分子生物学ppt复习资料

第一章 绪论 1、分子生物学研究领域共同遵守的三大基本原则: ①构成生物大分子的单体是相同的(共同的核酸语言(Nt) 共同的蛋白质语言(aa)) ②生物遗传信息表达的中心法则相同 ③生物大分子单体的排列(核苷酸、氨基酸) 广义的分子生物学(作业中有) :泛指对生物大分子的研究,包括:蛋白质、核酸和多糖。 狭义的分子生物学(作业中有) :则仅指对核酸(包括 DNA 和 RNA)的研究 第二章 遗传物质的分子本质 末端终止法( Sanger and Coulson, 1977), 双脱氧链终止法:引物合成法或酶催引物合成法(其原理是:DNA 链中核苷酸以 3’,5’-磷酸二酯键连接, 合成 DNA 所用的底物是 2’-脱氧核苷三磷酸。 2’,3’ddNTP 与普通 dNTP 不同,它们在脱氧核糖的 3’位置缺少一个羟基。在 DNA 聚合酶作用下通过三磷酸基团掺入到延伸的 DNA 链中,但 由于没有 3’羟基,不能同后续的 dNTP 形成磷酸二酯键,因此,正在延伸的 DNA 链不能继续延伸。在 DNA 合成反应 混合物的 4 种普通 dNTP 中加入少量的一种 ddNTP, 链延伸将与偶然发生但却十分特异的链终止竞争, 产物是一系列 的核苷酸链,其长度取决于引物末端到出现过早链终止位置间的距离。在 4 组独立酶反应中分别采用 4 种不同的 ddNTP,结果将产生 4 组寡核苷酸,它们将分别终止于模板链的 A、C、G 或 T 位置。)。 1、 DNA polymerase(DNA 聚合酶)的两种特性: (1). DNA polymerase 能利用单链 DNA 作模板合成准确的 DNA 互补链 (2). DNA polymerase 能利用 2‘,3’-双脱氧 ddNTP 作底物,将其掺入至寡核苷酸链的 3‘端,从而终止 DNA 链的 生长。 2、DNA 的分子二级结构(作业中有 DNA 超螺旋结构) : (1)Main 主链:DNA 的密度表明 DNA 分子由两条反向平行(5’→3’)的围绕同一轴心旋转的多核苷酸链组成,脱 氧核糖通过 3’,5’-磷酸二酯键相连形成主链,呈右手双螺旋,处于螺旋的外侧。 (2)Base position:糖-磷酸键是在双螺旋的外侧,碱基位于螺旋内部,碱基的分子平面与螺旋轴垂直,同一平面的 碱基在两条主链间形成碱基对。 (3) Helical parameters(螺旋参数): 直径为 2nm,每一圈螺旋的高度为 34A°,由 10 个核苷酸组成,相邻核苷酸间呈 36 度角,距离为 3.4A° Helical turn: 10 base pairs/a turn ,3.4 nm/a turn (4)Base pairing(配对) :一条链的碱基与另一条链的碱基通过氢键相连形成碱基对。A 与 T,G 与 C 配对,分别 可形成 2 个和 3 个氢键。 (5) major or wide groove(大沟)和 minor or narrow groove (小沟): 从双螺旋中心到两条主链的连线将 DNA 的平面分 为两个不等的扇形,一个大于 180 度,一个小于 180 度,分别对应于大沟和小沟。 3、DNA 二级结构的稳定作用力 1.两条多核苷酸链间的互补碱基对之间的氢键 2.碱基对疏水的芳香环堆积所产生的疏水作用力,以及堆积的碱基对间的范德华力, 3.磷酸集团上的负电荷与介质中的阳离子化合物之间形成的盐键 4、染色体四级结构的特征 5、DNA 的三种螺旋:A 型、B 型和 Z 型 A 型:通常情况下我们观察到的是被认为适用于所有 DNA 的稳定形式即 B-DNA,在低温条件下,DNA 可被诱导 形成另一种螺旋,称为 A 型。A 行和 B 型一样为右旋,但较宽,结构更加紧密。碱基对倾斜于螺旋轴线,且偏离细 线。A 型螺旋重复每匝为 11 个碱基对。A 型的主要意义在于它是 RNA 及 RNA 与 DNA 杂合体的主要螺旋形式。 左旋 Z-DNA:在单一交替的嘧啶-嘌呤序列的合成 DNA 中会形成 Z-DNA 是稳定的。
1
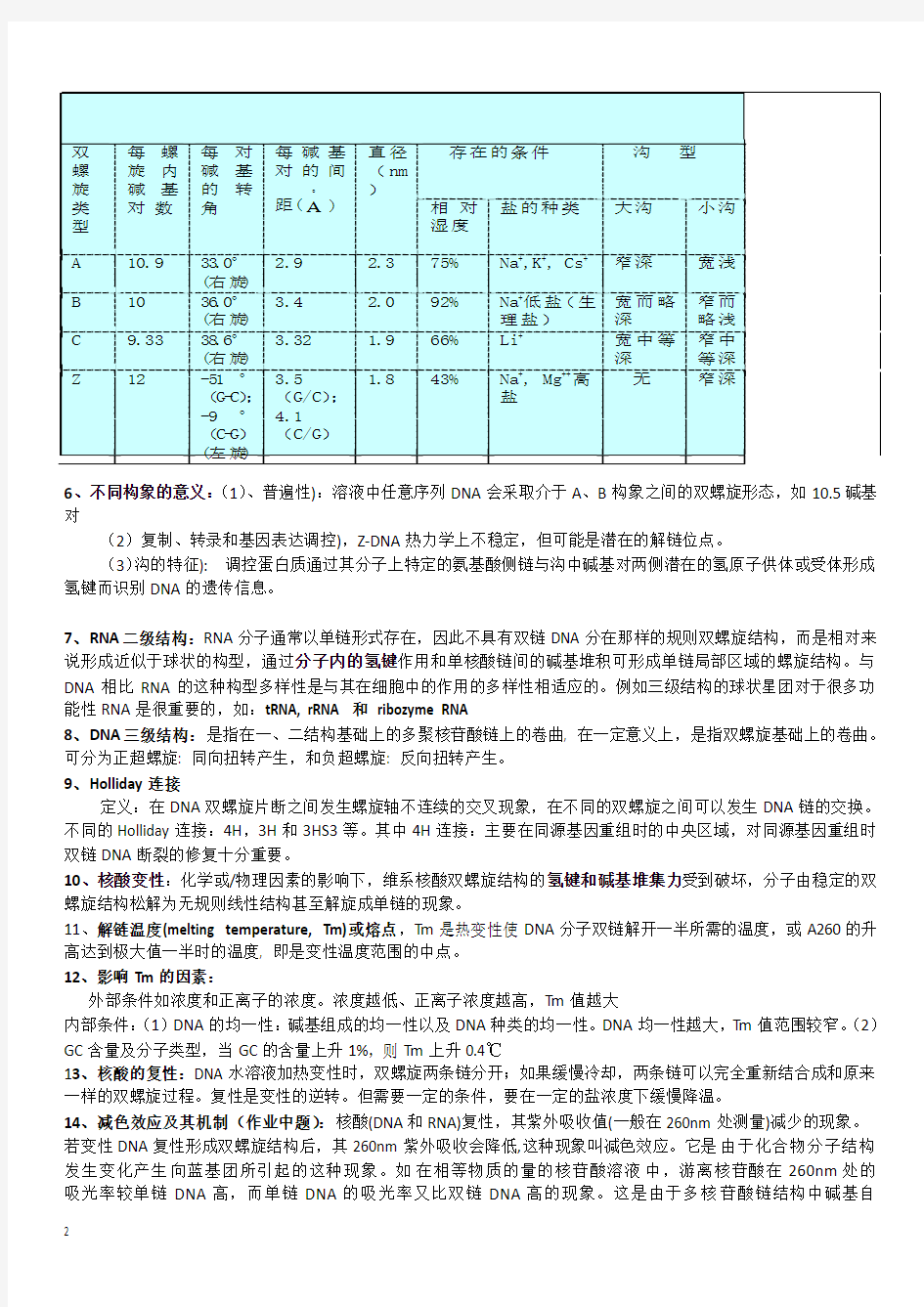
双 螺 旋 类 型 A B C Z
每 螺 旋 内 碱 基 对 数
每 碱 的 角
对 基 转
每 碱 基 对 的 间
?
直径 ( nm )
存在的条件
沟
型
距( A )
相 对 湿度 2.3 2.0 1.9 1.8 75% 92% 66% 43%
盐的种类
大沟
小沟
10.9 10 9.33 12
33.0 ° (右 旋 ) 36.0 ° (右 旋 ) 38.6 ° (右 旋 ) -51 ° ( C) G- ; -9 ° ( G) C(左 旋 )
2.9 3.4 3.32 3.5 (G/C) ; 4.1 (C/G)
Na+,K+, Cs+ Na+低盐 (生 理盐) + Li Na+, Mg++ 高 盐
窄深 宽而略 深 宽中等 深 无
宽浅 窄而 略浅 窄中 等深 窄深
6、不同构象的意义: 、普遍性):溶液中任意序列 DNA 会采取介于 A、B 构象之间的双螺旋形态,如 10.5 碱基 (1) 对 (2)复制、转录和基因表达调控),Z-DNA 热力学上不稳定,但可能是潜在的解链位点。 (3) 沟的特征): 调控蛋白质通过其分子上特定的氨基酸侧链与沟中碱基对两侧潜在的氢原子供体或受体形成 氢键而识别 DNA 的遗传信息。 7、RNA 二级结构:RNA 分子通常以单链形式存在,因此不具有双链 DNA 分在那样的规则双螺旋结构,而是相对来 说形成近似于球状的构型,通过分子内的氢键作用和单核酸链间的碱基堆积可形成单链局部区域的螺旋结构。与 DNA 相比 RNA 的这种构型多样性是与其在细胞中的作用的多样性相适应的。例如三级结构的球状星团对于很多功 能性 RNA 是很重要的,如:tRNA, rRNA 和 ribozyme RNA 8、DNA 三级结构:是指在一、二结构基础上的多聚核苷酸链上的卷曲, 在一定意义上,是指双螺旋基础上的卷曲。 可分为正超螺旋: 同向扭转产生,和负超螺旋: 反向扭转产生。 9、Holliday 连接 定义:在 DNA 双螺旋片断之间发生螺旋轴不连续的交叉现象,在不同的双螺旋之间可以发生 DNA 链的交换。 不同的 Holliday 连接:4H,3H 和 3HS3 等。其中 4H 连接:主要在同源基因重组时的中央区域,对同源基因重组时 双链 DNA 断裂的修复十分重要。 10、核酸变性:化学或/物理因素的影响下,维系核酸双螺旋结构的氢键和碱基堆集力受到破坏,分子由稳定的双 螺旋结构松解为无规则线性结构甚至解旋成单链的现象。 11、解链温度(melting temperature, Tm)或熔点,Tm 是热变性使 DNA 分子双链解开一半所需的温度,或 A260 的升 高达到极大值一半时的温度, 即是变性温度范围的中点。 12、影响 Tm 的因素: 外部条件如浓度和正离子的浓度。浓度越低、正离子浓度越高,Tm 值越大 内部条件: (1)DNA 的均一性:碱基组成的均一性以及 DNA 种类的均一性。DNA 均一性越大,Tm 值范围较窄。 (2) GC 含量及分子类型,当 GC 的含量上升 1%,则 Tm 上升 0.4℃ 13、核酸的复性:DNA 水溶液加热变性时,双螺旋两条链分开;如果缓慢冷却,两条链可以完全重新结合成和原来 一样的双螺旋过程。复性是变性的逆转。但需要一定的条件,要在一定的盐浓度下缓慢降温。 14、减色效应及其机制(作业中题) :核酸(DNA 和 RNA)复性,其紫外吸收值(一般在 260nm 处测量)减少的现象。 若变性 DNA 复性形成双螺旋结构后,其 260nm 紫外吸收会降低,这种现象叫减色效应。它是由于化合物分子结构 发生变化产生向蓝基团所引起的这种现象。如在相等物质的量的核苷酸溶液中,游离核苷酸在 260nm 处的 吸光率较单链 DNA 高,而单链 DNA 的吸光率又比双链 DNA 高的现象。这是由于多核苷酸链结构中碱基自
2
由旋转受阻所致。通过在波长 260nm 处记录溶液的光密度,能够追踪 DNA 的变性作用。 15、增色效应及其机制(作业中题) :核酸(DNA 和 RNA)分子解链变性或断链,其紫外吸收值(一般在 260nm 处测量) 增加的现象。DNA 分子具有吸收 250 - 280nm 波长的紫外光的特性,其吸收峰值在 260nm 。 DNA 分 子中碱基间电子的相互作用是紫外吸收的结构基础 , 但双螺旋结构有序堆积的碱基又 " 束缚 " 了这种作 用。变 性 DNA 的双链解开 , 碱基中电子的相互作用更有利于紫外吸收 , 故而产生增色效应。 16、DNA 的复性对片段有两个要求: (1) 互补顺序的碰撞和排列; (2) 碱基的正确配对和氢键的形成。 17、拓扑异构酶(作业中有,见下 18) :能够调控 DNA 分子超螺旋水平的酶。为了改变 DNA 的连接数,拓扑异构 酶必须暂时断裂 DNA 分子的一条或两条链,通过攻击磷酸骨架上的酪氨酸残基,使自身与 DNA 的一端形成磷酸酪 氨酸键以建立临时的共价连接。共有两类拓扑异构酶:Ⅰ型拓扑异构酶在 DNA 的一股链上产生一个切口,使另一条 链得以穿越,连接数每次改变+1;而Ⅱ型酶由 ATP 的水解提供能量,则在 DNA 的双链上产生切口,使另一双链 DNA 片段得以穿过,每次连接数改变+ 2。 18、 (作业中题)对于右手螺旋 DNA 分子来说,每一圈螺旋由 10 个碱基对组成。如果每圈初级螺旋的碱基对数小 于 10.5,则其二级结构处于紧缠状态,由此产生的超螺旋为( 负超螺旋 ) ,反之为(正超螺旋 ) 。 DNA 超螺旋由 DNA topoisomerase ( 拓扑异构酶 )产生。 DNA topoisomerase: 兼有 DNA(内切)酶和 DNA(连接 )酶的功能。--------see (DNA 重组)。 第三章:基因,基因组与基因组学 移动基因(作业中题) :也叫转位因子,是指能够在一个 DNA 分子内部或两个以上 DNA 分子之间移动的 DNA 片段。 在细菌中指在质粒和染色体间或质粒和质粒之间移动的 DNA 片段。是 DNA 重组的一种形式。细的转位因子包括插 入序列,转座子及可转座的噬菌体。 断裂基因(作业中题) :真核生物结构基因,由若干个编码区和非编码区互相间隔开但又连续镶嵌而成,去除非编 码区再连接后,可翻译出由连续氨基酸组成的完整蛋白质,这些基因称为断裂基因 假基因(作业中题) :基因组中存在的一段与正常基因非常相似但不能表达的 DNA 序列。分为两大类:一类保留了 相应功能基因的间隔序列,另一类缺少间隔序列,称为加工过的假基因或返座假基因。 重叠基因(作业中题):指两个或两个以上的结构基因共同一段 DNA 顺序的现象,重叠基因仅在噬菌体和病毒中 存在。 顺式作用元件:指同一 DNA 分子中具有转录调节功能的特异 DNA 序列。包括启动子、增强子,调控序列和可诱导 序列等。 反式作用因子:指能直接或间接地识别或结合在各类顺式作用元件核心序列上参与调控靶基因转录效率的蛋白质。 多为转录因子。 SD sequence:原核生物基因含有核糖体结合位点(ribosome-binding site, RBS),转录产生的富含嘌呤的序列可以 与核糖体 16S rRNA3?-端富含嘧啶的序列互补配对,帮助翻译的正确起始。真核生物无 SD 序列,40S 核糖体与 mRNA5?-端的“帽子”结构相互作用, 帮助翻译的正确起始。 高等真核生物, mRNA 的 3?-端有一段保守序列 AAUAAA, 与 3?-端的加工和多聚腺苷酸化有关,称为加尾信号。 C 值(C value,作业中题): 一个单倍体基因组的全部 DNA 含量总是恒定的,是物种的一个特征,称之。不同 物种 C 值:从小于 106bp 至 1011bp。真菌和高等植物同属于真核生物,而后者的 C 值大得多。 C 值矛盾(C value paradox,作业中题): 生物体复杂性和 DNA 含量之间并不总是正相关。表现在: ① 与预期 的编码蛋白质的基因数量相比,基因组 DNA 的含量过多;② 一些物种之间的复杂性变化范围并不大,而 C 值却有 很大变化。 模式生物(Model orgnism,作业中题):通过对选定的生物物种进行科学研究,用于揭示某种具有普遍规律的生命 现象,这种被选定的生物物种包括动物、植物和微生物称为模式生物。大肠杆菌、酿酒酵母、拟南芥菜、秀丽应隐 杆线虫、果蝇、小鼠等都是已完成基因组测序的模式生物。 基因家族: 真核生物基因组中来源相同、结构相似、功能相关的一组基因。按家族成员分布形式可分为基因簇和散 布基因家族。
3
