝内。裂缝中为一个疏水微环境,也含有某些极性氨基酸残基有利于催化,底物在此裂缝内有效浓度很高。 5. 酶与底物结合形成ES复合物主要靠次级键:氢键、盐键、范德华力和疏水相互作用。 6. 酶活性部位具有柔性和可运动性。 (二)研究酶活性部位的方法 (1)酶侧链基团化学修饰法: 1.特异性共价修饰:如二异丙基磷酰氟(DFP)专一地与酶活性部位Ser-OH的羟基共价结合,使酶失活。如胰凝乳蛋白酶共28个Ser,但DFP只与活性中心的Ser反应,见P386。反应后,用HCl将酶部分水解,得含二异丙基磷酸酯(DIP)基团的肽的片断,序列分析定出DIP-Ser为Ser195。 2.亲和标记:用与底物结构相似的修饰剂,对酶活性部位进行专一性共价修饰。 如TPCK(结构式见P387),结构与胰凝乳蛋白酶的底物对甲苯磺酰-L-苯丙氨酸乙酯(TPE)类似,TPCK只与胰凝乳蛋白酶中His57结合,说明His57为该酶活性部位的一个氨基酸残基。(2)X-射线晶体结构分析: 可提供酶分子三维结构,了解酶活性部位氨基酸残基所处相对位置与状态,与底物结合后酶分子在底物周围氨基酸残基排列状况,被作用键周围残基状况等。由此提出活性中心处氨基酸残基组成及催化作用形式。 如溶菌酶:对它水解的糖苷键周围氨基酸残基分析后确定酶的催化基团为Glu35和Asp52。 又如胰凝乳蛋白酶经X-射线晶体结构分析,表明活性部位由Ser195、His57和Asp102组成,并提出这三个氨基酸残基联在一起形成一个“电荷中继网”,如P409 图10-40所示:①没有底物时,His57未质子化,三联体排列为A;②在加上底物后,Ser195转移一个质子给His57,带正电荷的咪唑环通过带负电荷的Asp102静电相互作用被稳定,成为B。详细酶作用机制见P410 图10-42。 (三)影响酶催化效率的有关因素: (1)底物和酶的邻近效应与定向效应。 酶催化反应高效率一重要原因是将分子间反应变为分子内反应。 邻近效应:底物与酶先形成中间体络合物,两分子成一个分子,分子内反应速度比分子间反应速度提高。P388 图10-2 所示:用咪唑催化乙酸对硝基苯酯水解来证实:咪唑可催化乙酸对硝基苯酯的酯键水解成乙酸和对硝基苯酚;若将咪唑分子先共价连接在底物上,在分子内催化酯键
酶促反应的特点与作用机制
20 ~ 20 学年度第学期 教师课时授课教案 学科系:医学院授课教师: 专业:科目:生物化学 教研室主任签字:学科系系办主任签字:年月日年月日
第二节酶促反应的特点与作用机制 一、酶促反应的特点 酶是一类催化剂,具有一般催化剂的特征:在化学反应前后没有质和量的改变;只能催化热力学上允许进行的反应;只加速可逆反应的进程,不改变平衡点;对可逆反应的正反应和逆反应都具有催化作用。但酶的化学本质是蛋白质,又具有一般催化剂所没有的特征。 (一)高度的催化效率 酶的催化效率通常比非催化反应高108~1020倍,比一般催化剂高107-1013倍。例如蔗糖酶催化蔗糖水解的速率是H+催化作用的2.5×1012倍,脲酶催化尿素的水解速率是H+催化作用的7×1012倍,且不需要较高的反应温度。研究表明,酶能更有效地降低反应的活化能,使参与反应的活化分子数量显著增加,从而大大提高酶的催化效率。 (二)高度的专一性 一种酶只能催化一种或一类化合物,或一种化学键,发生一定的化学反应,生成一定的产物,这种特性称为酶的专一性或特异性。根据酶对底物选择的严格程度不同,酶的专一性可分为三种类型。 1.绝对专一性酶只作用于某一特定的底物,进行一种专一的反应,生成一种特定的产物,称为绝对专一性。例如,尿酶只催化尿素水解成NH3和CO2,而对尿素的衍生物如甲基尿素没有催化作用。 2.相对专一性有些酶能作用于一类化合物或一种化学键,这种不太严格的选择性称为相对专一性。如磷酸酶对一般的磷酸键都能水解,不论是甘油磷酸酯,还是葡萄糖磷酸酯;蔗糖酶不仅水解蔗糖,
