几种破乳方法的比较
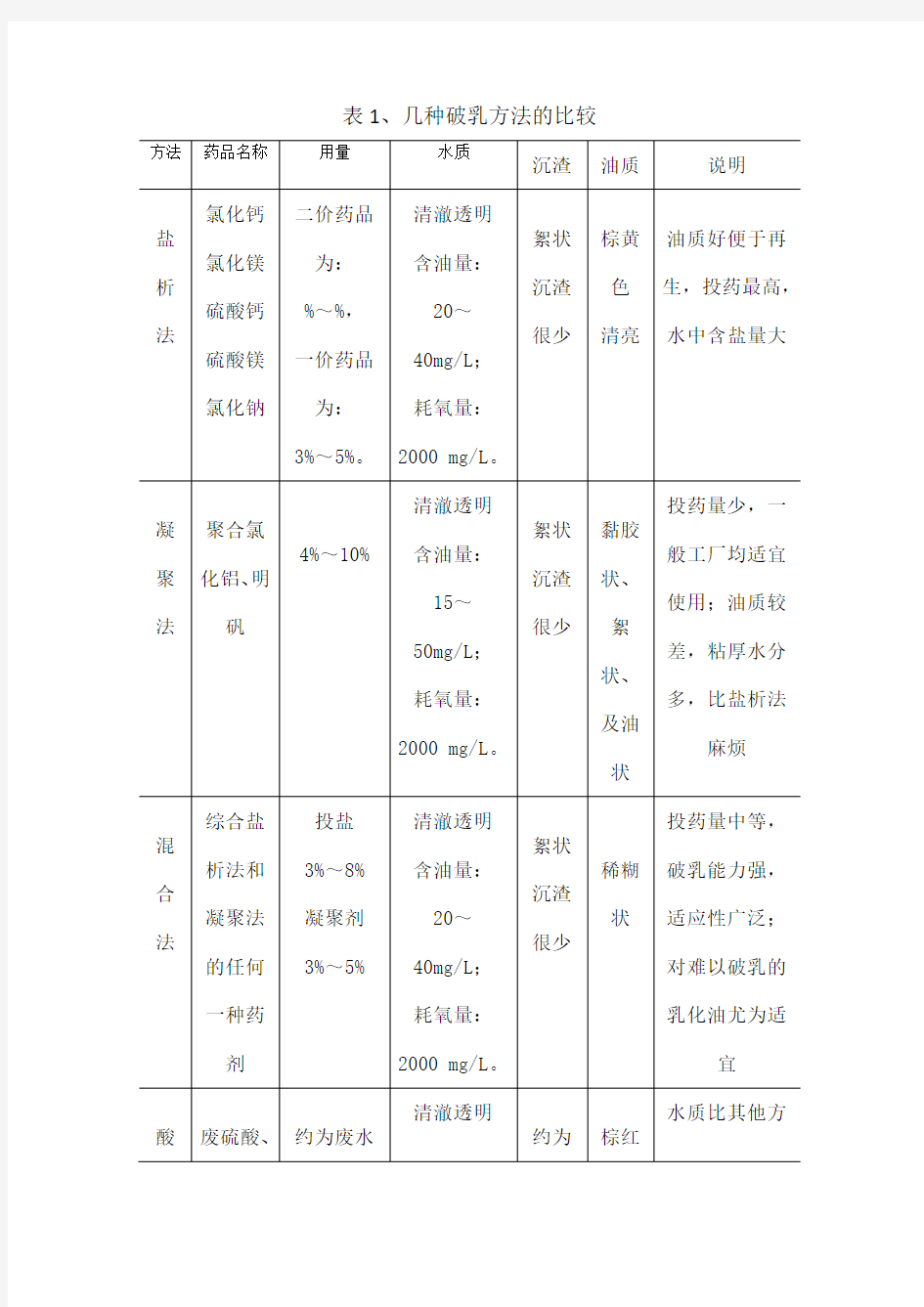

表1、几种破乳方法的比较
各种排序算法比较
排序算法 一、插入排序(Insertion Sort) 1. 基本思想: 每次将一个待排序的数据元素,插入到前面已经排好序的数列中的适当位置,使数列依然有序;直到待排序数据元素全部插入完为止。 2. 排序过程: 【示例】: [初始关键字] [49] 38 65 97 76 13 27 49 J=2(38) [38 49] 65 97 76 13 27 49 J=3(65) [38 49 65] 97 76 13 27 49 J=4(97) [38 49 65 97] 76 13 27 49 J=5(76) [38 49 65 76 97] 13 27 49 J=6(13) [13 38 49 65 76 97] 27 49 J=7(27) [13 27 38 49 65 76 97] 49 J=8(49) [13 27 38 49 49 65 76 97] Procedure InsertSort(Var R : FileType); //对R[1..N]按递增序进行插入排序, R[0]是监视哨// Begin for I := 2 To N Do //依次插入R[2],...,R[n]// begin R[0] := R[I]; J := I - 1; While R[0] < R[J] Do //查找R[I]的插入位置// begin R[J+1] := R[J]; //将大于R[I]的元素后移// J := J - 1 end R[J + 1] := R[0] ; //插入R[I] // end End; //InsertSort // 二、选择排序 1. 基本思想: 每一趟从待排序的数据元素中选出最小(或最大)的一个元素,顺序放在已排好序的数列的最后,直到全部待排序的数据元素排完。 2. 排序过程: 【示例】: 初始关键字[49 38 65 97 76 13 27 49] 第一趟排序后13 [38 65 97 76 49 27 49] 第二趟排序后13 27 [65 97 76 49 38 49] 第三趟排序后13 27 38 [97 76 49 65 49] 第四趟排序后13 27 38 49 [49 97 65 76] 第五趟排序后13 27 38 49 49 [97 97 76]
几种育种方法的比较
育种的方法和应用 生物育种是一门很复杂的技术,针对不同的生物应采用不同的育种方式,要对各种育种方式进行比较,选择简易、可操作的方式。同一种育种方式应用于不同的生物也会有不尽相同的育种过程,所以我们无论在生产实践中还是有关习题训练中都应灵活应用。 一、几种育种的方法的比较 在高中阶段所介绍的育种方法主要有:诱变育种、杂交育种、多倍体育种、单倍体育种、细胞工程育种(组织培养育种)、基因工程育种(转基因育种)、植物激素育种等。 1、杂交育种 (1)原理:基因重组。 (2)方法:连续自交,不断选种。(不同个体间杂交产生后代,然后连续自交,筛选所需纯合子) (3)发生时期:有性生殖的减数分裂第一次分裂后期或四分体时期, (4)优点:使同种生物的不同优良性状集中于同一个个体,具有预见性。’ (5)缺点:育种年限长,需连续自交才能选育出需要的优良性状。 (6)举例:矮茎抗锈病小麦等。 2、诱变育种 (1)原理:基因突变。 (2)方法:用物理因素(如x射线、1射线等)、化学因素(如亚硝酸、秋水仙素等各种化学药剂)、生物因素或空间诱变育种(用宇宙强辐射、微重力等条件)来处理生物。 (3)发生时期:有丝分裂间期或减数分裂第一次分裂间期(DNA分子复制的时候)。 (4)优点:能提高变异频率,加速育种进程,可大幅度改良某些性状,创造人类需要的变异类型,从中选择培育出优良的生物品种;变异范围广。 (5)缺点:有利变异少,须大量处理材料;诱变的方向和性质不能控制;改良数量性状效果较差,具有盲目性。 (6)举例:青霉素高产菌株、太空椒、高产小麦、“彩色小麦”等。 3、多倍体育种 (1)原理:染色体变异。 (2)方法:秋水仙素处理萌发的种子或幼苗(秋水仙素能抑制细胞有丝分裂过程中纺锤体的形成)。 (3)优点:可培育出自然界中没有的新品种,且培育出的植物器官大,产量高,营养丰富。 (4)缺点:结实率低,发育延迟。 (5)举例:三倍体无子西瓜、八倍体小黑麦。 4、单倍体育种 (1)原理:染色体变异。 (2)方法:花药离体培养获得单倍体植株,再用秋水仙素等诱导剂人工诱导染色体数目加倍。 (3)优点:自交后代不发生性状分离,能明显缩短育种年限,加速育种进程。 (4)缺点:技术相当复杂,需与杂交育种结合,其中的花药离体培养过程需要组织培养技术手段的支持,多限于植物。 (5)举例:“京花一号”小麦。 5、细胞工程育种 (1)方式:植物组织培养植物体细胞杂交细胞核移植 (2)原理:植物细胞的全能性植物细胞膜的流动性动物细胞核的全能性 (3)方法:离体的植物器官、组织或细胞→愈伤组织→根、芽→植物体去掉细胞壁