基因簇:基因家族的各成员紧密成簇排列成大段的串联重复单位,定位于染色体的特殊区域。也包括没有功能的假 基因。 Interspersed gene family(散布基因家族):家族成员在 DNA 上无明显的物理联系,甚至分散在多条染色体上, 各成员在序列上有明显差别,其中也含有假基因(但来源于 RNA 介导的转座作用)。 重复序列:染色体上大量无转录活性的重复 DNA 序列家族,主要是基因以外的 DNA 序列。 卫星 DNA:高度重复 DNA 序列的碱基组成和浮力密度同主体 DNA 有区别,在浮力密度梯度离心时,可形成不同 于主 DNA 带的卫星带。 基因组学(作业中题) :指对所有基因进行基因组作图(包括遗传图谱、物理图谱、转录图谱)、核苷酸序列分析、基 因定位和基因功能分析的一门学科,是从基因组水平研究遗传的科学。 结构基因组学(作业中题) :包括基因组作图和基因组测序。研究基因组的结构并构建高分辨率的遗传图、物理图、 序列图和转录图以及蛋白质组成与结构的学科。 功能基因组学(作业中题) :利用结构基因组学研究所得的各种信息在基因组水平上研究编码序列及非编码序列生 物学功能的学科。 蛋白质组学(Proteomics, 作业中题): 对蛋白质性质和功能的大规模研究, 包括不同时相细胞内蛋白质的表达水平、 翻译后修饰以及与其他分子的相互作用的研究,揭示正常和疾病状态下,蛋白质表达的规律,从而研究疾病发生机 理并发现新药。 蛋白质组(proteome):基因组表达的全部蛋白质,是一个动态的概念,指的是某种细胞或组织中,基因组表达的所 有蛋白质。 转录组学(作业中题) :是一门在整体水平上研究细胞中基因转录的情况及转录调控规律的学科。简而言之, 转录组学是从 RNA 水平研究基因表达的情况。 外显子(exon) 是指在结构基因中,有编码作用的 DNA 序列。外显子比内含子多一个。 内含子(intron) 指位于两个外显子之间没有编码作用的 DNA 序列。内含子序列比外显子多。 DNA 的遗传多态性标记及其种类(作业中题) :答:包括以下三类 (1) 、.第一代遗传标记 RFLP:用限制性内切酶特异性切割 DNA 分子,由于 DNA 的点突变而产生不同长度的等位 片段,可用凝胶电泳显示多态性,用于基因突变分析、基因定位和遗传病基因的早期检测等。 (2) 、.第二代遗传标记 小卫星 DNA 重复序列多态性:基因组 DNA 中有数十到数百个核苷酸片断的重复,重复 的次数在人群中高度变异。 (3) 、.第三代遗传标记 微卫星 DNA 重复序列或短串联重复(STR)多态性:基因组中 1-6 个碱基的重复,如 (CA)n,(GT)n 等产生,以 CA 重复序列的利用度为最高。在染色体 DNA 中散在分布,数量可达 5 到 10 万,是目前最 有用的遗传标记。 “RNA 世界”假说及支持该假说的主要生化及分子生物学证据(作业中题) : 答:“RNA 世界”假说内容为生命进化的早期,没有蛋白质(酶) ,某些 RNA 可以催化 RNA 的复制——也就是说 RNA 是唯一的遗传物质,是生命的源头。 但 RNA 是唯一的既能携带遗传信息又可以是功能分子的生物高分子化合物。因此,生命发生之初,很可能 是在原始海洋深处的火山口边,高温、高压的条件下,在可作为催化剂的矿物质边富集了可能是雷电中合 成的原始核苷酸。经过亿万年的进化,形成了具有自我复制能力的 RNA.在人工条件下,这种进化的某些过 程已被成功地模拟。原始的具有自我复制能力的 RNA,再在以后的亿万年进化过程中,逐渐将其携带遗传信 息的功能传给了 DNA,将其功能分子的功能传给了蛋白质。核糖体是核酶的发现大大支持了 RNA 世界的假 说。 启动子(promoter): 启动子是位于基因转录起始点上游的一段特定的 DNA 顺序,是 RNA 聚合酶与之相识别、结 合的部位,从而启动基因的转录。 原核启动子区:两个保守序列-35 序列和-10 序列。 真核启动子区:TATA 框、CAAT 框、GC 框等。 增强子( enhancer): 增强子是指能够增强基因转录作用的一段特定的 DNA 顺序,其作用是增强启动子效应,与 基因的转录启动无关。增强子的位置比较灵活,它可以位于转录起始点的上游,也可以位于转录起始点的下游。它
4
通过与特异性的蛋白质结合而促进基因的转录。 终止子(terminator): 终止子是一段具有转录终止功能的特定 DNA 顺序。位于编码区下游,转录终止点上游。原 核生物终止子含有回文序列,产生的 RNA 可以形成发夹结构,使 RNA 聚合酶减慢移动或暂停 RNA 的合成。 第四章 反向复制 一、DNA 复制的基本特点: (1、半保留复制:在复制中,DNA 双螺旋的两条互补链解开,分别作为模板指导以脱氧核苷酸 5?-三磷酸为前体由 5?→3?方向合成新生互补链。这样每个子细胞接受亲代 DNA 双链中的一条。 (2、半不连续复制:在半保留复制中,两条 DNA 的新生链合成在同一复制叉中同时进行,但 DNA 复制原则上只 能从 5?→3?方向进行,因为两条 DNA 链是反向平行的,所以前导链是从起始点按 5?→3?方向进行连续复制,而后随 连不立即复制,其复制从复制叉处起始,按 5?→3?方向朝向起始点形成第一个冈崎片段。随着复制叉的前进,前导 链继续被复制成连续长链,而后随链按反方向以不连续方式被复制。合成不久后冈崎片段被 DNA 连接酶连成一条 连续的 DNA。 (3、RNA 引导:冈崎片段和前导链的头几个一样,每个片段 5? 端的头几个核苷酸均是核糖核苷酸,因此 DNA 合 成是由 RNA 引导的。这些引物在片段连接之前被去掉,产生间隙由 DNA 填补。用 RNA 引导 DNA 复制的原因很 可能是出于保证 DNA 复制的高忠实性。 二、DNA 复制的相关的酶和蛋白 (1、DNA 聚合酶(DNA pol):以单链或双链 DNA 为模板,催化由脱氧核糖核苷三磷酸合成 DNA 的酶。在原核细 胞有 DNA 聚合酶Ⅰ、Ⅱ、Ⅲ。真核生物中有 DNA 聚合酶 α、β、γ、δ、ε 五种,其中 δ 为主要的聚合酶,γ 存在于 线粒体中。 (1)原核生物 DNA 聚合酶: DNA-polI: 能催化脱氧核苷酸加到引物链的 3′ -OH 末端, 引物延伸方向 5′ 3′ 该酶需要的条件: 种 dNTP、 → 。 4 Mg 、DNA 模板(template)、引物(primer),此酶有三种活性:5′ 3′ → 聚合酶,5′ 3′ 外切酶(主要功能是 → 切除引物,填补冈崎片段产生的空隙及 DNA 损伤的修复),3′ →5′外切酶(校正活性)。具有的聚合酶和 5’-外 切酶活性的配合使用可导致本来一条链带有切口的 DNA 分子发生切口平移 polⅡ:5′ 3′ → 聚合酶活性,3′ →5′外切酶 polⅢ:是大肠杆菌主要的 DNA 聚合酶,其全酶由 10 种亚基组成,α 、ε 、θ 组成核心酶,α 亚基具有 5′ 3′ → 方向合成 DNA 的催化活性,ε 亚基具有 3′ 5′ → 核酸外切酶的活性,起校对作用。DNA polⅢ为异二聚体,使 DNA 解开的双链可同时进行复制。这种复杂的亚基结构使其具有更高的忠实性、协同性和持续性。 (2)真核生物 DNA 聚合酶:包括 α、β、γ、δ 和 ε γ 参与线粒体 DNA 复制; α 及 δ、ε 参与核染色体 DNA 复制; α:链合成的引发,具引发酶,无 3?-外切酶活性 δ 和ε :链的延长,具 3?-外切酶活性,校对功能。PCNA 为 δ 的辅助蛋白,三个亚基组成滑动钳,以提高 δ 的进行性。 β:两个结构域,N-端为 5’-脱氧核糖磷酸酶和与单链 DNA 结合的活性;C-端具聚合酶活性。参与 DNA 损伤的 修复,增补 DNA 链上短的空隙。 γ:线粒体 DNA 复制和损伤修复,具聚合酶、3’-外切酶和 5’-脱氧核糖磷酸酶活性。 (2、拓扑异构酶:DNA 复制时必先要解旋和解链,拓扑异构酶对 DNA 分子兼有内切酶和连接酶的作用,有I型 和Ⅱ型。前者可切断双链中的一条链,使解旋中不致打结(适时又把切口封闭,DNA 呈松弛态,这过程不耗能)。 后者在无 ATP 供能时,可同时切开超螺旋状态 DNA 的两条链,使其松弛,然后将切口封闭(用于分离复制后的两 个子环)。在利用 ATP 时,该酶可使松弛态的 DNA 变成负超。(详见 p3-17)
5
2+
DNA 的复制
(作用机制:两次转酯反应。 第一次由酶活性中心的 1 个 Tyr-OH 亲核进攻 DNA 链上的 3?, 5?-磷酸二酯键,致 DNA 链断裂,形成磷酸酪氨 酸酯键相连的酶与 DNA 的共价中间物; 之后,DNA 的另一条链或另一 DNA 双螺旋通过切口,导致其拓扑学性质发生改变; 第二次由 DNA 断裂处的自由 OH 亲核进攻酶第一次转酯反应形成的磷酸酪氨酸酯键,导致断裂的 3?, 5?-磷酸二 酯键重新生成。) (3、DNA 解链酶(DNA helicase):解链酶可通过水解 ATP 供能来解开双链,每解开一对碱基,需水解 2 分子 ATP。 该酶和 rep 蛋白共同参与解链,rep 蛋白是沿着前导链的模板(母链)3′ →5′方向移动,而解链酶按母链 5′ →3′ 方向移动。 (4、引物酶(primase)和引发体:DNA 聚合酶没有催化两个游离 dNTP 聚合的能力,而 RNA 聚合酶依靠模板 可酶促游离 NTP 聚合,生成的一段短 RNA 引物提供 3′OH末端供 dNTP 加入、延长。所以,解开双链并不是马上 进行复制,先以模板脱氧核苷酸序列,按碱基互补原则合成一小段 RNA 引物,这一过程称“引发”。催化 RNA 引 物合成的 RNA 聚合酶称引物酶(或引发酶)。该酶只有与相关的蛋白结合为引发体才有明显的活性。引发体是解 链酶、DnaC、引物酶和 DNA 起始复制区组成的复合结构。 (5、 DNA 连接酶 (DNA ligase) 催化相邻 DNA 片段间甚至两个双螺旋 DNA 分子两端的连接, : 即把有缺口的 3′ - OH 末端与相邻核苷酸 5′-磷酸连接形成磷酸二酯键,连接反应是耗能的。 (6、单链结合蛋白(single-strand bindingprotein,SSB):一旦 DNA 双螺旋解开成单链,SSB 便牢固地结合 到分开的单链上,防止它们重新形成双螺旋,保证模板链的复制和不被核酸内切酶水解。 (7、端粒和端粒酶:人的 DNA 端粒含有 TTAGGG 重复序列,长度 5~15kb。端粒的功能:保护染色体末端免于 化学修饰或核酸酶降解以及不正常的融合和重组;解决染色体复制时末端丢失问题。 (8、端粒酶及其复制机制(作业中题):端粒酶是一种核糖核蛋白,由 RNA 和 protein 组成的不依赖 DNA 的 DNA 聚合酶,可以与自身携带的 RNA 模板合成端粒重复序列,兼有模板和逆转录酶两方面的作用。 端粒酶的复制机制: 端粒酶以“滑动”机制(slippage mechanism)来延长端粒的长度,每合成 1 拷贝的重复序 列,酶滑至新的端粒末端,重新启动重复序列的合成。 过程: RNA 中的端粒 DNA 重复序列与端粒 DNA 最后一段重复序列互补配对, 其 而其余的重复序列凸出在端粒 的一侧作为模板; 随后发生逆转录反应, 在端粒 DNA 的 3?-端添加 1 拷贝的重复序列(GGGTTG)。 随后端粒酶移位, 重复上 述过程,直到端粒突出的一端能够作为合成新的冈崎片段的模板,以填补上一个冈崎片段 RNA 被切除后留 下的空隙。 三、DNA 复制的过程 (1、DNA 复制的起始阶段 (1)复制子:基因组中能单独进行复制的单位,每个起始点到终止点的区域为一个复制子。原核生物 DNA 分子 中只有一个复制起始点(ori),真核生物有多个 ori。 复制是从特定部位即复制起始点开始,复制的基本单位为复制子 ? Helicase (DNA 解链酶):水解 ATP 获能解开双链 DNA ? SSB 蛋白 (单链结合蛋白):保持单链结构 ? Primase (引发酶 or RNA 聚合酶):合成 RNA 引物 ? 随从链是由引发体来完成的 (2、DNA 链的延长 复制的延伸在复制内进行。是以 DNA 为模板在 DNA 聚合酶作用下,将四种 dNTP 聚合成 DNA 的过程。 (3、DNA 复制的终止阶段 E.coli DNA 具有复制终止位点 Ter(6 个) ,此处可以结合一种特异的蛋白质分子 Tus,通过阻止 Dna B 解链酶 的解链活性而终止复制。
6