也能水解棉子糖,使之生成蜜二糖和果糖。 3.立体异构专一性有些酶对底物的立体构型有要求,仅作用于底物的一种立体异构体,这种特性称为酶的立体异构专一性。如L-氨基酸氧化酶只作用于L-氨基酸,对D-氨基酸则没有催化作用;淀粉酶只能水解淀粉中的α-1,4-糖苷键,而不能水解纤维素中的β-1,4-糖苷键。 (三)酶具有不稳定性 酶所催化的反应都是在比较温和的条件下进行的,如常温、常压、接近中性的环境等。由于酶的化学本质是蛋白质,任何能引起蛋白质变性的理化因素,如强酸、强碱、重金属盐、高温、紫外线、X射线等均能影响酶的催化活性,甚至使酶完全失活。 (四)酶促反应具有可调节性 酶促反应受多种因素的调控,以适应内外环境变化和生命活动的需要。例如在细胞内酶的分布具有区域化;酶原的激活使酶在合适的环境被激活和发挥作用;代谢物对关键酶、变构酶的抑制与激活和酶的共价修饰等调节;酶的含量受到酶蛋白合成的诱导、阻遏与酶降解速率的调节。 二、酶的作用机制 (一)酶能更有效地降低反应活化能 在任何一种热力学允许的反应体系中,底物分子所含能量各不相同,只有那些能量达到或超过一定水平的过渡态分子(即活化分子)オ有可能发生化学反应,底物分子达到活化分子所需要的最小能量称为
第十章 酶的作用机制和酶活性的调节讲义
中国海洋大学海洋生命学院
生物化学讲义
2007 年
第十章
酶的作用机制和酶的调节
目的和要求:理解、掌握酶活性部位的相关概念和特点;掌握酶催化高效性的相关机理;了解 几种酶的催化机制,理解结构和功能的适应性;了解酶活性的调节方式,掌握酶活性的别构调 节、可逆共价调节和酶原激活调节方式及生物代谢中的作用。 一、酶的活性部位 ㈠ 酶的活性部位的特点 1、概念:三维结构上比较接近的少数特异的氨基酸残基参与底物的结合与催化作用,这一与酶活力直 接相关的区域称酶的活性部位。 结合部位:专一性 催化部位:催化能力,对需要辅酶的酶分子,辅酶或其一部分就是活性中心的组成部分;组成酶活 性部位的氨基酸数目对不同酶而言存在差异,占整个酶氨基酸残基小部分 酶活性部位的基团:亲核性基团,丝氨酸的羟基,半胱氨酸的巯基和组氨酸的咪唑基。酸碱性基团: 天冬氨酸和谷氨酸的羧基,赖氨酸的氨基,酪氨酸的酚羟基,组氨酸的咪唑基和半胱氨酸的巯基等。 2、特点 ⑴ 活性部位在酶分子的总体中只占相当小的部分(1%~2%) ⑵ 酶的活性部位是一个三维实体 ⑶ 酶的活性部位并不是和底物的形状互补的 ⑷ 酶的活性部位是位于酶分子表面的一个裂隙内 ⑸ 底物通过次级键结合到酶上 ⑹ 酶活性部位具有柔性 ㈡ 研究酶活性部位的方法 1、酶分子基团的侧链化学修饰 ⑴ 非特异性共价修饰:活力丧失程度与修饰剂浓度有正比关系;底物或可逆的抑制剂可保护共价修饰 剂的修饰作用。 ⑵ 特异性共价修饰:分离标记肽段,可判断活性部位的氨基酸残基,如二异丙基氟磷酸(DFP)专一 性与胰凝乳蛋白酶活性部位丝氨酸残基的羟基结合。 ⑶ 亲和标记: 利用底物类似物和酶活性部位的特殊亲和力将酶加以修饰标记来研究酶活性部位的方法。 修饰剂的特点:①结构与底物类似,能专一性引入到酶活性部位;②具活泼化学基团,能与活性部 位某一氨基酸共价结合,相应的试剂称“活性部位指示剂” 。 胰凝乳蛋白酶和胰蛋白酶,TPE 是酶的底物,TPCK 是酶的亲和试剂,当酶与 TPCK 温浴后,酶活 性丧失,这种结合具有空间结构的需求,同时也阻止其他试剂如 DFP 结合。对酶活性中心的组氨酸 咪唑环进行修饰。 ⑷ 自杀性底物标记:底物与酶结合并被催化所生成的产物可以与活性部位的基团专一性结合从而抑制 酶的催化活性。 2、动力学参数测定法:通过动力学方法求得相关参数,作出相应判断。 3、X-射线晶体衍射法:如溶菌酶和胰蛋白酶活性中心的测定