转染步骤及经验(精华)
转染步骤及经验(精华) 一、基础理论 转染是将外源性基因导入细胞内的一种专门技术。分类:物理介导方法:电穿孔法、显微注射和基因枪;化学介导方法:如经典的磷酸钙共沉淀法、脂质体转染方法、和多种阳离子物质介导的技术;生物介导方法:有较为原始的原生质体转染,和现在比较多见的各种病毒介导的转染技术。理想细胞转染方法,应该具有转染效率高、细胞毒性小等优点。病毒介导的转染技术,是目前转染效率最高的方法,同时具有细胞毒性很低的优势。但是,病毒转染方法的准备程序复杂,常常对细胞类型有很强的选择性,在一般实验室中很难普及。其它物理和化学介导的转染方法,则各有其特点。需要指出的一点,无论采用哪种转染技术,要获得最优的转染结果,可能都需要对转染条件进行优化。影响转染效率的因素很多,从细胞类型、细胞培养条件和细胞生长状态到转染方法的操作细节(见后文)。 二、转染操作流程(以常用的6孔板为例) (1) 细胞培养: 取6孔培养板,以3x104/cm2密度铺板,37℃5%CO2培养箱中培养至70%~90%汇合。(不同细胞略有不同,根据实验室优化的条件进行,汇合过分,转染后不利筛选细胞)。 (2) 转染液制备: 在EP管中制备以下两液(为转染每一个孔细胞所用的量) A液:用不含血清培养基稀释1-10μg DNA,终量100μL, B液:用不含血清培养基稀释对应量的转染试剂,终量100μL; 轻轻混合A、B液(1:1混匀),室温中置15分钟,稍后会出现微浊现象,但并不妨碍转染。 (3) 转染准备:用2mL不含血清培养液漂洗两次,再加入2mL不含血清及PS的培养液。 (4) 转染:把A/B复合物缓缓加入培养液中(缓慢滴加),轻轻摇匀,37℃温箱置6~8小时,吸除无血清转染液,换入正常培养液继续培养。 三、转染注意事项 1. 血清 A. DNA-阳离子脂质体复合物形成时不能含血清,因为血清会影响复合物的形成。 B.一般细胞对无血清培养可以耐受几个小时没问题,转染用的培养液可以含血清也可以不加,但血清一度曾被认为会降低转染效率,转染培养基中加入血清需要对条件进行优化。 C. 对于对血清缺乏比较敏感的细胞,可以使用一种营养丰富的无血清培养基OPTI-MEMⅠ培养基, 或者在转染培养基中使用血清。对血清缺乏比较敏感的贴壁细胞,建议使用LIPOFECTAMINE 2000。无血清培养基OPTI-MEM(GIBICO)很好用,有条件的话,就用它代替PBS洗细胞两遍,注意洗的时候要轻,靠边缘缓缓加入液体,然后不要吹吸细胞,而是转动培养板让液体滚动在细胞表面。如果洗的太厉害,细胞又损失一部分,加了脂质体后,细胞受影响就更大了,死亡细胞会增多。 2.抗生素(PS) 抗生素,比如青霉素和链霉素,是影响转染的培养基添加物。这些抗生素一般对于真核细胞无毒,但阳离子脂质体试剂增加了细胞的通透性,使抗生素可以进入细胞。这降低了细胞的活性,导致转染效率低。所以,在转染培养基中不能使用抗生素,甚至在准备转染前进行细胞铺板时也要避免使用抗生素。这样,在转染前也不必润洗细胞。对于稳定转染,不要在选择性培养基中使用青霉素和链霉素,因为这些抗生素是GENETICIN选择性抗生素的竞争性抑制剂。另外,为了保证无血
各种排序算法的总结和比较
各种排序算法的总结和比较 1 快速排序(QuickSort) 快速排序是一个就地排序,分而治之,大规模递归的算法。从本质上来说,它是归并排序的就地版本。快速排序可以由下面四步组成。 (1)如果不多于1个数据,直接返回。 (2)一般选择序列最左边的值作为支点数据。(3)将序列分成2部分,一部分都大于支点数据,另外一部分都小于支点数据。 (4)对两边利用递归排序数列。 快速排序比大部分排序算法都要快。尽管我们可以在某些特殊的情况下写出比快速排序快的算法,但是就通常情况而言,没有比它更快的了。快速排序是递归的,对于内存非常有限的机器来说,它不是一个好的选择。 2 归并排序(MergeSort)
归并排序先分解要排序的序列,从1分成2,2分成4,依次分解,当分解到只有1个一组的时候,就可以排序这些分组,然后依次合并回原来的序列中,这样就可以排序所有数据。合并排序比堆排序稍微快一点,但是需要比堆排序多一倍的内存空间,因为它需要一个额外的数组。 3 堆排序(HeapSort) 堆排序适合于数据量非常大的场合(百万数据)。 堆排序不需要大量的递归或者多维的暂存数组。这对于数据量非常巨大的序列是合适的。比如超过数百万条记录,因为快速排序,归并排序都使用递归来设计算法,在数据量非常大的时候,可能会发生堆栈溢出错误。 堆排序会将所有的数据建成一个堆,最大的数据在堆顶,然后将堆顶数据和序列的最后一个数据交换。接下来再次重建堆,交换数据,依次下去,就可以排序所有的数据。