子代 DNA 的分离:拓扑异构酶 IV 或 XerCD (位点特异性重组酶)识别终止区内的 dif 位点,切开以连环体相连 的两子代 DNA 分子(去连环化),在交换后实行再连接。 真核与原核生物 DNA 复制大体相同,差别为: 1、染色质和核小体结构对 DNA 复制的影响 2、复制叉移动的速度远远低于原核细胞 3、真核生物是多点复制,具多个复制起始区,原核生物是单点复制。 4、冈崎片段的长度短于原核细胞 5、复制严格限制在细胞周期的 S 期 6、真核生物在完成全部复制之前,各个起始点上 DNA 的复制不能再开始;原核生物起始点上可连续开始新的 DNA 复制。 7、真核生物线性 DNA 末端具有端粒结构。 保证 DNA 复制高度忠实性的机制 1、四种 dNTP 浓度的平衡: 由核苷酸还原酶(nucleotide reducetase) 2、DNA pol 的高度选择性:几何选择(geometric selection)和构象变化(confromational changes) 3、 DNA pol 的自我校对:3?-核酸外切酶 4、错配修复 5、RNA 作为引物:DNA 合成开始参入的核苷酸难与模板链形成稳定的双螺旋,易发生错配,RNA 作为引物 则可减少错配 逆转录:以 RNA 为模板合成 DNA,存在于逆转录病毒以及原核生物和真核生物如端粒酶的延长等。 逆转录酶是一种多功能酶: 1、RNA 指导的 DNA 聚合酶活性 2、RNA 酶 H(RNaseH)活性:水解 RNA-DNA 杂交体上的 RNA 3、DNA 指导的 DNA 聚合酶活性:合成互补 DNA。 4、没有 3`→5`外切酶活性。 第五章、DNA 损伤、修复和突变 DNA 损伤的因素(作业中题) A、细胞内在因素: ? ? ? DNA 结构本身的不稳定; DNA 复制过程中自然发生的错误,主要是碱基错配; 细胞内活性氧(reactive oxygen species, ROS)的破坏作用 ;
B、环境因素有化学因素和物理因素,前者包括各种化学诱变剂如黄曲霉素(aflatoxin)、芥子气、烷基化试剂和癌 症化疗药物顺铂(cis-platinum),后者包括紫外辐射和离子辐射(ionizing radiation, IR) DNA 自发性损伤:由于 DNA 内在的化学活性以及细胞中存在的正常活性化分子所致。包括:碱基错配,脱氨基作 用 (环外氨基),碱基丢失 → 自发水解,碱基修饰与链断裂。 氧化性损伤:需氧细胞在代谢过程中产生一系列活性氧簇( reactive oxygen species, ROS),包括:超氧化物、氢 过氧化物及羟基自由基等,会在正常条件下发生氧化损伤。 烷基化引起 DNA 损伤:1、碱基烷基化: G-C→A-T ;2、 碱基脱落: 甲基磺酸甲酯可使鸟嘌呤 7N 烷基化,活 化 β-糖苷键,连接碱基与五碳糖间的共价键变弱,容易折断缺失碱基,造成脱嘌呤作用。 3、导致 DNA 断链: 磷 酸二酯键上的氧被烷基化;4、导致 DNA 链交联 电离辐射引起 DNA 损伤的类型:1、产生 OH 自由基,导致碱基变化;2、脱氧核糖分解;3、 DNA 链断裂:单链 断裂(无差错修复)和双链断裂(错误修复);4、DNA 链、蛋白质的交联。 DNA 损伤类型: (1、碱基损伤
7
a 、碱基丢失:黄曲霉素致脱嘌呤反应。 b、 碱基转换:自发或在某些化学试剂如亚硝酸作用下发生脱氨基反应,如 A 和 C 转变成 I 和 U。 c、 碱基修饰:如烷基化试剂。 d、 碱基交联:紫外线 e、 碱基错配 (2、DNA 链的损伤 a、 链的断裂:离子辐射(X 和γ 射线)和化学试剂如博来霉素。 b、 DNA 链间的交联:双功能试剂,如顺铂和丝裂霉素 C (citomycin C, MMC)。 c、DNA 与蛋白质之间的交联:UV ★★★DNA 修复(作业中题) DNA 的修复主要类型:直接修复;切除修复;双链断裂修复;易错修复;重组修复;跨损伤修复 (SOS 修复) 常见的 DNA 损伤及其修复机制 DNA 损伤因素 X 射线、氧自由基、烷化剂 自发脱 碱基 DNA 损伤类型 单链断裂、无碱基位点、氧 化性碱基(如 8-氧鸟嘌呤) 尿嘧啶 环丁烷嘧啶二聚体等大的 紫外线光产物和稳定的多 环芳烃化合物等大分子 DNA 加合物 双链断裂和链间交联 碱基错配和缺失(插入) 修复机制 碱基切除修复
紫外线和多环芳烃
核苷酸切除修复
抗癌药(如顺铂和丝裂霉素) 复制错误和烷化剂 一、直接修复:
双链断裂修复(同源 重组修复和末端连 接) 错配修复
(1)DNA 断裂口直接修复:在 DNA 5?-P 端和 3?-OH 端未受损害的情况下,DNA 连接酶能够直接修复 DNA 的断裂口。 (2)DNA 紫外线损伤的光复合酶直接修复。DNA 光解酶(光复活酶)能够直接识别和结合嘧啶二聚体,并利用 作为捕光色素的辅基所吸收的光能,将环丁烷环打开。具有专一性,是无差错的。 (3)烷基化碱基的直接修复,烷基转移酶能够转移碱基上的烷基和甲基化磷酸二酯键上的甲基,烷基被转移带 该蛋白本身后使之失活。 二、切除修复:(上课做过题目的)切除修复是与转录相偶联的,因而被转录 DNA 比修复比未被转录 DNA 区域修 复得更快,这有助于限制缺陷基因产物的生成。 (1)碱基切除修复( BER),指切除和替换 DNA 单链断裂和小的碱基改变及氧化性损伤等产生 的 DNA 碱基损伤,由多个酶完成,先后经历识 别、切除、重新合成和重新连接四步。 其过程如左图所示:ab、相当专一的 DNA 糖基 化酶识别修饰碱基与糖基之间的 N-糖苷键, 留下 一个脱嘌呤或脱嘧啶(AP)位点。自发碱基丢失 也可以产生 AP 位点。
8
c、 内切核酸酶(5‘-和 3’-两种)在该位点切开 DNA, AP 其外切酶活性可继续切除 1 个缺口(切除 5?-脱氧核糖磷酸)。 d、E.coli 中的缺口可由 DNA 聚合酶Ⅰ填补。 e、最后的磷酸二酯键由 DNA 连接酶完成。 注:在真核生物中,BER 中的缺口主要由 DNA 聚合酶β 来填补。
(2) 核苷酸切除修复(NER) ? 体内识别 DNA 损伤最多的修复通路,主要用来修复导致 DNA 结构发生扭曲并影响到 DNA 复制的损伤, 如 DNA 约 30° 弯曲的嘧啶二聚体 ? 不识别任何特殊的碱基损失,而是识别双螺旋形状的改变;修复时切除含有损伤碱基的那一段 DNA ? 起始切点:损伤部位附近的 3`,5`-磷酸二酯键 主要由 5 步反应组成: ? 1 探测损伤:特殊蛋白与受损伤 DNA 的有序结合 ? 2 切开损伤链:内切酶在损伤部位的两侧各切除精确数目的碱基以切开 DNA 链 ? 3 去除损伤:2 个切口之间的损伤 DNA 片断 ? 4 填补缺口:DNA pol 完成 ? 5 缝合切口:DNA 连接酶 在真核生物中,NER 中较长的缺口主要由 DNA 聚合酶 δ 或 ? 来填补。 NER 可进一步分为全局性基因组 NER(global genome NER, GGR)和转录偶联性 NER (transcription-coupled NER, TCR),两者主要差别在于识别损伤的机制不同。 GGR: 负责整个基因组的损伤,速度慢,效率低 TCR:专门修复正在转录的基因在模板链上的损伤,速度快,效率高,由 RNA 聚合酶识别损伤,当聚合酶转录 到受损伤的部分而前进受阻时,TCR 即启动。 错配修复(MMR):纠正 DNA 双螺旋上错配的碱基对(非 Watson-Crick)碱基对,此外还能修复一些因“复制打滑” 而诱发产生的核苷酸插入或缺失环(4nt), 使复制的保真性提高 102~103 倍 过程 ? ? ? ? 1、识别新生链中非 m6A 的 GATC 序列 2、扫描新生链中错配碱基 3、酶切含错配碱基的 DNA 区段 4、重新合成 DNA
三、双链断裂修复(DSBR) 由于难以找到互补链,难以彻底修复。所以双链断裂有两种修复机制: (1) 同源重组修复:通过从同源染色体获得修复断裂的模板,精确性较高。 (2) 非同源末端连接(NHEJ): 在无序列同源的情况下,让断裂的末端重新连接,精确性低,是人类修复双链 断裂主要方式。 四、损伤跨越:某些情况,损伤无法修复(如复制叉已打开母链,使切除修复系统无法利用互补链作为修复合成的 模板),或修复系统还没有机会去修复。为维持复制的连续性,细胞利用以下两套方式:重组跨越或重组修复、跨越 合成。 (1) 重组修复机制
9
重组修复: 利用同源重组将 DNA 模板进行交换以克服损伤对复制的障碍, 而随后的复制仍使用细胞内高保真的 聚合酶。 DNA 复制时会越过受损区域进行,经重组修复,受损的 DNA 仍然存在于子代的一个细胞中。 (2) 跨越合成(TLS) TLS 又称跨损伤合成:由特殊的 DNA pol 取代在损伤位点处催化复制的 DNA pol(原核 DNA pol III,真核为聚 合酶 α,δ 和 ε), 在子链上(模板链上损伤碱基的对面)随机插入核苷酸(正确的或错误的), 以实现对损伤位点无错或易 错的跨越。 五、SOS 修复(非常重要) 指细胞在受到潜在致死性压力(如 UV 辐射、胸腺嘧啶饥饿、丝裂霉素 C 作用、DNA 复制必需基因失活等因素) 之后,出现有利于细胞生存、以突变为代价的代谢预警反应。 SOS 修复诱导 DNA 聚合酶活性,涉及近 20 个 sos 基因的表达,整个反应受到阻遏蛋白-LexA 和激活蛋白 -RecA 的调节。LexA (一种 DNA 结合抑制蛋白), umuC 和 umuD, 编码 DNA 聚合酶活性, 允许复制跨过损伤 位点, 常插入一个或几个 A 碱基。 细胞周期检查点控制 物细胞 DNA 受到损伤时细胞除诱导修复基因的转录外,还可暂时阻断细胞周期,防止受损 DNA 继续复制,如 无法修复,则可诱导细胞进入凋亡。细胞通过细胞周期检查点控制(又称关卡控制)对 DNA 损伤的应答反应 检查点:细胞周期中可以终止细胞分裂或周期进程的那些点成为检查点。检查点在间隙期发挥作用,以保证细胞 有能力进行又一轮的 DNA 复制(G1 期的 R 点)及在细胞分裂之前成功完成 DNA 的复制(G2 期检验点)。 DNA 损伤和修复的生物学意义: 对于基因组的不稳定性、癌症和细胞死亡是至关重要的。DNA 修复途径可以识别和修复特异的 DNA 损伤,保
染色体畸变 碱基错配 复制错误 突变 自发突变 基因突变 诱发突变 脱嘌呤 化学错误 脱氨基 氧化损伤 放射线----产生嘌呤二聚体 碱基类似物:5BU,2AP 化学诱变剂 碱基修饰物:NA,HA,烷化剂 DNA 插入剂:吖啶类 DNA 复制跳格
证生物物种的遗传稳定性。 DNA 的突变 发生在 DNA 分子上可遗传的永久性结 构变化通称为突变。 带有一个给定突变的基因、基因组、细 胞或个体被称为突变体。 DNA 突变的种类(作业中题): 1、 点突变: 碱基对转换,指 DNA 分子单 一位点上所发生的碱基对改变,分为: 转换(是指同类碱基之间的替换。)
表 21-1 突变的各种类型 点突变,碱基替换 转换 Py 与 Py,Pu 与 Pu 之间变换,多见 颠换 Py 与 Pu 之间变换,少见 插入 1-2 个碱基 移码突变 丢失 1-2 个碱基 缺失突变
10
颠换(是嘌呤与嘧啶之间的替换。) 点突变的后果: 取决于发生的位置和具 体的突变方式 发生在蛋白质基因编码区的点突变: 沉默突变:突变的密码子编码同样的 aa, 对蛋白质的结构和功能不会产生任何影 响,即同义突变 错义突变:突变的密码子编码不同的 aa。 如果错误的 aa 与原来的 aa 属于同种性
发生移码 缺失大片段 DNA(十几到几千个碱基)