1
第十章 酶的作用机制和酶的调节
第十章酶的作用机制和酶的调节 一、是非判断题 1.某些调节酶的V-[S]的S形曲线表明,酶与少量底物的结合增加了酶对后续底物分子的亲和力。() 2.诱导酶是指当细胞加入特定诱导物后,诱导产生的酶,这种诱导物往往是该酶的产物。 () 3.酶反应的最适pH值只取决于酶蛋白本身的结构。() 答案 1.对。2.错。3.错。 二、填空题 1.胰凝乳蛋白酶的活性中心主要含有、、和基,三者构成一个氢键体系,使其中的上的成为强烈的亲核基团,此系统称为系统或。 2.与酶催化的高效率有关的因素有、、、 、等。 3.变构酶的特点是:(1),(2),它不符合一般的,当以V对[S]作图时,它表现出型曲线,而非曲线。它是酶。 4.酶的活性中心包括和两个功能部位,其中直接与底物结合,决定酶的专一性,是发生化学变化的部位,决定催化反应的性质。 5.解释别构酶作用机理的假说有模型和模型两种。 答案 1.Ser195;His57;Asp102;Ser195;氧原子;电荷转接;电荷中继网 2.邻近效应;定向效应;诱导应变;共价催化;活性中心酸碱催化 3.由多个亚基组成;除活性中心外还有变构中心;米氏方程;S;双;寡聚酶 4.结合部位;催化部位;结合部位;催化部位 5.齐变;序变 三、选择题 1.酶的活性中心是指: A.酶分子上含有必需基团的肽段B.酶分子与底物结合的部位 C.酶分子与辅酶结合的部位D.酶分子发挥催化作用的关键性结构区 E.酶分子有丝氨酸残基、二硫键存在的区域 2.酶催化作用对能量的影响在于: A.增加产物能量水平B.降低活化能C.降低反应物能量水平 D.降低反应的自由能E.增加活化能 3.酶的活化和去活化循环中,酶的磷酸化和去磷酸化位点通常在酶的哪一种氨基酸残基上:A.天冬氨酸B.脯氨酸C.赖氨酸
酶的作用机理
酶 一、酶的作用机理: 1、降低反应活化能:在任何化学反应中,反应物分子必须超过一定的能阈,成为活化的状态,才能发生变化,形成产物。这种提高低能分子达到活化状态的能量,称为活化能。催化剂的作用,主要是降低反应所需的活化能,以致相同的能量能使更多的分子活化,从而加速反应的进行。酶能显著地降低活化能,故能表现为高度的催化效率。例如H2O2酶将H2O2转变为H2O 和O2,酶能降低反应活化能,使反应速度增高千百万倍以上。 2、复合物学说:酶催化某一反应时,首先在酶的活性中心与底物结合生成酶-底物复合物,此复合物再进行分解而释放出酶,同时生成一种或数种产物,此过程可用下式表示:E+S →ES→E+P,ES的形成改变了原来反应的途径,可使底物的活化能大大降低,从而使反应加速。 3、高效率的机理: (1)趋近效应和定向效应:酶可以将它的底物结合在它的活性部位,由于化学反应速度与反应物浓度成正比,若在反应系统的某一局部区域,底物浓度增高,则反应速度也随之提高,此外,酶与底物间的靠近具有一定的取向,这样反应物分子才被作用,大大增加了ES复合物进入活化状态的机率。 (2)张力作用:底物的结合可诱导酶分子构象发生变化,比底物大得多的酶分子的三、四级结构的变化,也可对底物产生张力作用,使底物扭曲,促进ES进入活性状态。 (3)酸碱催化作用:酶的活性中心具有某些氨基酸残基的R基团,这些基团往往是良好的质子供体或受体,在水溶液中这些广义的酸性基团或广义的碱性基团对许多化学反应是有力的催化剂。 (4)共价催化作用:某些酶能与底物形成极不稳定的、共价结合的ES复合物,这些复合物比无酶存在时更容易进行化学反应。