Shell排序通过将数据分成不同的组,先对每一组进行排序,然后再对所有的元素进行一次插入排序,以减少数据交换和移动的次数。平均效率是O(nlogn)。其中分组的合理性会对算法产生重要的影响。现在多用D.E.Knuth的分组方法。 Shell排序比冒泡排序快5倍,比插入排序大致快2倍。Shell排序比起QuickSort,MergeSort,HeapSort慢很多。但是它相对比较简单,它适合于数据量在5000以下并且速度并不是特别重要的场合。它对于数据量较小的数列重复排序是非常好的。 5 插入排序(InsertSort) 插入排序通过把序列中的值插入一个已经排序好的序列中,直到该序列的结束。插入排序是对冒泡排序的改进。它比冒泡排序快2倍。一般不用在数据大于1000的场合下使用插入排序,或者重复排序超过200数据项的序列。
几种数学计算方法的比较
有限元法,有限差分法和有限体积法的区别 有限差分方法(FDM)是计算机数值模拟最早采用的方法,至今仍被广泛运用。该方法将求解域划分为差分网格,用有限个网格节点代替连续的求解域。有限差分法以Taylor级数展开等方法,把控制方程中的导数用网格节点上的函数值的差商代替进行离散,从而建立以网格节点上的值为未知数的代数方程组。该方法是一种直接将微分问题变为代数问题的近似数值解法,数学概念直观,表达简单,是发展较早且比较成熟的数值方法。对于有限差分格式,从格式的精度来划分,有一阶格式、二阶格式和高阶格式。从差分的空间形式来考虑,可分为中心格式和逆风格式。考虑时间因子的影响,差分格式还可以分为显格式、隐格式、显隐交替格式等。目前常见的差分格式,主要是上述几种形式的组合,不同的组合构成不同的差分格式。差分方法主要适用于有结构网格,网格的步长一般根据实际地形的情况和柯朗稳定条件来决定。 构造差分的方法有多种形式,目前主要采用的是泰勒级数展开方法。其基本的差分表达式主要有三种形式:一阶向前差分、一阶向后差分、一阶中心差分和二阶中心差分等,其中前两种格式为一阶计算精度,后两种格式为二阶计算精度。通过对时间和空间这几种不同差分格式的组合,可以组合成不同的差分计算格式。 有限元方法的基础是变分原理和加权余量法,其基本求解思想是把计算域划分为有限个互不重叠的单元,在每个单元内,选择一些合适的节点作为求解函数的插值点,将微分方程中的变量改写成由各变量或其导数的节点值与所选用的插值函数组成的线性表达式,借助于变分原理或加权余量法,将微分方程离散求解。采用不同的权函数和插值函数形式,便构成不同的有限元方法。有限元方法最早应用于结构力学,后来随着计算机的发展慢慢用于流体力学的数值模拟。在有限元方法中,把计算域离散剖分为有限个互不重叠且相互连接的单元,在每个单元内选择基函数,用单元基函数的线形组合来逼近单元中的真解,整个计算域上总体的基函数可以看为由每个单元基函数组成的,则整个计算域内的解可以看作是由所有单元上的近似解构成。在河道数值模拟中,常见的有限元计算方法是由变分法和加权余量法发展而来的里兹法和伽辽金法、最小二乘法等。根据所采用的权函数和插值函数的不同,有限元方法也分为多种计算格式。从权函数的选择来说,有配置法、矩量法、最小二乘法和伽辽金法,从计算单元网格的形状来划分,有三角形网格、四边形网格和多边形网格,从插值函数的精度来划分,又分为线性插值函数和高次插值函数等。不同的组合同样构成不同的有限元计算格式。对于权函数,伽辽金(Galerkin)法是将权函数取为逼近函数中的基函数;最小二乘法是令权函数等于余量本身,而内积的极小值则为对代求系数的平方误差最小;在配置法中,先在计算域内选取N个配置点。令近似解在选定的N个配置点上严格满足微分方程,即在配置点上令方程余量为0。插值函数一般由不同次幂的多项式组成,但也有采用三角函数或指数函数组成的乘积表示,但最常用的多项式插值函数。有限元插值函数分为两大类,一类只要求插值多项式本身在插值点取已知值,称为拉格朗日(Lagrange)多项式插值;另一种不仅要求插值多项式本身,还要求它的导数值在插值点取已知值,称为哈密特(Hermite)多项式插值。单元坐标有笛卡尔直角坐标系和无因次自然坐标,有对称和不对称等。常采用的无因次坐标是一种局部坐标系,它的定义取决于单元的几何形状,一维看作长度比,二维看作面积比,三维看作体积比。在二维有限元中,三角形单元应用的最早,近来四边形等参元的应用也越来越广。对于二维三角形和四边形电源单元,常采用的插值函数为有Lagrange插值直角坐标系中的线性插值函数及二阶或更高阶插值函数、面积坐标系中的线性插值函数、二阶或更高阶插值函数等。 对于有限元方法,其基本思路和解题步骤可归纳为 (1)建立积分方程,根据变分原理或方程余量与权函数正交化原理,建立与微分方程初边值
五种排序算法的分析与比较