质——中性突变;否则可引起分子病如镰刀型贫血和 cystic fibrosis。 无义突变(nonsense mutations):突变的密码子变为终止密码子,如 TGC(Cys 突变成 TGA。分为琥珀型(amber, TAG)、赭石型(ocher,TAA)和乳白型(opal, TGA); 或者相反, 使突变 mRNA 翻译发生通读, 肽链延长, 称加长突变或通读突变(enlongation or read-through mutation), 可能改变多肽性质如影响其稳定性 2、移码突变或移框突变:指在一个蛋白质基因的编码区发生一个或多个核苷酸(非 3 的整数倍)的缺失或插入。 3、隐性突变和显性突变(了解) 隐性:突变仅导致一种蛋白质没有活性(loss of function),产生隐性性状。 显性:突变产生的蛋白质对细胞有毒,这种毒性不能被另外一条染色体上正常基因表达出来的正常的蛋白质所抵消 或中和。 4、回复突变 (了解) 正向突变:突变方向是从野生型向突变型;回复突变,在起始突变位点上发生第二次突变,致使突变从突变表 现型向野生型发生。 校正突变或抑制突变或称假回复突变,指发生在完全不同的另外一个位点,但能够中和起始突变位点(和或抵 消起始突变)的第二次突变,分为基因内校正和基因间校正。 突变的原因 (作业中题) 主要分为自发突变和诱发突变两种 一、自发突变 A、自发点突变: (1) DNA 复制过程的错配 (2)自发的化学变化:脱嘌呤、脱氨(基)作用、氧化作用损伤碱基 (3).氧化损伤 过氧化物原子团(O2-),(H2O2),(-OH)等需氧代谢的副产物是活性氧化剂,可导致 DNA 的氧化损 伤; B、自发的移码突变 (1)复制打滑 当 DNA pol 拷贝到一些具有短重复序列区域(如微卫星序列)的时候,子链和母链之间易发生错配而形成突环 结构。当突环出现在母链,复制向前打滑,导致缺失突变,反之为插入突变。 (2)转座作用 ,又称为异常重组,因为它不需要序列间具有同源性,也不是位点特异性的。被转座子插入的基因 一般会失活,在两个转座子拷贝之间的基因可以通过它们之间的重组而被切除。宿主 DNA 序列的颠倒或其他方式 的重排液可能发生。(补充:转座子或可转座元件是一些比较短 DNA 序列,可以转移进细胞基因组的几乎任何位 置。) 二、 诱发突变 (一) 放射线:紫外线、X-射线、γ 射线、宇宙射线 (二) 化学物质 碱基类似物如:5-溴尿嘧啶(5-bromouracil,5-BU)、氨基嘌呤(2-aminopurine 2-AP)、迭氮胸苷(AZT, azidothymidine) 等 碱基的修饰剂如:亚硝酸(Nitrous acid, NA)、羟胺(HA)、烷化剂(氮芥和硫芥等),它们的作用是使碱基烷基化 DNA 嵌入剂(扁平环状分子)如:原黄素(proflavin)、吖啶橙(acridine orange)溴化 3,8-二氨基-5-乙基-6-苯 基菲啶(ethidium bromide)、ICR 的复合物等
11
第六章
重组
重组:发生在 DNA 分子内或 DNA 分子间核 苷酸序列的交换、重排和转移现象,已有遗传 物质的重新组合。主要包括 3 种形式:同源重 组(普遍性重组)、位点特异性重组、转座。 一、同源重组:在两个 DNA 分子的同源序列 之间直接进行交换的一种重组形式。 不依赖于 序列的特异性,只依赖于序列同源性。 同源重组发生的条件 重组的交换序列具有同源性 两个双链 DNA 分子之间 相互靠近,并互补 配对 特定的重组酶(recombinase)的催化 形成异源双链(heteroduplex) 发生联会(synapsis) 同源重组的分子机制:Holliday 模型(作业 中题图表并述): 2 个同源的 DNA 分子相互靠近 → 2个
(6) (5) 5’ A 3’ 3’ 5’ a (4) (3) (2) (1) 5’ A 3’ 3’ 5’ a 5’ A 3’ 3’ 5’ a B 3’ 5’ 5’ b 3’ 联会 B 3’ 5’ 5’ b 3’ 酶切 3’ 5’ 5’ b 3’ 游离端移动联会 B 3’ (10) 5’ 5’ b 3’ 游离端交叉连接 B 3’ 5’ 5’ b 3 形成单链交叉 3’ 5’ 5 b 3’ 分支点移动 B A B B (8) A B 两臂旋转 b a (9) A B
5’ A 3’ 3’ 5’ a 5’ A 3’ 3’ 5’ a
b a
A B
A B
b a (11) A B A
b a b
a 5’ A 3’ 3’ 5’ a
b
a
B
DNA 分子各有 1 条链在相同的位置被特异的 内切酶切开→被切开的链交叉并与同源的链 连接,形成 χ 状 Holliday 连接→分叉的迁移 → Holliday 连接的拆分 参与同源重组的重要蛋白质和酶(作业中题) 1、 RecA 蛋白:同源重组中最重要蛋白质, 具有单体和多聚体两种形式, 单体具有两个活 性位点,分别与单链和双链 DNA 结合。
(7)
A
B
A
b
a
b
a
B
a
b
图 23-8 Holiday 重组模型。
主要功能为: (1)促进 2 个 DNA 分子之间的链交换; (2)参与 SOS 反应—作为共蛋白酶促进 LexA 和 UmuD 的自水解。 重组中主要作用为促进同源序列配对和链的交换,分为 3 个阶段: 联会前阶段:与单链 DNA 分子结合,形成蛋白-DNA 丝状复合物; 联会阶段:与双链 DNA 分子结合,形成三链 DNA 中间体,随后单链 DNA 侵入双链 DNA,寻找同源序列; 链交换和分支迁移阶段: RecA 包被的单链 DNA 从 5?-3?取代双链 DNA 分子中的同源老链形成异源双链,并发生分 叉迁移。 2、 RecBCD 蛋白:参与同源重组,功能:产生 3’-单链末端,为链入侵作准备。 RecBCD 复合体具有 (1)强的核酸酶(包括外切核酸酶 V、 核酸内切酶和单链 DNA 外切酶); (2)解旋酶; (3)ATPase。 RecBCD 在 Chi 位点产生单链 3?游离未端。 RecBCD 介导的解旋和剪切产生 3'游离末端,用来起始异源双链的连接。 3 、RuvA, RuvB, RuvC 蛋白: RuvA: 识别 Holliday 连接,协助 RuvB 催化分叉的迁移; RuvB: 具 ATPase 活性的解链酶,催化重组中分叉的迁移; RuvC: 特殊的核酸内切酶,促进 Holliday 连接的分离。 E.coli 同源重组途径 1 RecBCD 途径:最主要的重组途径。 参与成分:RecBCD 及 RecA, SSB, RuvABC, DNA 聚合酶 I,连接酶和旋转酶,以及 χ 序列
12
2 RecF 途径和 RecE 途径 RecF 途径:质粒之间进行重组的途径 二、位点特异性重组:发生在 DNA 特异性位点上的重组,参与重组的特异性位点需要专门的蛋白质识别和结合。 发生在 2 个 DNA 分子间或 1 个 DNA 分子内部 功能: (1) 调节噬菌体 DNA 与宿主染色体 DNA 的整合(需要整合酶和整合宿主因子的参与) (2) 调节特定基因的表达 (3) 调节胚胎发育期间程序性的 DNA 重排(如脊椎动物抗体和 T 细胞受体基因)。 (4) 抗体的多样性 三、转座重组:不需要序列间的同源性、也没有位点的特异性,是随机的 转座子:也叫跳跃基因,是指能够在一个 DNA 分子内部或两个以上 DNA 分子之间移动的 DNA 片段。在细菌中指在 质粒和染色体间或质粒和质粒之间移动的 DNA 片段。是 DNA 重组的一种形式。细的转位因子包括插入序列,转座 子及可转座的噬菌体。 转座的遗传学效应(作业中题): (1) 引起插入突变:如各种 IS、Tn 转座子 (2) 产生新的基因:如抗药性基因 (3) 产生染色体畸变:DNA 的缺失与倒位、重复等 (4) 引起生物的进化:人、小鼠和水稻基因组约有 40%的序列由转座子衍生而来,低等真核生物和细菌内比例 较小,约占 1%-5%。 原核生物的转座子(作业中题): 第一类转座子:IS,最简单的转座元件,细菌染色体、质粒和某些噬菌体的正常组分。特征如下: (1) 长度小(700-1800bp); (2) 两端通常含有 10-40bp 的 IR(末端反向重复序列)序列; (3) 内部一般只有一个基因,表达产物是专门催化转位反应的转座酶,无抗性基因; (4) 通过剪切和插入的方式进行转座,导致靶位点序列重复; (5) 少数(IS91)没有明显的 IR 序列,通过滚环复制和插入的方式进行转座。 IS(转座子或插入序列)能够从 DNA 的一位点插入到另一个位点,导致靶位点基因以及和靶位点基因在同一个操 纵子内、但位于靶位点基因下游的基因表达受阻,此现象称为极性效应(polar effect)或极性突变。 第二类转座子:复杂型转座子(complex transposon),特征有 (1) 较长(2.5kb-20kb); (2) 两侧有 35-40bp 的 IR 序列; (3) 内部不止一个结构基因:通常包括 tnpA—转座酶, tnpB—解离酶, 抗性基因(一个或几个) 转座以后导致约 5bp 长的靶位点序列发生重复,结果导致在转座子两侧产生直接重复序列。 第三类:复合型转座子(composite transposon) (1)由 2 个 IS 和一段带有抗生素抗性(如新霉素磷酸转移酶导致新霉素失活)或其他毒性抗性的间插序列组合而 成,其中的 2 个 IS 位于转座子的两侧,具有相同或相反的方向。 (2) 每一个 IS 具有典型的第一类转座子的特征, 可独立转位, 也可与间插序列一道作为一个整体进行集体转移。 第四类:Mu 噬菌体(Bacteriophage Mu) (1)在溶源期,Mu DNA 通过转座随机整合至宿主 DNA 中。复制时,Mu 通过复制型转位随机插入到宿主 DNA 的其他区域,很容易诱发宿主的各种突变。 (2)作为转座子,Mu DNA 为 38kb 线性双链,两侧缺乏 IR 序列,20 多个基因中 A 和 B 与转座有关,其中 A 编码转座酶。 真核生物的转座子 与原核转座子的差别: (1) 真核转座子在转座过程中的剪切和插入是分开进行的
13
(2) 真核转座子的复制很多需要经过逆转录即 RNA 中间物来进行 复制型 DNA 转座子: 转座子序列复制一份并转移到新的位置,转位前后,原位置上的拷贝仍存在。(以滚环复制 的方式进行复制,然后再插入到新的位点。拟南芥和线虫基因组约 2%属于此类) 保留型 DNA 转座子: 转座时原有的拷贝被原封不动地转移保留到新的位点。真核生物绝大多数。 ( 包括玉米的 As-Ds 系统、果蝇的 P 元件、“水手”元件、水稻和线虫的微型反向转座元件) 逆转录转座子 LTR 逆转座子: 两端含有 LTR 序列,果蝇 Copia 元件和酵母 Ty 元件。 非 LTR 逆转座子:无 LTR,有一端含 1 小段重复序列(通常为 polyA):LINE 和 SINE。 第七章 转录 转录特点 (1)转录发生在 DNA 分子上某些特定的区域; (2)以四种核糖核苷三磷酸(NTP)为底物,并需要 Mg2+的激活; (3)以 DNA 为模板,需 DNA 解链,无需引物的存在能单独起始链的合成; (4)按 5′-3′方向合成; (5)第一个引入的 NTP 通常是嘌呤核苷酸(占 90%左右),且以三磷酸形式存在; (6)转录具有高度的忠实性,RNA 的序列和模板是互补的; (7)转录受到严格调控的,调控的位点主要发生在转录的起始阶段。 模板链:将作为转录模板的 DNA 单链称为模板链或反义链或 Watson 链, 有义链:非模板链称为有义链或编码链或 Crick 链。 RNA 合成和 DNA 复制的区别 (1)转录时只有一条 DNA 链为模板,而复制时两条链都可作为模板; (2)DNA-RNA 杂合双链不稳定,RNA 合成后释放,而 DNA 复制叉形成后一直打开,新链和母链形成子链; (3)RNA 合成不需引物,而 DNA 复制需引物; (4) 转录的底物是 NTP,复制的底物是 dNTP; (5)聚合酶系不同。 DNA 转录的酶学:即依赖 DNA 的 RNA 聚合酶( DNA dependent RNA polymerase, DDRP ) 其特点有: ? 以 DNA 为模板,转录的方式是不对称的。 ? RNA 链的延长方向是 5’→3’的连续合成。 ? 需要 Mg2+或 Mn2+离子。 ? 缺乏外切酶活性,所以没有校正功能。 ? 不需要引物,能直接催化 RNA 的从头合成。 ? RNA pol 启动转录需要识别启动子 ? RNA pol 反应的速度低,平均速率只 50nt/s。 抑制剂: 真细菌的 RNA pol 的特异性抑制:利福霉素(rifamycin)和利链霉素(streptolydigin) /利福平和利敌链菌素 对象:β 亚基,前者抑制转录的起始,阻止第三个或第四个核苷酸的参入;后者与聚合酶结合,抑制延伸。 不抑制真核细胞核 RNA pol,抑制线粒体或叶绿体体的 RNA pol。 真核生物的三种 RNA 聚合酶特点 (作业中题) RNA 聚合酶Ⅰ转录大部分的 rRNA 的基因,存在于核仁中,并对α -鹅膏蕈不敏感; RNA 聚合酶Ⅱ转录所有的编码基因和一些 SnRNA 基因,存在于核质中,对α -鹅膏蕈非常敏感; RNA 聚合酶Ⅲ转录 tRNA,5S rRNA,某些 U6SnRNAs 的基因以及一些小分子 RNA,也存在于核质中,对α -鹅膏蕈 中度敏感;详见下图:
14
表 12-2 真核生物的三种 RNA 聚合酶的特点 RNA Pol 位置 Pol Ⅰ Pol Ⅱ Pol Ⅲ 核仁 核质 核质 产物 28s,18s,5.8s rRNAs hnRNA,mRNA,某些 SnRNA tRNA,5S rRNA,某些 SnRNAs 相对活性 50~70% 20~40% ~10% 对α -鹅膏蕈的敏感性 不敏感 高度敏感 中等敏感