第10章 酶的作用机制和酶的调节
第十章酶的作用机制和酶的调节 第一节酶的活性部位 一、酶活性部位的特点 只是酶分子的一小部分;通过诱导契合形成与底物互补的特定三维结构;通过次级键与底物相互作用;有一定的柔性(邹承鲁的对比研究发现活性部位的柔性比其他部位更强);包括底物结合基团和催化基团,但二者的区分有时并不严格。 二、研究酶活性部位的方法 (一)侧链基团的化学修饰法 非特异性共价修饰(作用于酶分子中某一基团),酶活力丧失与修饰剂浓度成比例,底物或竞争性抑制剂可降低修饰作用。特异性共价修饰(作用于特定酶的特定基团),如二异丙基氟磷酸(DFP)只与胰凝乳蛋白酶活性部位的丝氨酸羟基结合;亲和标记试剂可以与活性部位的特定基团共价定量结合,如对甲苯磺酰-L-苯丙氨酰氯甲基酮(TPCK)与胰凝乳蛋白酶活性部位丝氨酸羟基的结合。 (二)动力学参数测定法 (三)X-射线晶体结构分析法 (四)定点诱变法 第二节酶催化反应的独特性质
第三节影响酶催化效率的有关因素 一、底物和酶的邻近效应(approximation,proximity)与定向效应(orientation) 底物分子结合到酶的活性中心,使底物在酶活性中心的有效浓度大大增加,从而加快反应的速度。 一个有机化学模型。咪唑催化对硝基苯酯的水解,分子内反应比分子间反应快24 倍。 底物分子中参与反应的基团相互接近,并被严格定向定位,使酶促反应具有高效率称定向效应。邻羟基苯丙酸内脂的形成反应,两个甲基使羧基和羟基更好的定向,使反应速率提高2.5×1011。 二、底物的形变(distortion)和诱导契合(inducednt) 脯氨酸消旋酶的过渡态类似物与酶的亲和力远大于其底物,说明酶可以引起底物的形变。
酶的作用机制
第10章、酶的作用机制和酶的调节(P384) 本章重点:1、酶的活性中心,2、影响酶活性的条件和因素,3、酶和底物之间的相互作用和关系,4、酶催化反应机制,5酶概念的发展-核酶本章的主要研究内容: (一)酶的活性部位 (1)酶活性部位:酶的特殊催化能力只局限在大分子的一定区域,活性部位又称活性中心。 酶分子中与酶活力直接相关的区域称为活性中心,分为: ①结合部位:负责与底物结合,决定酶的专一性。 ②催化部位:负责催化底物键的断裂或形成,决定酶的催化能力。 对需要辅酶的酶,辅酶分子或辅酶分子某一部分结构往往是酶活性部位组成部分。 (2)酶活性部位特点: 1. 活性部位只占酶分子中相当小的部分,通常1~2%。P384 表10-1列举一些酶活性部位的氨基酸残基。如溶菌酶一共129个氨基酸残基,活性部位为Asp52和Glu35;胰凝乳蛋白酶241个残基,活性部位为His57,Asp102,Ser 。 195 2. 活性部位为三维实体。活性部位氨基酸残基在一级结构上可能相距甚远,甚至不在一条肽链上,但在空间结构上相互靠近。因此空间结构破坏酶即失活。活性中心以外部分可为酶活性中心提供三维结构。 3. 酶与底物的结构互补是指在酶和底物结合过程中,相互构象发生一定变化后才互补。如P385 图10-1。 4. 