五种排序算法的分析与比较 广东医学院医学信息专业郭慧玲 摘要:排序算法是计算机程序设计广泛使用的解决问题的方法,研究排序算法具有重要的理论意义和广泛的应用价值。文章通过描述冒泡、选择、插入、归并和快速5种排序算法,总结了它们的时间复杂度、空间复杂度和稳定性。通过实验验证了5种排序算法在随机、正序和逆序3种情况下的性能,指出排序算法的适用原则,以供在不同条件下选择适合的排序算法借鉴。 关键词:冒泡排序;选择排序;插入排序;归并排序;快速排序。 排序是计算机科学中基本的研究课题之一,其目的是方便记录的查找、插入和删除。随着计算机的发展与应用领域的越来越广,基于计算机硬件的速度和存储空间的有限性,如何提高计算机速度并节省存储空间一直成为软件设计人员的努力方向。其中,排序算法已成为程序设计人员考虑的因素之一[1],排序算法选择得当与否直接影响程序的执行效率和内外存储空间的占用量,甚至影响整个软件的综合性能。排序操作[2,3],就是将一组数据记录的任意序列,重新排列成一个按关键字有序的序列。而所谓排序的稳定性[4]是指如果在排序的序列中,存在前后相同的两个元素,排序前和排序后他们的相对位臵不发生变化。 1 算法与特性 1.1冒泡排序 1.1.1冒泡排序的基本思想
冒泡排序的基本思想是[5,6]:首先将第1个记录的关键字和第2个记录的关键字进行比较,若为逆序,则将2个记录交换,然后比较第2个和第3个记录的关键字,依次类推,直至n-1个记录和第n个记录的关键字进行过比较为止。然后再按照上述过程进行下一次排序,直至整个序列有序为止。 1.1.2冒泡排序的特性 容易判断冒泡排序是稳定的。可以分析出它的效率,在最好情况下,只需通过n-1次比较,不需要移动关键字,即时间复杂度为O(n)(即正序);在最坏情况下是初始序列为逆序,则需要进行n-1次排序,需进行n(n-1)/2次比较,因此在最坏情况下时间复杂度为O(n2),附加存储空间为O(1)。 1.2选择排序 1.2.1选择排序的基本思想 选择排序的基本思想是[5,6]:每一次从待排序的记录中选出关键字最小的记录,顺序放在已排好序的文件的最后,直到全部记录排序完毕.常用的选择排序方法有直接选择排序和堆排序,考虑到简单和易理解,这里讨论直接选择排序。直接选择排序的基本思想是n个记录的文件的直接排序可经过n-1次直接选择排序得到有序结果。 1.2.2选择排序的特性 容易得出选择排序是不稳定的。在直接选择排序过程中所需进行记录移动的操作次数最少为0,最大值为3(n-1)。然而,无论记录的初始排序如何,所需进行的关键字间的比较次数相同,均为n(n-1)/2,时间
废乳化油的破乳方法
废乳化油的破乳方法,主要有酸化法和聚化法两种。 酸化法就是往废乳化液中加入酸(如盐酸或硫酸)。 所加入的酸可利用工业废酸。 由于在目前的乳化液配方中,多数选用阴离子型乳化剂(如石油磺酸钠、磺化蓖麻油),所以遇到酸就会破坏,乳化生成相应的有机酸,使油水分离,而酸中氢离子的引入,也有助于破乳的过程。 酸的用量是待处理乳化液重量的0.2%,浓度为37%; 如果采用废酸时,则酸的用量应适当加大。 聚化法就是在废乳化油中添加盐类电解质(如0.4%氯化钙)和凝聚剂(如0.2%明矾),以达到乳化液破乳的目的。酸化法的优点是油质较好,成本低廉,水质也好,水质中含油量一般在20mg/L以下,化学耗氧量(COD)值也比其它破乳方法低;其缺点是沉渣较多。聚化法的优点是投药量少,一般工厂均有条件使用,但油质较差。 针对难处理乳化油破乳过程中存在的问题,通过对现有油水分离技术的总结和各种破乳方案的比较,提出了微波破乳—离心分离的新工艺。该工艺处理沉降罐中间层难处理乳化油技术指标优越,可有效解决该部分液压支架乳化油的破乳问题。 通过对现有离心机特点的分析,提出了适用于油、水、渣分离的BKD-1000三相立式离心机的设计方案,该机具有分离区整体旋转的特点,流体获得了较高的离心加速度。 微波破乳器的试验室模拟试验表明,采用微波破乳—离心分离工艺处理模拟乳化油,可使模拟乳化油油水有效分离,油中含水率由50.0%降至5.51%, 油的回收率达到98.33%。BKD-1000三相立式离心机的工业试验表明, 处理油田干化池含油污水可使油中含水率降至3.56%,油的回收率达到85.26%,排渣浓度达到62.18%,达到了现场提出的工业试验要求。
几种常见内部排序算法比较
常见内部排序算法比较 排序算法是数据结构学科经典的内容,其中内部排序现有的算法有很多种,究竟各有什么特点呢?本文力图设计实现常用内部排序算法并进行比较。分别为起泡排序,直接插入排序,简单选择排序,快速排序,堆排序,针对关键字的比较次数和移动次数进行测试比较。 问题分析和总体设计 ADT OrderableList { 数据对象:D={ai| ai∈IntegerSet,i=1,2,…,n,n≥0} 数据关系:R1={〈ai-1,ai〉|ai-1, ai∈D, i=1,2,…,n} 基本操作: InitList(n) 操作结果:构造一个长度为n,元素值依次为1,2,…,n的有序表。Randomizel(d,isInverseOrser) 操作结果:随机打乱 BubbleSort( ) 操作结果:进行起泡排序 InserSort( ) 操作结果:进行插入排序 SelectSort( ) 操作结果:进行选择排序 QuickSort( ) 操作结果:进行快速排序 HeapSort( ) 操作结果:进行堆排序 ListTraverse(visit( )) 操作结果:依次对L种的每个元素调用函数visit( ) }ADT OrderableList 待排序表的元素的关键字为整数.用正序,逆序和不同乱序程度的不同数据做测试比较,对关键字的比较次数和移动次数(关键字交换计为3次移动)进行测试比较.要求显示提示信息,用户由键盘输入待排序表的表长(100-1000)和不同测试数据的组数(8-18).每次测试完毕,要求列表现是比较结果. 要求对结果进行分析.