羧基末端结构域(CTD):,真核生物 RNA 聚合酶Ⅱ的羧基端含有一段 7 氨基酸序列,一致序列富含羟基 aa 残基 (YSPTSPT),这一七肽序列被称为羧基末端结构域即 CTD,对酶活性是必需的。 RNA pol 的多种功能 (真核细胞的 RNA pol 不能直接识别启动子, 必须借助于转录因子(TF)才能结合到启动子。 ) (1) 识别 DNA 双链上的启动子; (2) 使 DNA 双链在启动子处解旋成单链; (3) 通过阅读启动子序列,RNA pol 确定它自己的转录方向和模板链。 (4) 最后当它达到终止子时,停止转录。 原核生物的转录 3 个阶段:起始、延伸和终止。 启动子(promoter)是基因转录精确和特异性启动所必需的 DNA 序列。 结构和转录起始: (1)结构典型, 都含识别(R),结合(B)和起始(I)位点; (2)序列保守;(3)位置和距离都比较恒定;(4)直接和多聚酶 相结合;(5)常和操纵子相邻;(6)一般位于基因的 5′端;(7)决定转录的启动和方向。 转录起始位点(I)(作业中题原核启动子的结构特征) ? 转录开始时模板上的第一个碱基,在原核中常为 A 或 G,而且位置固定 ? 以编码链为准,启动子碱基的位置以转录的起始点(start point)为参照,转录起始点定为+1,上游(5?端)的序 列为负数,下游(3?端)的碱基为正数,没有 0。 (1)-10 序列:是由 Pribnow 和 Schaller(1975)发现,故也称为 Pribnow 框盒(Pribnow box)。保守序列为 TATAAT 位于-10bp 左右,A.T 较丰富,易于解链。其功能是: (1) 与 RNA pol 紧密结合; (2) 形成开放启动复合体; (3) 使 RNA pol 定向转录。 -10 序列对转录的效率影响 TATAAT→AATAAT,转录效率下降, 称为下降突变(down mutation)。 TATGTT→TATATT,转录效率上升,为上升突变(up mutation)。 以上突变为何会影响转录效率: 前者可能由于 T-A 的堆集能要小于 A-T 的堆积能,后者可能是由于堆集能降低和氢键的减少, (2)-35 序列:又称为 Sextama 盒(Sextama box),其保守序列为(TTGACA) 其功能是: (1) 为 RNA pol 的识别位点。ζ 亚基识别-35 序列,为转录选择模板 (2)-35 和-10 序列的距离是稳定的,一般为 17bp± 1bp, 此与 RNA pol 的结构有关。 增强元件:某些转录活性超强的基因如 rRNA 基因除-10 和-35 区外,在-40 和-60 间的区域还有一富含 AT(5?-AAAATTATTTT-3?)的增强元件(up element)的启动子序列,可使转录活性提高 30 倍。 转录的起始(限速) 1.全酶与模板的 DNA 接触,生成非专一的,不稳定的复合物在模板上移动; 2. 起始识别:全酶与-35 序列结合,产生封闭的酶-启动子二元复合物(closed binary complex); 3.全酶紧密地结合在-10 序列处,模板 DNA 局部变性,形成开放的启动子二元复合体,即起始转录泡;
15
4.第一个磷酸二酯键的形成。 前两个与模板链互补的 NTP 经聚合酶催化第一个 NTP 的 3?-OH 亲核进攻第二个 NTP 的 5?-α-P 而形成第一个磷酸二酯键,第一个 RNA-DNA-RNA pol 三元复合物(ternary complex)形成。 5.起始阶段的无效转录:开放复合物内的 RNA pol 往往会重复催化短 RNA 分子的合成并释放它们,这样的合成称 为“无效”合成(abortive synthesis)。一般长度为 6nt 左右。 6.启动子清空(promoter clearance):RNA pol 离开启动子以实现转录从起始进入延伸的过程。NusA 作为延伸因子代 替ζ 因子与核心酶结合。 RNA 核心酶和全酶在 DNA 上的分布: (1) ζ 和 1/3 的 RNA pol 结合成全酶,或在非特异位点的松散复合体中,或在启动子中的二元复合体中; (2) 其中半数的核心酶从事转录; (3) 余下的核心酶大量存在于闭合松散复合体中; (4)估计数量很少的全酶是游离的。 转录的终止的两种方式: 1 不需要 ρ(rho factor)因子的终止:需要 RNA 转录物 3’-端终止子(terminator)的序列,其两个特征:一是依赖于 位于 RNA 转录物 3?-端的一串 U 序列(导致 RNA 和反义 DNA 练得弱结合),另一是需要位于紧靠 U 序列上游的 一个富含 GC 碱基对(利于碱基配对的稳定性)的发夹结构。 终止子(terminator) ? ? 强终止子-内部终止子(intrinsic terminators) 弱终止子 -需要 ρ 因子,又称为 ρ 依赖性终止子(Rho-dependent terminator)
强终止子的结构特点 ? (1) 有回文结构存在; ? ? (2)茎的区域富内含 G-C; (3) 强终止子 3′端至少含有 3 个 U;
以上结构特点在终止中起何作用?发卡结构茎部富含 GC 碱基对,使其有很高的稳定性,从而使聚合酶在此停顿。 发卡之后的 U 碱基导致 RNA 和反义 DNA 练得弱结合,这有利于 RNA 链的解离,从而终止反应。 2 需要 ρ 因子的转录终止 ρ 因子: 60kDa, 同源六聚体蛋白, 具有 NTP 酶和解链酶活性, 能分别水解各种 NTP 和催化 RNA/DNA 和 RNA/RNA 双螺旋的解链,促使新生的 RNA 链从三元转录复合物中解离出来,从而终止转录。 作用模型: 在 RNA 合成起始以后, ρ 因子识别转录物 5’-端一段特殊的碱基序列并与之结合,靠 ATP 水解产生的 能量,沿着 5?→3?方向朝 RNA 聚合酶移动,到达 RNA 的 3?-OH 端后取代了暂停在终止位点上的 RNA 聚合酶,随 后, ρ 因子通过其解链酶的活性解开转录泡上的 RNA/DNA 杂交双螺旋,并从模板和酶上释放 RNA,完成转录过 程。需要消耗能量,而 ρ 因子具有终止转录和 NTP 酶两种功能。 真核生物的转录 真核生物的转录和原核转录的不同点: (1) 染色质和核小体结构对转录有深刻的影响 (2) 原核只有一种 RNA 聚合酶,亚基少,直接识别启动子并具有解链酶活性,而真核细胞有三种聚合酶 (3) 启动子的结构特点不同,真核有三种不同的启动子和有关的元件 (4) 启动子以外的序列参与调节基因的转录,顺式 作用元件(cis-acting element) (5) 真核的转录有很多蛋白质因子即转录因子的介入 (6) 转录和翻译不存在偶联关系 (7) 转录的产物多为单顺反子,而原核基因的转录产物大多数为多顺反子 真核生物的启动子包括(作业中题) :RNA 聚合酶Ⅰ启动子、RNA 聚合酶Ⅱ启动子和 RNA 聚合酶Ⅲ启动子。启动子Ⅱ最为复杂,它和原核的启动子有 很多不同: (1)有多种元件:TATA 框,GC 框,CAAT 框,OCT 框等;
16
(2)结构不恒定; (3)它们的位置、序列、距离和方向都不完全相同; (4)有的有远距离的调控元件存在,如增强子; (5)这些元件常常起到控制转录效率和选择起始位点的作用; (6)需多种转录因子介入。 Ⅱ类基因的启动子和调控区 1、起始子 起始点一般没有同源序列,mRNA 的第一个碱基倾向 A,另一侧翼由 Py (C)组成称为起始子(initiator, Inr)。提 供 RNA pol Ⅱ识别。无论 TATA 是否存在,Inr 对启动子的强度和起始位点的选择都是重要的 。 2、TATA 框(和 Inr 属于招募和定位元件,两者共同决定转录的起点) TATA 框又称 Hogness 框,其一致序列是:TATAAAA 常在-25 ~-30 bp 左右,相当于原核的-10 序列。 其作用是: (1) 选择正确的转录起始位点,保证精确起始, 故也称为选择子(selector)。 (2) 影响转录的速率。 3、增强子(enhancer):又称远上游序列,是一种能大幅度增强基因转录效率的顺式作用元件。 其特点是: ① 具有远距离效应。② 无方向性。③ 顺式调节。④ 无物种和基因的特异性,具组织的特异性。 ⑥ 有相位性,其作用和 DNA 的构象有关。⑦ 有的增强子可以对外部信号产生反应。 RNA polⅠ启动子 核心启动子(core promoter)或核心元件(core element),位于-31 到+6,负责转录的起始。 上游控制元件(UCE),从 -187 延伸到-107,可增加核心元件的转录起始的效率。核心启动子与 UCE 序列高度 同源,约 85%序列相同。 转录因子:一种是 UBF(UCE binding factor),识别 UCE 和核心启动子富含 GC 序列后,与启动子结合。另一种为 SL1(selectivivty factor),由多亚基组成,分为 TBP(TATA 盒结合蛋白)和 TAF(TBP 相关因子)两种成分。 RNA pol III 启动子:转录结构较稳定的小分子 RNA,如 tRNA、5S rRNA、7SL RNA、Small nucleolar RNA(SnoRNA)、SnRNA 和某些病毒 mRNA 等。 外部启动子(external):位于基因的上游,含 TATA 盒、近序列元件(proximal sequence element, PSE)和远端序列 元件(distal sequence element, DSE),如 7SL RNA 和 U6 snRNA 等。 内部启动子(internal):位于基因内部,如 tRNA、5S rRNA 和 Adenovirus 的 VA RNA 等。 转录因子:TFIII 有三种,A、B 和 C。 转录校对 首先 RNA pol 本身的高度选择性 转录校对两种方式:1、焦磷酸解编辑(pyrophosphorolytic editing): 使用 pol 的活性中心,以逆反应形式重新参入 焦磷酸,去除错误插入的核苷酸。 2、水解编辑(hydrolytic editing): pol 倒退若干个 nt,然后通过特殊的蛋白质(细菌为 GreA 和 GreB; TFII S 刺激 RNA pol II 的剪切活性)切除 3?-端几个 nt,包括错配的核苷酸。 RNA 复制: RNA 为模板合成 RNA 的过程, 以 由依赖于 RNA 的 RNA pol(RNA-dependent RNA polymerase, RdRP) 催化,RdRP 又名 RNA 复制酶(replicase)。一般由病毒基因组编码,也可能需要宿主细胞编码的辅助蛋白。 RNA 复制特征: 1、复制的方向始终为 5?-3? 2、绝大多数在模板的一端从头启动合成 3、属于易错、高突变的 RNA 合成,错误率比 DNA 聚合酶高约 104 倍。 4、对放线菌素 D 作用一般不敏感,但对核糖核酸酶敏感 5、复制绝大多数发生在宿主细胞的细胞质,少数在细胞核。
17
第八章
翻译
一、参与翻译的主要生物大分子的结构和功能(作业中题) (一)、核糖体(ribosome) 功能:生物细胞内,合成蛋白质的“工厂”。 组成:由蛋白质和 rRNA,一种亚细胞颗粒。 位置:直接或间接与细胞骨架结构有关联或与内质网膜结构相连。细菌核糖体大都通过与 mRNA 相互作用,被 固定在核基因组上。 (1) 核糖体的结构组成 (如右图所示) (作 业中题) 原核: 约 2/3 的 RNA 及 1/3 的蛋白质 真核: RNA 占 3/5,蛋白质占 2/5 解离为两个亚基: 其中小亚基只有一种 rRNA,原核为 16S,真核为 18S。 大肠杆菌核糖体小亚基由 21 种蛋白质组 成,分别用 S1……S21 表示,大亚基由 34 种蛋白质组成,分别用 L1……L34 表 示。 真核生物细胞核糖体大亚基含有约 50 种 蛋白质,小亚基有 33 种蛋白质。 (2)核糖体的功能位点(作业中题) 1)A 部位(acceptor site, A site), 即氨酰 tRNA 结合部位,也称为受体部位 2)P 部位(peptidal site, P site),即肽酰 tRNA 结合部位,也称为供体部位(donor site) 3) E 部位(exit site, E site), 即空载 tRNA 在离开核糖体之前与核糖体临时结合的部位 4)肽酰转移酶(peptidyl transferase)活性部位,该部位负责催化肽键的形成 5)mRNA 结合部位 6)多肽链离开通道(exit channel) 7) 一些可溶性蛋白质因子(起始因子、延伸因子和终止因子)的结合部位 小亚基上有 mRNA 结合位点,负责对序列特异的识别过程,如起始位点的识别和密码子与反密码子的相互作用。 大亚基负责氨基酸及 tRNA 携带的功能,肽键的形成、AA- tRNA、肽酰- tRNA 的结合等。A 位、P 位、肽酰转移酶 活性部位等主要在大亚基上。 (3)核糖体的组装和循环 ? ? ? 核糖体在体内及体外都可解离为亚基或结合成 70S/80S 颗粒。翻译的起始阶段需要游离的亚基,随后才 结合成 70S/80S 颗粒,继续翻译进程。 体外体系,核糖体的解离或结合取决于 Mg2+浓度。在 E.coli 内,Mg2+浓度在 10-3 M 以下时,70S 解离, 浓度达 10-2M 时则形成稳定的 70S 颗粒。 细胞中大多数核糖体处于非活性的稳定状态,单独存在,只有少数与 mRNA 一起形成多聚核糖体。 (二)、mRNA:mRNA 是翻译的模板,直接指导蛋白质的合成。 是至少含有一个由起始密码子开始、以终止密码子结束的一段由连续的核苷酸序列构成的开放阅读框(ORF)。 mRNA 5?-端和 3?-端通常含有非编码序列(non-coding sequence, NCS)或称非翻译区(UTR)。 原核 mRNA 在第一个结构基因的 5?-端有核糖体结合位点(ribosome binding site, RBS),RBS 含有富含嘌呤的 SD 序列,可被核糖体识别、结合。 (三)、tRNA tRNA 的结构