活性部位位于酶分子表面的一个裂缝内。裂缝中为一个疏水微环境,也含有某些极性氨基酸残基有利于催化,底物在此裂缝内有效浓度很高。 5. 酶与底物结合形成ES复合物主要靠次级键:氢键、盐键、范德华力和疏水相互作用。 6. 酶活性部位具有柔性和可运动性。 (二)研究酶活性部位的方法 (1)酶侧链基团化学修饰法: 1.特异性共价修饰:如二异丙基磷酰氟(DFP)专一地与酶活性部位Ser-OH的羟基共价结合,使酶失活。如胰凝乳蛋白酶共28个Ser,但DFP 只与活性中心的Ser反应,见P386。反应后,用HCl将酶部分水解,得含二异丙基磷酸酯(DIP)基团的肽的片断,序列分析定出DIP-Ser为Ser195。 2.亲和标记:用与底物结构相似的修饰剂,对酶活性部位进行专一性共价修饰。 如TPCK(结构式见P387),结构与胰凝乳蛋白酶的底物对甲苯磺酰-L-苯丙氨酸乙酯(TPE)类似,TPCK只与胰凝乳蛋白酶中His57结合,说明His57为该酶活性部位的一个氨基酸残基。 (2)X-射线晶体结构分析: 可提供酶分子三维结构,了解酶活性部位氨基酸残基所处相对位置与状态,与底物结合后酶分子在底物周围氨基酸残基排列状况,被作用键周围残
生物化学-生化知识点_酶的作用机制 (10章)
2.11 酶的作用机制 P384 10章 ①①①酶的活性部位 (1)酶活性部位:酶的特殊催化能力只局限在大分子的一定区域,活性部位又称活性中心。 酶分子中与酶活力直接相关的区域称为活性中心,分为: ①结合部位:负责与底物结合,决定酶的专一性。 ①催化部位:负责催化底物键的断裂或形成,决定酶的催化能力。 对需要辅酶的酶,辅酶分子或辅酶分子某一部分结构往往是酶活性部位组成部分。 (2)酶活性部位特点: 1. 活性部位只占酶分子中相当小的部分,通常1~2%。P384 表10- 1列举一些酶活性部位的氨基酸残基。如溶菌酶一共129个氨基酸残基,活性部位为Asp52和Glu35;胰凝乳蛋白酶241个残基,活性部位为His57,Asp102,Ser 。 195 2. 活性部位为三维实体。活性部位氨基酸残基在一级结构上可能相距甚远,甚至不在一条肽链上,但在空间结构上相互靠近。因此空间结构破坏酶即失活。 活性中心以外部分可为酶活性中心提供三维结构。 3. 酶与底物的结构互补是指在酶和底物结合过程中,相互构象发生一定变化后才互补。如P385 图10-1。 4. 活性部位位于酶分子表面的一个裂缝内。裂缝中为一个疏水微环境,也含有某些极性氨基酸残基有利于催化,底物在此裂缝内有效浓度很高。 5. 酶与底物结合形成ES复合物主要靠次级键:氢键、盐键、范德华力和疏水相互作用。 6. 酶活性部位具有柔性和可运动性。 ①①①研究酶活性部位的方法 ①1①酶侧链基团化学修饰法: 1.特异性共价修饰:如二异丙基磷酰氟(DFP)专一地与酶活性部位 Ser- OH的羟基共价结合,使酶失活。如胰凝乳蛋白酶共28个Ser,但DFP只与活性中心的Ser反应,见P386。反应后,用HCl将酶部分水解,得含二异丙基磷酸酯(DIP)基团的肽的片断,序列分析定出DIP-Ser为Ser195。 2.亲和标记:用与底物结构相似的修饰剂,对酶活性部位进行专一 性共价修饰。 如TPCK(结构式见P387),结构与胰凝乳蛋白酶的底物对甲苯磺酰-L-苯丙氨酸乙酯(TPE)类似,TPCK只与胰凝乳蛋白酶中His57结合,说明His57为该酶活性部位的一个氨基酸残基。 (2) X-射线晶体结构分析: 可提供酶分子三维结构,了解酶活性部位氨基酸残基所处相对位置与状态,