详细设计 1、起泡排序 算法:核心思想是扫描数据清单,寻找出现乱序的两个相邻的项目。当找到这两个项目后,交换项目的位置然后继续扫描。重复上面的操作直到所有的项目都按顺序排好。 bubblesort(struct rec r[],int n) { int i,j; struct rec w; unsigned long int compare=0,move=0; for(i=1;i<=n-1;i++) for(j=n;j>=i+1;j--) { if(r[j].key 数据结构各种排序方法的综合比较 结论: 排序方法平均时间最坏时间辅助存储 简单排序O(n2) O(n2) O(1) 快速排序O(nlogn)O(n2)O(logn) 堆排序O(nlogn)O(nlogn)O(1) 归并排序O(nlogn)O(nlogn)O(n) 基数排序O(d(n+rd))O(d(n+rd))O(rd) PS:直接插入排序、冒泡排序为简单排序,希尔排序、堆排序、快速排序为不稳定排序 一、时间性能 按平均的时间性能来分,有三类排序方法: 时间复杂度为O(nlogn)的方法有:快速排序、堆排序和归并排序,其中以快速排序为最好;时间复杂度为O(n2)的有:直接插入排序、起泡排序和简单选择排序,其中以直接插入为 最好,特别是对那些对关键字近似有序的记录序列尤为如此; 时间复杂度为O(n)的排序方法只有,基数排序。 当待排记录序列按关键字顺序有序时,直接插入排序和起泡排序能达到O(n)的时间复杂度;而对于快速排序而言,这是最不好的情况,此时的时间性能蜕化为O(n2),因此是应该尽量避免的情况。简单选择排序、堆排序和归并排序的时间性能不随记录序列中关键字的分布而改变。 二、空间性能 指的是排序过程中所需的辅助空间大小。 1. 所有的简单排序方法(包括:直接插入、起泡和简单选择)和堆排序的空间复杂度为O(1); 2. 快速排序为O(logn),为栈所需的辅助空间; 3. 归并排序所需辅助空间最多,其空间复杂度为O(n ); 4.链式基数排序需附设队列首尾指针,则空间复杂度为O(rd)。 三、排序方法的稳定性能 1. 稳定的排序方法指的是,对于两个关键字相等的记录,它们在序列中的相对位置,在排序之前和经过排序之后,没有改变。 2. 当对多关键字的记录序列进行LSD方法排序时,必须采用稳定的排序方法。 3. 对于不稳定的排序方法,只要能举出一个实例说明即可。 4. 快速排序和堆排序是不稳定的排序方法 各种转染试剂的中文转染方法 FuGENE6(Roche)转染步骤: 转染前一天将细胞分至培养板,转染当天细胞应50-80%融合。将细胞以1-3×105/2 ml接种于6孔板后孵育过夜将达到如此密度。 将FuGENE6 Reagent在室温孵育10-15分钟。使用之前将FuGENE6颠倒混匀一下。 1. 在PCR管中加入不含血清和双抗的营养液以稀释FuGENE6,直至总体积到100 ul。 2. 将3-6 ul FuGENE6 Reagent直接加入营养液,轻弹管壁混合。 3. 加入1-2 ug的DNA溶液(0.02-2.0 ug/ul),轻弹管壁混合。 4. 室温孵育20分钟。 5. 将6孔板中的旧营养液吸出,加入约1 ml不含血清和双抗的营养液洗涤一次,再加入2 ml不含血清和双抗的营养液。 6. 将转染复合物加入细胞,混匀使之均匀分布。 7. 3-8小时后,加入血清或换成含血清的营养液。 Lipofectamine 2000(Invitrogen)转染试剂转染步骤(6孔板): 1. 转染前一天,胰酶消化细胞并计数,细胞铺板,使其在转染日密度为90-95%。细胞铺板在2 ml含血清,不含抗生素的正常生长的培养基中。 2. 对于每孔细胞,使用250 ul无血清培养基(如OPTI-MEM I培养基)稀释4.0 ugDNA,轻轻混匀。 3. 使用前将Lipofectamine 2000转染试剂轻轻混匀,用250 ul无血清培养基(如OPTI-MEM I培养基)稀释10 ul Lipofectamine 2000转染试剂,轻轻混匀。Lipofectamine 2000稀释后,在5分钟内同稀释的DNA混合(<30分钟)。NOTE:若使用DMEM培养基,则需在5分钟内同稀释的DNA混合。 4. 混合稀释的DNA(第二步)和稀释的Lipofectamine 2000(第三步)。室温放置20分钟。 5. (optional)将6孔板中的旧营养液吸出,用无血清培养基清洗两次。加入2 ml无血清配养基。 6. 直接将复合物加入到每孔中,摇动培养板,轻轻混匀。 