18
(1)tRNA 一级结构的特征 a、一类小分子 RNA,长度通常在 73~93 nt, 其中 22 个碱基是恒定的 b、所有的 tRNA 在 3?端具有 CAA 序列,氨基酸通过酯键连接在末端 A 的 OH 基上 c、tRNA 含有大量修饰碱基,已发现上百种不同的共价修饰形式,如二氢尿嘧啶(dihydrouridine, D)和假尿苷(,Ψ)。 (2)三叶草型的二级结构(clover):4 个茎和 3 个环组成。 a、受体臂(acceptor arm)或 aa 臂:5?端和 3?端配对(常为 7bp)形成的茎区。在 3?端永远是 4 个碱基(XCCA)的单 链区,在其末端有 2?-OH 或 3?-OH,是被氨基酰化位点。此臂负责携带特异的氨基酸。 b、TψC 常由 5bp 的茎和 7nt 的环组成,因含有高度保守的 TψC 序列而得名。此臂负责和核糖体上的 rRNA 识别 结合; c、反密码子臂(anticodon arm):常由 5bp 的茎区和 7nt 的环区组成,它负责对密码子的识别与配对。 d、D 环 (D arm)的茎区长度常为 4bp,也称双氢尿嘧啶环。负责和氨酰-tRNA 聚合酶结合; e、额外环(extra arm):可变性大,又称可变环(the variable loop),从 4 nt 到 21 nt 不等,其功能是在 tRNA 的 L 型三维结构中负责连接两个区域(D 环-反密码子环和 TψC-受体臂)。 (3)tRNA 三级结构的特征(倒 L 型) (1)D 环和 TψC 环中的一些核苷酸形成氢键,使分子形成 L 形, 并使结构稳定。 (2)倒 L 型结构中,两段 RNA 双螺旋之间呈垂直关系,其中的一段由 TψC 茎和氨基酸受体茎并列而成,另外一段 双螺旋由 D 茎和反密码子茎并列而成 (3)氨基酸受体臂位于 L 型的一侧,距反密码子环在空间上分开 (4)tRNA 的功能 1、将氨基酸运载到核糖体 2、通过反密码子与 mRNA 上的密码子之间的相互作用对遗传密码进行解码。 tRNA 的种类 (1)起始 tRNA 和延伸 tRNA 能特异地识别 mRNA 模板上起始密码子的 tRNA 叫起始 tRNA,其他 tRNA 统称为延伸 tRNA。原核生物起始 tRNA 携带甲酰甲硫氨酸(formylmethonine, fMet),真核生物起始 tRNA 携带甲硫氨酸(Met)。 (2)同工受体 tRNA (isoaccepting tRNA) 携带同一种氨基酸的几种不同 tRNA 分子。 同工受体 tRNA 既要有不同的反密码子以识别该氨基酸的各种同义 密码,又要有某种结构上的共同性,能被 AA- tRNA 合成酶识别。 (3)校正 tRNA 校正 tRNA 分为无义突变及错义突变校正 tRNA 。 在蛋白质的结构基因中, 一个核苷酸的改变可能使代表某个氨基酸的密码子变成终止密码子 (UAG、 UGA、 UAA) , 使蛋白质合成提前终止,合成无功能的或无意义的多肽,这种突变就称为无义突变。而无义突变的校正 tRNA 可通 过改变反密码子区以校正无义突变。 (四)、氨酰- tRNA 合成酶 AA- tRNA 合成酶(aminoacyl-tRNA synthetase, aaRS)是一类催化氨基酸与 tRNA 结合的特异性酶,其反应式如 下: AA+tRNA+ATP→AA-tRNA-AMP+PPi 包括两步反应: 1. 氨基酸活化生成酶-氨基酰腺苷酸复合物: AA+ATP+酶(E)→E-AA-AMP+PPi 2. 氨酰基转移到 tRNA 3?末端腺苷酸残基上,与其 2?或 3?-羟基结合: E-AA-AMP+ tRNA→AA- tRNA +E+AMP 多肽链合成的保真性主要决定于 AA- tRNA 合成酶(aaRS)是否能使氨基酸与对应的 tRNA 相结合。 aaRS 既识别 tRNA,又识别 aa,它对两者均具有高度的专一性。不同的 tRNA 有不同碱基组成和空间结构,可被 tRNA 合成酶所识别。
19
然而对于同一 aa 的 tRNA 可能存在几种不同的同工受体 tRNA,如何识别? 通过 tRNA 序列突变实验发现,决定一种 aaRS 识别正确的 tRNA 的主要因素是 tRNA 分子上由几个核苷酸甚至一 个核苷酸组成的元件,这些元件常被称为 tRNA 的个性(identity),又称第二套遗传密码(the second genetic code)。 aaRS 如何选择正确的 AA? (1)通过酶活性中心优先结合正确的同源 aa,体积比同源 aa 大的被完全排除在活性中心外; (2) 通过酶的校对中心选择性编辑错误的非同源 aa。 能进入活性中心的小 aa 在错误形成误载的氨酰-AMP 或氨酰 -tRNA 后,被送入校对中心(editing site),错误的 aa 被水解,离开酶分子。这两种机制结合称之为“双筛”机制。 (五)、辅助因子 翻译的每一阶段均需特殊的可溶性蛋白质因子参与,包括: ? 起始因子(iniation factor, IF):参与肽链合成起始 ? 延伸因子(elongation factor, EF): ? 释放因子(release factor, RF): ? 核糖体循环因子(ribosome recycling factor, RRF): 二、翻译的一般性质(作业中题,翻译的基本特征) 自然界四类体内翻译系统:原核生物、真核生物细胞质、叶绿体和线粒体翻译系统。 (1)翻译的四个阶段:活化、起始、延伸、终止和释放 (2)翻译具有方向性(极性,polarity):一是阅读模板时的方向性,mRNA 从 5?-至 3?-端;二是多肽链延伸的方向 总是从 N-端→C-端。 (3)遗传密码——三联体 mRNA 上每 3 个核苷酸翻译成蛋白质多肽链上的一个 aa,这种由 3 个核苷酸决定 1 个 aa 的编码形式被称为三 联体密码(triplet codon)。 翻译时从起始密码子 AUG 开始,沿 mRNA5?→3?的方向连续阅读直到终止密码子,生成一条具有特定序列的多 肽链。 遗传密码的性质 ①遗传密码是三联体密码。②遗传密码无逗号。 ③遗传密码是不重迭的。④遗传密码具有通用性。 ⑤遗传密码具有简并性(degeneracy synonyms)。 ⑥密码子有起始密码子和终止密码子。⑦反密码子中的“ 摆动”(wobble)。 起始密码子:AUG,GUG,更少 UUG 终止密码子:UAA 赭石密码(E.coli 使用最多);UGA 琥珀密码;UAG 乳白(蛋白石)密码 摆动假说(wobble hypothesis)(作业中题)是由 Crick.F(1966 年)提出的。即当 tRNA 的反密码子与 mRNA 的 密码子配对时前两对严格遵守碱基互补配对法则,但第三对碱基有一定的自由度可以“摆动”。 翻译的机制: 翻译各个阶段的主要成分简表
阶段 1 aa 的活 化 2 肽链的 起始
20
必需组分 20 种 aa,20 种氨酰-tRNA 合成酶 20 种或更多的 tRNA,ATP, Mg2+ mRNA,N-甲酰甲硫氨酰-tRNA mRNA 上的起始密码子(AUG) 核糖体小亚基、大亚基 2+ GTP,Mg ,起始因子(IF-1,IF-2,IF-3)
现代分子生物学_复习笔记完整版.doc
现代分子生物学 复习提纲 第一章绪论 第一节分子生物学的基本含义及主要研究内容 1 分子生物学Molecular Biology的基本含义 ?广义的分子生物学:以核酸和蛋白质等生物大分子的结构及其在遗传信息和细胞信息传递中的作用为研究 对象,从分子水平阐明生命现象和生物学规律。 ?狭义的分子生物学:偏重于核酸(基因)的分子生物学,主要研究基因或DNA的复制、转录、表达和调控 等过程,也涉及与这些过程相关的蛋白质和酶的结构与功能的研究。 1.1 分子生物学的三大原则 1) 构成生物大分子的单体是相同的 2) 生物遗传信息表达的中心法则相同 3) 生物大分子单体的排列(核苷酸、氨基酸)的不同 1.3 分子生物学的研究内容 ●DNA重组技术(基因工程) ●基因的表达调控 ●生物大分子的结构和功能研究(结构分子生物学) ●基因组、功能基因组与生物信息学研究 第二节分子生物学发展简史 1 准备和酝酿阶段 ?时间:19世纪后期到20世纪50年代初。 ?确定了生物遗传的物质基础是DNA。 DNA是遗传物质的证明实验一:肺炎双球菌转化实验 DNA是遗传物质的证明实验二:噬菌体感染大肠杆菌实验 RNA也是重要的遗传物质-----烟草花叶病毒的感染和繁殖过程 2 建立和发展阶段 ?1953年Watson和Crick的DNA双螺旋结构模型作为现代分子生物学诞生的里程碑。 ?主要进展包括: ?遗传信息传递中心法则的建立 3 发展阶段 ?基因工程技术作为新的里程碑,标志着人类深入认识生命本质并能动改造生命的新时期开始。 ? 第三节分子生物学与其他学科的关系 思考 ?证明DNA是遗传物质的实验有哪些? ?分子生物学的主要研究内容。 ?列举5~10位获诺贝尔奖的科学家,简要说明其贡献。
现代分子生物学复习题
现代分子生物学复习题
现代分子生物学 一.填空题 1.DNA的物理图谱是DNA分子的限制性内切酶酶解片段的排列顺序。 2.核酶按底物可划分为自体催化、异体催化两种类型。 3.原核生物中有三种起始因子分别是IF-1、 IF-2 和IF-3 。 4.蛋白质的跨膜需要信号肽的引导,蛋白伴侣的作用是辅助肽链折叠成天然构象的蛋白质。 5.真核生物启动子中的元件通常可以分为两种:核心启动子元件和上游启动子元件。 6.分子生物学的研究内容主要包含结构分子生物学、基因表达与调控、DNA重组技术三部分。 7.证明DNA是遗传物质的两个关键性实验是肺炎球菌感染 小鼠、T2噬菌体感染大肠杆菌这两个实验中主要的论点证据是:生物体吸收的外源DNA改变了其遗传潜能。 8.hnRNA与mRNA之间的差别主要有两点: hnRNA在转变为mRNA的过程中经过剪接、 mRNA的5′末端被加上一个m7pGppp帽子,在mRNA3′ 东隅已逝 2 桑榆非晚!
末端多了一个多聚腺苷酸(polyA)尾巴。 9.蛋白质多亚基形式的优点是亚基对DNA的利用来说是一 种经济的方法、可以减少蛋白质合成过程中随机的错误对蛋白质活性的影响、活性能够非常有效和迅速地被打开和被关闭。 10.质粒DNA具有三种不同的构型分别是: SC构型、 oc 构型、 L构型。在电泳中最前面的是SC构型。 11.哺乳类RNA聚合酶Ⅱ启动子中常见的元件TATA、GC、 CAAT所对应的反式作用蛋白因子分别是TFIID 、SP-1 和 CTF/NF1 。 12.与DNA结合的转录因子大多以二聚体形式起作用,转 录因子与DNA结合的功能域常见有以下几种螺旋-转角-螺旋、锌指模体、碱性-亮氨酸拉链模体。 13.转基因动物常用的方法有:逆转录病毒感染法、DNA 显微注射法、胚胎干细胞法。 14.RNA聚合酶Ⅱ的基本转录因子有、TFⅡ-A、TFⅡ-B、 TFII-D、TFⅡ-E他们的结合顺序是: D、A、B、E 。 其中TFII-D的功能是与TATA盒结合。 15.酵母DNA按摩尔计含有32.8%的T,则A为_32.8%_,G 为_17.2%_和C为_17.2%__。 16.操纵子包括_调控基因、调控蛋白结合位点和结构基因。 17.DNA合成仪合成DNA片段时,用的原料是模板DNA 东隅已逝 3 桑榆非晚!