中保温24-48小时。无需去掉复合物或更换培养基。 7. 在37℃,5%CO 2 或者在4-5小时后更换培养生长基也不会降低转染活性。 8. 在细胞中加入复合物24-72小时后,分析细胞抽提物或进行原位细胞染色,检测报告基因活性。这依赖于细胞类型和启动子活性。对稳定表达,在开始转染一天后将细胞传代至新鲜培养基中,两天后加入筛选抗生素。进行稳定表达需要数天或数周。 贴壁细胞的稳定转染: 转染后24小时,将细胞以≥1:10的比例传代至新鲜培养基中,次日加入选择性培养基。 Lipofectamine 2000转染试剂转染步骤(24孔板): 破乳的常用方法 液-液萃取中非常重要的操作是急速地振动样品。此步骤可确保两相的完全接触,有助于质量传递。在分液漏斗发生完全的混合,产生大量的界面区域使得有效的分配出现。由于物质剧烈的振动,在液-液萃取中乳化现象经常发生,特别是那些含有表面活性剂和脂肪的样品。收集欲测物质必须先进行破乳。为 ,改变溶剂或了防止乳化形成,应用采取加热或加盐的方法破乳。通过改变K D 化学平衡作用的添加剂,诸如使用缓冲剂调节pH,盐调节离子强度等。用于破乳的常用技术如下: ①加盐; ②使用加热-冷却萃取容器; ③通过玻璃棉塞过滤乳化液样品; ④通过相过滤纸过滤乳化液样品; ⑤通过离心作用; ⑥加进少量的不同的有机溶剂。 在液-液萃取过程中,有机相、水相、乳化物和外力是乳化形成的主要因素,如果破坏乳化形成的条件就可以防止和避免乳化的形成。诸如,在脏器、血液等生物样品的萃取前,在研钵中先加入等量的无水硫酸钠与样品同时研磨,直至干沙状后,经有机溶剂萃取就不会发生乳化现象,而且可获得较高的萃取效率。但本法不适用溶液萃取。在水溶液样品中加入氯化钠使之饱和,再用有机溶剂萃取可有效地防止因为有机相与水相比重接近易引起的乳化现象。在生物体试样中含有蛋白、油脂等乳化物,它们具有降低有机相和水相界面张力的功能,将有机相液珠与水相粘合在一起,形成相对稳定的乳状液。如果除去这些乳化物就能避免乳化的形成。除去乳化物的方法很多,应当根据萃取的目的决定。例如,在萃取生物试样中不挥发性有机物时,常用的方法有:酸性乙醇浸取法、三氯乙酸沉淀蛋白法、冷冻除油脂法等均可除掉样品中的蛋白、脂肪等乳化物。此外,提高两相的体积比,一般地保持两相体积比为1:(5~10)时,可有效地防止乳化。在剧烈振摇时发生乳化,采用缓慢振摇可防止乳化。 在液-液萃取过程中发生乳化现象时,可根据乳化的程度采用适当的方法消除乳化。 如果样品出现高度乳化(即全部乳化),可采用离心法破乳。破乳率随离心转数的增加而增大,也随作用时间的延长而增大。通常采用2000r/min,作用 2min后的破乳率可达100%。但离心法不适用微乳液的破乳。也可以采用无水硫酸钠研磨法破乳,将乳浊液转入研钵中,使用无水硫酸钠研磨至沙状后再进行萃取可消除乳化现象。还可以采用蒸干法,将乳浊液置入蒸发皿中,于100℃沸水浴蒸干后,再用有机溶剂萃取。但本法不适用挥发性物质的萃取。 几种常用的育种方法比较(总结整理) 一、诱变育种: 诱变育种是指利用人工诱变的方法获得生物新品种的育种方法 原理:基因突变 方法:辐射诱变,激光、化学物质诱变,太空(辐射、失重)诱发变异→选择育成新品种 优点:能提高变异频率,加速育种过程,可大幅度改良某些性状;变异范围广。 缺点:有利变异少,须大量处理材料;诱变的方向和性质不能控制。改良数量性状效果较差。 二、杂交育种: 杂交育种是指利用具有不同基因组成的同种(或不同种)生物个体进行杂交,获得所需要的表现型类型的育种方法。其原理是基因重组。 方法:杂交→自交→选优 优点:能根据人的预见把位于两个生物体上的优良性状集于一身。 缺点:时间长,需及时发现优良性状。 三、单倍体育种: 单倍体育种是利用花药离体培养技术获得单倍体植株,再诱导其染色体加倍,从而获得所需要的纯系植株的育种方法。(主要是考虑到结合中学课本,经查阅相关资料无误。)其原理是染色体变异。优点是可大大缩短育种时间。 原理:染色体变异,组织培养 方法:选择亲本→有性杂交→F1产生的花粉离体培养获得单倍体植株→诱导染色体加倍获得可育纯合子→选择所需要的类型。 优点:明显缩短育种年限,加速育种进程。 缺点:技术较复杂,需与杂交育种结合,多限于植物。 四、多倍体育种: 原理:染色体变异(染色体加倍) 方法:秋水仙素处理萌发的种子或幼苗。 优点:可培育出自然界中没有的新品种,且培育出的植物器官大,产量高,营养丰富。缺点:只适于植物,结实率低。 五、细胞工程育种: 细胞工程育种是指用细胞融合的方法获得杂种细胞,利用细胞的全能性,用组织培养的方法培育杂种植株的方法。 原理:细胞的全能性 方法:(1)植物:去细胞壁→细胞融合→组织培养 (2)动物克隆:核移植→胚胎移植 优点:能克服远缘杂交的不亲和性,有目的地培育优良品种。动物体细胞克隆,可用于保存濒危物种、保持优良品种、挽救濒危动物、利用克隆动物相同的基因背景进行生物医学研究等。 