分子生物学复习资料绝对重点
分子生物学复习资料 (第一版) 一名词解释 1 Southern blot / Northern blot—DNA斑迹法 / RNA转移吸印技术。是为了检测待检基因或其表达产物的性质和数量(基因拷贝数)常用的核酸分子杂交技术。二者均属于印迹转移杂交术,所不同的是前者用于检测DNA样品;后者用于检测RNA样品。 2 cis-acting element / trans-acting factor—顺式作用元件 / 反式作用因子。均为真核生物基因中的转录调控序列。顺式作用元件是与结构基因表达调控相关、能被基因调控蛋白特异性识别和结合的特定DNA序列,包括启动子和上游启动子元件、增强子、反应元件和poly (A)加尾信号。反式作用因子是能与顺式作用元件特异性结合、对基因表达的转录起始过程有调控作用的蛋白质因子,如RNA聚合酶、转录因子、转录激活因子、抑制因子。 3VNTR / STR—可变数目串联重复序列 / 短串联重复。均为非编码区的串联重复序列。 前者也叫高度可变的小卫星DNA,重复单位约9~24bp,重复次数变化大,变化高度多态性;后者也叫微卫星DNA,重复单位约2~6 bp,重复次数约10~60次,总长度通常小于150bp 。(参考第7题) 4 viral oncogene / cellular oncogene—病毒癌基因 / 细胞癌基因。病毒癌基因指存在于逆转录病毒中、体外能使细胞转化、体内能导致肿瘤发生的基因;细胞癌基因也叫原癌基因,指存在于细胞内,与病毒癌基因同源的基因序列。正常情况下不激活,与细胞增殖相关,是维持机体正常生命活动所必须的,在进化上高等保守。当原癌基因的结构或调控区发生变异,基因产物增多或活性增强时,使细胞过度增殖,从而形成肿瘤。 5 ORF / UTR—开放阅读框 / 非翻译区。均指在mRNA中的核苷酸序列。前者是特定蛋白质多肽链的序列信息,从起始密码子开始到终止密码子结束,决定蛋白质分子的一级功能;后者是位于前者的5'端上游和3'端下游的、没有编码功能的序列,主要参与翻译起始调控,为前者的多肽链序列信息转变为多肽链所必需。 6 enhancer / silencer—增强子 / 沉默子。均为顺式作用元件。前者是一段含多个作用元件的短DNA序列,可特异性与转录因子结合,增强基因的转录活性,可以位于基因任何位置,通常在转录起始点上游-100到-300个碱基对处;后者是前者内含的负调控序列,结合特异蛋白因子时,对基因转录起阻遏作用。 7 micro-satellite / minisatellite—微卫星DNA / 小卫星DNA 。卫星DNA是出现在非编码区的串联重复序列,特点是有固定重复单位且重复单位首尾相连形成重复序列片段,串联重复单位长短不等,重复次数大小不一。微卫星DNA即STR;小卫星DNA分为高度可变的小卫星DNA(即VNTR)和端粒DNA。(参考第3题) 8 SNP / RFLP—单核苷酸多态性 / 限制性片段长度多态性。前者是指在基因组水平上由单个核苷酸的变异所引起的DNA序列多态性,它是人类遗传变异中最常见的一种,占所
现代分子生物学总结(朱玉贤、最新版)
现代分子生物学总结(朱玉贤、最新版)
一、绪论 两个经典实验 1、肺炎球菌在老鼠体内的毒性实验:先将光滑型致病菌(S型)烧煮杀活性以后、以及活的粗糙型细菌(R型)分别侵染小鼠发现这些细菌自然丧失了治病能力;当他们将经烧煮杀死的S型细菌和活的R型细菌混合再感染小鼠时,实验小鼠每次都死亡。解剖死鼠,发现有大量活的S型细菌。实验表明,死细菌DNA 进行了可遗传的转化,从而导致小鼠死亡。 2、T2噬菌体感染大肠杆菌:当细菌培养基中分别带有35S或32P标记的氨基酸或核苷酸,子代噬菌体就相应含有35S标记的蛋白质或32P标记的核酸。分别用这些噬菌体感染没有放射性标记的细菌,经过1~2个噬菌体DNA 复制周期后进行检测,子代噬菌体中几乎不含带35S标记的蛋白质,但含30%以上的32P 标记。说明在噬菌体传代过程中发挥作用的可能是DNA而不是蛋白质。 基因的概念:基因是产生一条多肽链或功能RNA分子所必需的全部核苷酸序列。
二、染色体与DNA 嘌呤嘧啶 腺嘌呤鸟嘌呤胞嘧啶尿嘧啶胸腺嘧啶 染色体 性质:1、分子结构相对稳定;2、能够自我复制,使亲、子代之间保持连续性;3、能指导蛋白质的合成,从而控制生命过程;4、能产生可遗传的变异。 组蛋白一般特性:1、进化上极端保守,特别是H3、H4;2、无组织特异性;3、肽链上氨基酸分布的不对称性;4、存在较普遍的修饰作用;5、富含赖氨酸的组蛋白H5 非组蛋白:HMG蛋白;DNA结合蛋白;A24非组蛋白
真核生物基因组DNA 真核细胞基因组最大特点是它含有大量的重复序列,而且功能DNA序列大多被不编码蛋白质的非功能蛋白质所隔开。人们把一种生物单倍体基因组DNA的总量称为C值,在真核生物中C 值一般是随着生物进化而增加的,高等生物的C 值一般大于低等动物,但某些两栖类的C值甚至比哺乳动物还大,这就是著名的C值反常现象。真核细胞DNA序列可被分为3类:不重复序列、中度重复序列、高度重复序列。 真核生物基因组的特点:1、真核生物基因组庞大,一般都远大于原核生物的基因组;2、真核基因组存在大量的的重复序列;3、真核基因组的大部分为非编码序列,占整个基因组序列的90%以上,这是真核生物与细菌和病毒之间的最主要的区别;4、真核基因组的转录产物为单顺反之;5、真核基因组是断裂基因,有内含子结构;6、真核基因组存在大量的顺式元件,包括启动子、增强子、沉默子等;7、真核基因组中存在大量的DNA多态性;8、真核基因组具有端粒结构。
分子生物学复习题
1、分子生物学的定义。 从分子水平研究生物大分子的结构与功能从而阐明生命现象本质的科学,主要指遗传信息的传递(复制)、保持(损伤和修复)、基因的表达(转录和翻译)与调控。 2、简述分子生物学的主要研究内容。 a.DNA重组技术(基因工程) (1)可被用于大量生产某些在正常细胞代谢中产量很低的多肽 ; (2)可用于定向改造某些生物的基因组结构 ; (3)可被用来进行基础研究 b.基因的表达调控 在个体生长发育过程中生物遗传信息的表达按一定时序发生变化(时序调节),并随着内外环境的变化而不断加以修正(环境调控)。 c.生物大分子的结构和功能研究(结构分子生物学) 一个生物大分子,无论是核酸、蛋白质或多糖,在发挥生物学功能时,必须具备两个前提: (1)拥有特定的空间结构(三维结构); (2)发挥生物学功能的过程中必定存在着结构和构象的变化。 结构分子生物学就是研究生物大分子特定的空间结构及结构的运动变化与其生物学功能关系的科学。它包括3个主要研究方向: (1) 结构的测定 (2) 结构运动变化规律的探索 (3) 结构与功能相互关系 d.基因组、功能基因组与生物信息学研究 3、谈谈你对分子生物学未来发展的看法? (1)分子生物学的发展揭示了生命本质的高度有序性和一致性,是人类认识论上的重大飞跃。生命活动的一致性,决定了二十一世纪的生物学将是真正的系统生物学,是生物学范围内所有学科在分子水平上的统一。 (2)分子生物学是目前自然学科中进展最迅速、最具活力和生气的领域,也是新世纪的带头学科。
(3)分子生物学是由生物化学、生物物理学、遗传学、微生物学、细胞学、以及信息科学等多学科相互渗透、综合融会而产生并发展起来的,同时也推动这些学科的发展。 (4)分子生物学涉及认识生命的本质,它也就自然广泛的渗透到医学、药学各学科领域中,成为现代医药学重要的基础。 1、DNA双螺旋模型是哪年、由谁提出的?简述其基本内容。 DNA双螺旋模型在1953年由Watson和Crick提出的。 基本内容: (1) 两条反向平行的多核苷酸链围绕同一中心轴相互缠绕,两条链均为右手双螺旋。 (2) 嘌呤与嘧啶碱位于双螺旋的内侧,3′,5′- 磷酸与核糖在外侧,彼此通过磷酸二酯键相连接,形成DNA分子的骨架。 (3) 双螺旋的平均直径为2nm,两个相邻碱基对之间相距的高度即碱基堆积距离 为0.34nm,两个核苷酸之间的夹角为36。。 (4) 两条核苷酸链依靠彼此碱基之间形成的氢键相连系而结合在一起,A与T相配对形成两个氢键,G与C相配对形成3个氢键。 (5) 碱基在一条链上的排列顺序不受任何限制,但根据碱基互补配对原则,当一条多核苷酸的序列被确定后,即可决定另一条互补链的序列。
现代分子生物学课后答案(朱玉贤_第三版)上
第一章绪论 2.写出DNA和RNA的英文全称。 答:脱氧核糖核酸(DNA, Deoxyribonucleic acid),核糖核酸(RNA, Ribonucleic acid)4.早期主要有哪些实验证实DNA是遗传物质?写出这些实验的主要步骤。 答:一,肺炎双球菌感染实验,1,R型菌落粗糙,菌体无多糖荚膜,无毒,注入小鼠体内后,小鼠不死亡。2,S型菌落光滑,菌体有多糖荚膜,有毒,注入到小鼠体内可以使小鼠患病死亡。3,用加热的方法杀死S型细菌后注入到小鼠体内,小鼠不死亡; 二,噬菌体侵染细菌的实验:1,噬菌体侵染细菌的实验过程:吸附→侵入→复制→组装→释放。2,DNA中P的含量多,蛋白质中P的含量少;蛋白质中有S而DNA中没有S,所以用放射性同位素35S标记一部分噬菌体的蛋白质,用放射性同位素32P标记另一部分噬菌体的DNA。用35P标记蛋白质的噬菌体侵染后,细菌体内无放射性,即表明噬菌体的蛋白质没有进入细菌内部;而用32P标记DNA的噬菌体侵染细菌后,细菌体内有放射性,即表明噬菌体的DNA进入了细菌体内。 三,烟草TMV的重建实验:1957年,Fraenkel-Conrat等人,将两个不同的TMV株系(S株系和HR株系)的蛋白质和RNA分别提取出来,然后相互对换,将S株系的蛋白质和HR株系的RNA,或反过来将HR株系的蛋白质和S株系的RNA放在一起,重建形成两种杂种病毒,去感染烟草叶片。 6.说出分子生物学的主要研究内容。 答:1,DNA重组技术;2,基因表达调控研究;3,生物大分子的结构功能研究----结构分子生物学;4,基因组、功能基因组与生物信息学研究。 第二章染色体与DNA 3.简述真核生物染色体的组成及组装过程 真核生物染色体除了性细胞外全是二倍体,DNA以及大量蛋白质及核膜构成的核小体是染色体结构的最基本单位。核小体的核心是由4种组蛋白(H2A、H2B、H3和H4)构成的扁球状8聚体。 蛋白质包括组蛋白与非组蛋白。组蛋白是染色体的结构蛋白,它与DNA组成核小体,含有大量赖氨酸核精氨酸。非组蛋白包括酶类与细胞分裂有关的蛋白等,他们也有可能是染色体的结构成分 由DNA和组蛋白组成的染色体纤维细丝是许多核小体连成的念珠状结构。 1.由DNA与组蛋白包装成核小体,在组蛋白H1的介导下核小体彼此连接形成直径约10nm的核小体串珠结构,这是染色质包装的一级结构。 2.在有组蛋白H1存在的情况下,由直径10nm的核小体串珠结构螺旋盘绕,每圈6个核小体,形成外径为30nm,内径10nm,螺距11nm的螺线管,这是染色质包装的二级结构。 3.由螺线管进一步螺旋化形成直径为0.4μm的圆筒状结构,称为超螺线管,这是染色
分子生物学复习题及其答案
一、名词解释 1、广义分子生物学:在分子水平上研究生命本质的科学,其研究对象是生物大分子的结构和功能。2 2、狭义分子生物学:即核酸(基因)的分子生物学,研究基因的结构和功能、复制、转录、翻译、表达调控、重组、修复等过程,以及其中涉及到与过程相关的蛋白质和酶的结构与功能 3、基因:遗传信息的基本单位。编码蛋白质或RNA等具有特定功能产物的遗传信息的基本单位,是染色体或基因组的一段DNA序列(对以RNA作为遗传信息载体的RNA病毒而言则是RNA序列)。 4、基因:基因是含有特定遗传信息的一段核苷酸序列,包含产生一条多肽链或功能RNA 所必需的全部核苷酸序列。 5、功能基因组学:是依附于对DNA序列的了解,应用基因组学的知识和工具去了解影响发育和整个生物体的特定序列表达谱。 6、蛋白质组学:是以蛋白质组为研究对象,研究细胞内所有蛋白质及其动态变化规律的科学。 7、生物信息学:对DNA和蛋白质序列资料中各种类型信息进行识别、存储、分析、模拟和转输 8、蛋白质组:指的是由一个基因组表达的全部蛋白质 9、功能蛋白质组学:是指研究在特定时间、特定环境和实验条件下细胞内表达的全部蛋白质。 10、单细胞蛋白:也叫微生物蛋白,它是用许多工农业废料及石油废料人工培养的微生物菌体。因而,单细胞蛋白不是一种纯蛋白质,而是由蛋白质、脂肪、碳水化合物、核酸及不是蛋白质的含氮化合物、维生素和无机化合物等混合物组成的细胞质团。 11、基因组:指生物体或细胞一套完整单倍体的遗传物质总和。 12、C值:指生物单倍体基因组的全部DNA的含量,单位以pg或Mb表示。 13、C值矛盾:C值和生物结构或组成的复杂性不一致的现象。 14、重叠基因:共有同一段DNA序列的两个或多个基因。 15、基因重叠:同一段核酸序列参与了不同基因编码的现象。 16、单拷贝序列:单拷贝顺序在单倍体基因组中只出现一次,因而复性速度很慢。单拷贝顺序中储存了巨大的遗传信息,编码各种不同功能的蛋白质。 17、低度重复序列:低度重复序列是指在基因组中含有2~10个拷贝的序列 18、中度重复序列:中度重复序列大致指在真核基因组中重复数十至数万(<105)次的重复顺序。其复性速度快于单拷贝顺序,但慢于高度重复顺序。 19、高度重复序列:基因组中有数千个到几百万个拷贝的DNA序列。这些重复序列的长度为6~200碱基对。 20、基因家族:真核生物基因组中来源相同、结构相似、功能相关的一组基因,可能由某一共同祖先基因经重复和突变产生。 21、基因簇:基因家族的各成员紧密成簇排列成大段的串联重复单位,定位于染色体的特殊区域。 22、超基因家族:由基因家族和单基因组成的大基因家族,各成员序列同源性低,但编码的产物功能相似。如免疫球蛋白家族。 23、假基因:一种类似于基因序列,其核苷酸序列同其相应的正常功能基因基本相同、但却不能合成功能蛋白的失活基因。 24、复制:是指以原来DNA(母链)为模板合成新DNA(子链)的过程。或生物体以DNA/RNA
现代分子生物学考研复习重点