两种转染试剂转染 C2C12 细胞效率比较分精品论文 两种转染试剂转染 C2C12 细胞效率比较分 析 韦伟,赵元元,张维娅,赵书红,李新云 5 ,农业动物遗传育种与繁殖教育部重点实验室~华中农业大学~武汉 430070, 摘 要:C2C12 细胞是鼠的骨骼肌成肌细胞~常用于体外研究肌细胞成肌分化~研 究表明 C2C12 细 胞的转染效率较低~为了提高 C2C12 细胞的转染效率~建立理想的转染条件 ~本研究对 比分析了 FuGENE HD 和 Lipofectamine 2000 两种常用转染试剂的转染效 率。研究结果表明 10 FuGENE HD 转染寡核苷酸的效率比 Lipofectamine 2000 高~而转染 质粒的效率比 Lipofectamine 2000 低。另外我们还发现培养基中的血清会降低细胞的转染 效率。本研究结 果为提高 C2C12 细胞的转染效率提供了新的信息。 关键词:转染效率,寡核苷酸,质粒,C2C12 细胞 中图分类号:Q-33 15 Compare analysis of the transfection efficiency of two transfection regents in C2C12 Cells Wei Wei, Zhao Yuanyuan, Zhang Weiya, Zhao Shuhong, Li Xinyun (Key Lab of Agricultural Animal Genetics, Breeding and Reproduction of Ministry of Education, 20 Huazhong Agricultural University, WuHan 430070) Abstract: C2C12 cells are the myoblast of mice, which are used as the model for investigating the differentiation of myoblast in vitro. The transfection efficiency of the C2C12 cells was not good in many studies. In order to improve the transfection efficiency of C2C12 cells and contribute an ideal condition of transfection. The transfection efficiency of two transfection reagents, FuGENE 25 HD and Lipofectamine 2000, was analyzed in this study. According the results, the transfection efficiency of FuGENE HD was higher than that of Lipofectamine 2000 when oligo nucleic acids was transfected, but it was lower than Lipofectamine 2000 when plasmid was transfected in the C2C12 cells. Also, we found that serum in cultured medium could inhibit the transfection efficiency. These results offered useful information for improving the transfection efficiency of 30 C2C12 cells. Key words: transfection efficiency; oligo nucleic acids; plasmid; C2C12 cells 0 引言 简转染是指将外源遗传物质转入到真核细胞内的过程。转染对现代分子生物学研究意义 各种排序法的比较 按平均时间将排序分为四类: (1)平方阶(O(n2))排序 一般称为简单排序,例如直接插入、直接选择和冒泡排序; (2)线性对数阶(O(nlgn))排序 如快速、堆和归并排序; (3)O(n1+£)阶排序 £是介于0和1之间的常数,即0<£<1,如希尔排序; (4)线性阶(O(n))排序 如桶、箱和基数排序。 各种排序方法比较: 简单排序中直接插入最好,快速排序最快,当文件为正序时,直接插入和冒泡均最佳。 影响排序效果的因素: 因为不同的排序方法适应不同的应用环境和要求,所以选择合适的排序方法 应综合考虑下列因素: ①待排序的记录数目n; ②记录的大小(规模); ③关键字的结构及其初始状态; ④对稳定性的要求; ⑤语言工具的条件; ⑥存储结构; ⑦时间和辅助空间复杂度等。 不同条件下,排序方法的选择 (1)若n较小(如n≤50),可采用直接插入或直接选择排序。 当记录规模较小时,直接插入排序较好;否则因为直接选择移动的记录数少于直接插人,应选直接选择排序为宜。 (2)若文件初始状态基本有序(指正序),则应选用直接插人、冒泡或随机的快速排序为宜; (3)若n较大,则应采用时间复杂度为O(nlgn)的排序方法:快速排序、堆排序或归并排序。 快速排序是目前基于比较的内部排序中被认为是最好的方法,当待排序的关键字是随机分布时,快速排序的平均时间最短; 堆排序所需的辅助空间少于快速排序,并且不会出现快速排序可能出现的最坏情况。这两种排序都是不稳定的。 若要求排序稳定,则可选用归并排序。从单个记录起进行两两归并,排序算法并不值得提倡,通常可以将它和直接插入排序结合在一起使用。先利用直接插入排序求得较长的有序子文件,然后再两两归并之。因为直接插入排序是稳定的,所以改进后的归并排序仍是稳定的。 几种控制方法的性能比较 专业: 控制理论与控制工程 姓名: 周燕红 学号: 200930210690 摘要:本文对同一控制对象分别采用常规PID 控制,模糊控制和基于遗传算法的PID 控制进行仿真,并对仿真结果进行分析,从而得出各个控制方法的性能优劣。 关键字:常规PID ;模糊控制器;遗传算法 1 常规PID 控制 1.1 PID 控制原理 在模拟控制系统中,控制器最常用的控制规律是PID 控制。模拟PID 控制系统原理框图如图1-1所示。系统由模拟PID 控制器和被控对象组成。 图1 PID 控制系统原理框图 简单说来,PID 控制器各校正环节的作用如下: (1) 比例环节:成比例的反应控制系统的偏差信号error(t),偏差一旦差生,控制器立即 产生控制作用,以减小偏差。P K 越大,系统的响应速度越快,调节精度越高,但易产生超调,甚至会使系统不稳定。反之,若过小,则调节精度降低,响应速度缓慢,使系统的静态、动态性能变坏。 (2) 积分环节:主要用于消除稳态误差,提高系统的误差度。积分作用的强弱取决于积 分时间常数I T ,I T 越大,积分作用越弱,若过大将使系统稳态误差难以消除,影响系统调节精度。反之则越强,稳态误差消除越快,但过小,在响应过程初期会产生积分饱和现象,从而引起响应过程的较大超调。 (3) 微分环节:反应偏差信号的变化趋势(变化速率),并能在偏差信号变得太大之前, 在系统中引入一个有效的早期修正信号,从而加快系统的动作速度,减少调节时间。微分作用的强弱取决于微分时间常数D T ,D T 越大,微分作用越强,但过大会使响应过程提前制动,而且会降低系统的抗干扰性能。 1.2衰减曲线法整定PID 参数 衰减曲线法是一种在经验凑试法基础上经过反复实验而得出的一种参数整定方法。可按过度过程达到4:1递减曲线法整定控制参数,也可按过度过程达到10:1递减曲线法整定控制参数。参数整定步骤: (1) 设置调节器积分时间Ti 为无穷大,微分时间常数为0,比例度为较大值,并将 系统投入运行。 (2) 系统稳定后,作设定值阶跃扰动,并观察响应。若系统衰减太快,则减小比例 #include 各种转染方法比较 转染方法原理主要应 用 特点主要的厂家及产品 DEAE-葡聚糖法带正电的DEAE-葡聚糖与核酸 带负电的磷酸骨架相互作用形成 的复合物被细胞内吞 瞬时转 染 相对简便、重复比磷酸钙好,但对细 胞有一定的毒副作用,转染时需除血 清且一般只用于BSC-1,CV-1,COS 细胞系 Sigma-Aldrich(DEAE-Dextran Transfection Kit) DNA复合物吸附细胞膜稳定转 染,染瞬 转染 不适用于原代细胞(所需的DNA浓 度较高),操作简便但重复性差,有些 细胞不适用 细胞建议用CSCL梯度离心,转染 是拷贝数较多 GIBCO BRL,Promega 阳离子脂质体法带正电的脂质体与核酸带负电的 磷酸基团形成复合物,然后脂质 体上剩余的电核与细胞膜上的唾 液酸残基的负电核结合;另一种 解释是通过细胞是内吞作用而被 进入细胞。(若DNA浓度过高, 中和脂质体表面电核,而降低了 与细胞的结合能力) 稳定转 染,瞬时 转染,所 有细胞 使用方法简单,可携带大片段DNA, 通用于各种类型的裸露DNA或 RNA,能转染各种类型的细胞,没 有免疫原性。虽在体外基因转染中 有很高的效率,但在体内,能被血 清清除,并在肺组织内累积,诱发 强烈的抗炎反应,导致高水平的毒 性,这在很大程度上限制了其应用 Invitrogen(Lipofectamine 2000,Lipofectamine, Lipofectin,Lipofectamine Plus,Cellfectin) Roche(Dosper,DOTAP,FuGENE 6) CPG Biotech Co(GeneLimo Plus,GeneLimo Super) Promega(Transfast,Tfx, Transfectam)数据结构各种排序方法的综合比较
各种转染试剂的中文转染方法
破乳的常用方法
几种常用的育种方法比较
两种转染试剂转染 C2C12 细胞效率比较分
各种排序法比较
几种控制方法比较
五种排序的算法(包括主函数调用)
各种转染方法比较