现代分子生物学考研复习资料整理 第一章绪论 分子生物学:是研究核酸、蛋白质等所有生物大分子的形态、结构及其重要性、规律性和相互关系的科学 分子生物学的主要研究内容 1、DNA重组技术 2、基因表达调控研究 3、生物大分子的结构功能研究——结构分子生物学 4、基因组、功能基因组与生物信息学研究 5、DNA的复制转录和翻译 第二章染色体与DNA 半保留复制:DNA在复制过程中碱基间的氢键首先断裂,双螺旋解旋并被分开,每条链分别作为模板合成新链,产生互补的两条链。这样新形成的两个DNA分子与原来DNA分子的碱基顺序完全一样,因此,每个子代分子的一条链来自亲代DNA,另一条链则是新合成的,所以这种复制方式被称为DNA半保留复制 DNA半不连续复制:DNA双螺旋的两条链反向平行,复制时,前导链DNA的合成以5′-3′方向,随着亲本双链体的解开而连续进行复制;后随链在合成过程中,一段亲本DNA单链首先暴露出来,然后以与复制叉移动相反的方向、按照5′-3′方向合成一系列的冈崎片段,然后再把它们连接成完整的后随链,这种前导链的连续复制和后随链的不连续复制称为DNA 的半不连续复制 原核生物基因组结构特点:1、基因组很小,大多只有一条染色体2、结构简练3、存在转录单元,多顺反子4、有重叠基因 真核生物基因组的结构特点:1、真核基因组庞大,一般都远大于原核生物的基因组2、真核基因组存在大量的重复序列3、真核基因组的大部分为非编码序列,占整个基因组序列的90%以上,该特点是真核生物与细菌和病毒之间最主要区别4、真核基因组的转录产物为单顺反子5、真核基因是断裂基因,有内含子结构6、真核基因组存在大量的顺式作用元件,包括启动子、增强子,沉默子等7、真核基因组中存在大量的DNA多态性8、真核基因组具有端粒结构 DNA转座(移位)是由可移位因子介导的遗传物质重排现象 DNA转座的遗传学效应:1、转座引入插入突变2、转座产生新的基因3、转座产生的染色体畸变4、转座引起生物进化 转座子分为插入序列和复合型转座子两大类 环状DNA复制方式:θ型、滚环型和D-环型 第三章生物信息的传递(上)从DNA到RNA 转录:指拷贝出一条与DNA链序列完全相同的RNA单链的过程 启动子:是一段位于结构基因5′段上游区的DNA序列,能活化RNA聚合酶,使之与模板DNA准确地结合并具有转录起始的特异性 原核生物启动子结构:存在位于-10bp处的TATA区和-35bp处的TTGACA区,其是RNA聚合酶与启动子的结合位点,能与σ因子相互识别而具有很高的亲和力 终止子:是给予RNA聚合酶转录终止信号的DNA序列(促进转录终止的DNA序列) 终止子的类型:不依赖于ρ因子和依赖于ρ因子 增强子:能增强或促进转录起始的序列 增强子的特点:1、远距离效应2、无方向性3、顺式调节4、无物种和基因的特异性5、具
分子生物学复习题(基本完整版)
分子生物学复习题 第一章 1、蛋白质得三维结构称为构象(conformation),指得就是蛋白质分子中所有原子在三维空间中得排布,并不涉及共价键得断裂与生成所发生得变化。 2、维持与稳定蛋白质高级结构得因素有共价键(二硫键)与次级键,次级键有4种类型,即离子键、 氢键、疏水性相互作用与范德瓦力. 3、蛋白质得二级结构就是指肽链中局部肽段得构象,它们就是完整肽链构象(三级结构)得结 构单元,就是蛋白质复杂得立体结构得基础,因此二级结构也可以称为构象单元。α螺旋、β折叠就是常见得二级结构。 4、一些肽段有形成α螺旋与β折叠两种构象得可能性(或形成势),这类肽段被称为两可肽。5、两个或几个二级结构单元被连接肽段连接起来,进一步组合成有特殊几何排列得局域立体结构,称为超二级结构(介于二、三级结构间)。超二级结构得基本组织形式有αα,βαβ与ββ等3类 6、蛋白质家族(family):一类蛋白质得一级结构有30%以上同源性,或一级结构同源性很低,但它们得结构与功能相似,它们也属于同一家族。例如球蛋白得氨基酸序列相差很大,但属于同一家族.超家族(superfamily):有些蛋白质家族之间,一级结构序列得同源性较低,但在许多情况下,它们得结构与功能存在一定得相似性。这表明它们可能存在共同得进化起源。这些蛋白质家族属于同一超家族。 7、结构域就是一个连贯得三维结构,就是可互换并且半独立得功能单位,在真核细胞中由一个 外显子编码,由至少40个以上多至200个残基构成最小、最紧密也最稳定得结构,作为结构与功能单位,会重复出现在同一蛋白质或不同蛋白质中。 8、蛋白质一级结构所提供得信息有哪些?α螺旋、β折叠各自得特点? 第二章 1、DNA就是由脱氧核糖核苷酸组成得长链多聚物,就是遗传物质。具有下列基本特性:①具有稳定得结构,能进行复制,特定得结构能传递给子代;②携带生命得遗传信息,以决定生命得产生、生长与发育;③能产生遗传得变异,使进化永不枯竭。 2、DNA链得方向总就是理解为从5'-P端到3’—OH端。DNA得一级结构实际上就就是DNA 分子内碱基得排列顺序。 3、DNA就是双螺旋结构:主链由脱氧核糖与磷酸基团以3’,5’—磷酸二酯键交互连接构成得,在 双螺旋得外侧,碱基在内侧,碱基必须配对。一条链绕着另一条链旋转、盘绕,一条链上得嘌呤与另一条链上得嘧啶相互配对,嘌呤与嘧啶以氢键保持在一起. 4、双螺旋DNA熔解成单链得现象称为DNA变性。已经变性得DNA在一定条件下重新恢复 双链得过程称为复性。 5、染色质就是以双链DNA为骨架,与组蛋白(histon)、非组蛋白(non—histon)以及少量得 各种RNA等共同组成丝状结构.在染色质中,DNA与组蛋白得组成非常稳定,非组蛋白与RNA随细胞生理状态不同而有变化。 6、常染色质就是在细胞间期核内染色体折叠压缩程度较低,处于伸展状态,碱性染性着色较浅
医学分子生物学复习总结学习资料.doc
医学分子生物学复习资料
蛋白质、糖蛋白与蛋白聚糖、脂蛋白、细胞信号传导 名词解释: 1、构型:指一个有机分子中各个原子特有的固定的空间排列。这种排列不经过 共价键的断裂和重新形成是不会改变的。不同构型之间相互转化会涉及化学键 的断裂,构型的改变往往使分子的光学活性发生变化。 2、构象:构成分子的原子和基团因为化学键的旋转而形成在三维空间的不同的 排布、走向。不同的构象之间可以相互转化而不涉及化学键的破裂。构象改变 不会改变分子的光学活性。 3、肽平面:肽键具有部分双键性质而不能自由旋转,这样C、N 原子同它们连接的 O、H和两个 Cα共六个原子就被约束在一个刚性平面上,这个平面被称为肽平面。 4、基序或模体:相邻的几个二级结构相互作用形成有规则的组合体称为超二级 结构,是特殊的序列或结构的基本组成单元,又称为基序或模体。 5、结构域:蛋白质的超二级结构进一步组合折叠成半独立紧密的球状结构域。 6、糖蛋白:在分子组成中以蛋白质为主,其一定部位以共价键与若干糖链(约4%)相连所构成的分子。 7、蛋白聚糖:蛋白聚糖是一类由蛋白质和糖胺聚糖通过共价键相连而成的化合物,其分子中的含糖量通常为50%~90%。 8、血脂:血浆所含的脂类统称为血脂,它包括甘油三酯、磷脂、胆固醇及游离 脂酸。 9、血浆脂蛋白:在血浆中血脂与蛋白质结合,形成血浆脂蛋白。 10、载脂蛋白:血浆脂蛋白中蛋白质部分称为载脂蛋白。 11、脂蛋白受体:脂蛋白受体是一类位于细胞膜上的糖蛋白,它们能以高亲和 性的方式与其相应的脂蛋白配体相互作用,介导细胞对脂蛋白的摄取和代谢, 从而进一步调节血浆脂蛋白和血脂的水平。 12、细胞通讯( cell communication):指一个细胞发出的信息通过介质传递 到另一个细胞产生相应反应的过程。
现代分子生物学朱玉贤课后习题答案
现代分子生物学(第3版)朱玉坚第二章染色体与DNA课后思考 题答案 1 染色体具有哪些作为遗传物质的特征? 1 分子结构相对稳定 2 能够自我复制,使亲子代之间保持连续性 3 能够指导蛋白质的合成,从而控制整个生命过程 4 能够产生可遗传的变异 2.什么是核小体?简述其形成过程。 由DNA和组蛋白组成的染色质纤维细丝是许多核小体连成的念珠状结构。核小体是由H2A,H2B,H3,H4各两个分子生成的八聚体和由大约200bp的DNA组成的。八聚体在中间,DNA分子盘绕在外,而H1则在核小体外面。每个核小体只有一个H1。所以,核小体中组蛋白和DNA的比例是每200bpDNA有H2A,H2B,H3,H4各两个,H1一个。用核酸酶水解核小体后产生只含146bp核心颗粒,包括组蛋白八聚体及与其结合的146bpDNA,该序列绕在核心外面形成1.75圈,每圈约80bp。由许多核小体构成了连续的染色质DNA细丝。 核小体的形成是染色体中DNA压缩的第一阶段。在核小体中DNA盘绕组蛋白八聚体核心,从而使分子收缩至原尺寸的1/7。200bpDNA完全舒展时长约68nm,却被压缩在10nm的核小体中。核小体只是DNA压缩的第一步。 核小体长链200bp→核酸酶初步处理→核小体单体200bp→核酸酶继续处理→核心颗粒146bp 3简述真核生物染色体的组成及组装过程 除了性细胞外全是二倍体是有DNA以及大量蛋白质及核膜构成核小体是染色体结构的最基本单位。核小体的核心是由4种组蛋白(H2A、H2B、H3和H4)各两个分子构成的扁球状8聚体。 蛋白质包括组蛋白与非组蛋白。组蛋白是染色体的结构蛋白,它与DNA组成核小体,含有大量赖氨酸核精氨酸。非组蛋白包括酶类与细胞分裂有关的蛋白等,他们也有可能是染色体的结构成分 由DNA和组蛋白组成的染色体纤维细丝是许多核小体连成的念珠状结构---- 1.由DNA与组蛋白包装成核小体,在组蛋白H1的介导下核小体彼此连接形成直径约10nm的核小体串珠结构,这是染色质包装的一级结构。 2.在有组蛋白H1存在的情况下,由直径10nm的核小体串珠结构螺旋盘绕,每圈6个核小体,形成外径为30nm,内径10nm,螺距11nm的螺线管,这是染色质包装的二级结构。 3.由螺线管进一步螺旋化形成直径为0.4μm的圆筒状结构,称为超螺线管,这是染色质包装的三级结构。 4.这种超螺线管进一步螺旋折叠,形成长2-10μm的染色单体,即染色质包装的四级结构。 4. 简述DNA的一,二,三级结构的特征 DNA一级结构:4种核苷酸的的连接及排列顺序,表示了该DNA分子的化学结构 DNA二级结构:指两条多核苷酸链反向平行盘绕所生成的双螺旋结构 DNA三级结构:指DNA双螺旋进一步扭曲盘绕所形成的特定空间结构 5.原核生物DNA具有哪些不同于真核生物DNA的特征? 1, 结构简练原核DNA分子的绝大部分是用来编码蛋白质,只有非常小的一部分不转录,这与真核DNA的冗余现象不同。 2, 存在转录单元原核生物DNA序列中功能相关的RNA和蛋白质基因,往往丛集在基因组的一个或几个特定部位,形成功能单元或转录单元,它们可被一起转录为含多个mRNA的分子,称为多顺反子mRNA。 3, 有重叠基因重叠基因,即同一段DNA能携带两种不同蛋白质信息。主要有以下几种情况①一个基因完全在另一个基因里面②部分重叠③两个基因只有一个碱基对是重叠的 6简述DNA双螺旋结构及其在现代分子生物学发展中的意义 DNA的双螺旋结构分为右手螺旋A-DNA B-DNA 左手螺旋Z-DNA DNA的二级结构是指两条都核苷酸链反向平行
现代分子生物学名词解释
现代分子生物学名词解释 1. 现代分子生物学必考要点超全版必考要点 2. 基因 产生一条多肽链或功能RNA所必需的全部核苷酸序列。 3. 基因组 基因组是生物体内遗传信息的集合,是指某个特定物种细胞内全部DNA分子的总和。 4. 顺反子 由顺/反测验定义的遗传单位,与基因等同,都是代表一个蛋白质质的DNA 单位组成。一个顺反子所包括的一段DNA与一个多肽链的合成相对应。 5. 基因表达 DNA分子在时序和环境的调节下有序地将其所承载的遗传信息通过转录和翻译系统转变成蛋白质分子(或者RNA分子),执行各种生理生化功能,完成生命的全过程。 6. ribozyme【已考试题】 即核酶,由活细胞所分泌的具有像酶那样催化功能的RNA分子。 7. SD序列 原核生物起始密码AUG上游7~12个核苷酸处的一段保守序列,能与16S rRNA 3′端反向互补,被认为在核糖体-mRNA的结合过程中起作用。 8. 限制性内切酶 限制性内切酶是一类能够识别双链DNA分子中的某种特定核苷酸序列,并在相关位置切割DNA双链结构的核酸内切酶。 9. 内含子和外显子 真核细胞DNA分子中能转录到mRNA前体分子中但会在翻译前被切除的非编码区序列称内含子。而编码区称为外显子。 10. C值和C值反常现象 C值指一种生物单倍体基因组DNA的总量,一般随生物进化而增加,但也存在某些低等生物的C值比高等生物大,即C值反常现象。原因是真核生物基因组中含大量非编码序列。 11. 卫星DNA 在DNA链上串联重复多次的短片段碱基序列。因能在密度梯度离心中区别与主DNA峰而单独成小峰而得名。 12. 重叠基因 一段能够携带多种不同蛋白质信息的DNA片段。 13. 断裂基因【已考试题】 在DNA分子的结构基因内既含有能转录翻译的片段,也含有不转录翻译的片段,这类基因称断裂基因。 14. 复制子【已考试题】 DNA分子上一个独立的复制单位,包括复制原点。 15. 同义突变
分子生物学复习资料
分子生物学复习资料 2.分子杂交:是核酸研究中一项最差不多的实验技术。其差不多原理确实是应用核酸分子的变性和复性的性质,使来源不同的DNA或RNA片段,按碱基互补关系形成杂交双链分子。 3.限制性核酸内切酶:是能够识别DNA的特异序列,并在识别位点或其周围切割双链DNA的一类内切酶,简称限制酶。 4.cDNA:与RNA链互补的单链DNA,以其RNA为模板,在适当引物的存在下,由RNA与DNA进行一定条件下合成的,确实是cDNA。 5.基因组DNA:组成生物基因组的所有DNA。 6.基因(gene):是生物体遗传物质的差不多单位,是载有特定遗传信息的DNA分子的片段。 7.基因组(genome):一个细胞或病毒所携带的全部遗传信息或整套基因。 8.基因表达(gene expression):是指储存遗传信息的基因通过一系列步骤表现出其生物功能的整个过程,即转录和翻译过程。 9.时刻特异性(temporal specificity):单细胞生物的某一特定基因的表达严格按特定的时刻顺序发生;多细胞生物的相应基因的表达在机体发育的不同时期严格按一定的时刻顺序开启或关闭。 10.空间特异性(spatial specificity):在机体发育的某一时期,同一个基因的表达产物在不同的组织器官分布不同。 11.组成性基因表达(总管基因):不大受环境变动而变化的一类基因表达,在个体生长过程中,几乎在所有的组织中连续表达或变化专门小。这些基因一样被称为总管基因。 12.反式调剂(trans-regulation):由某一基因表达产生的蛋白质因子,通过与另一基因的特异的顺式作用元件相互作用,调剂其表达。 13.顺式调剂(cis-regulation):由蛋白质因子可特异识别、结合自身基因的调剂序列,调剂自身基因的表达。 14.单顺反子:在多数真核生物中,编码蛋白质的基因的初级转录物,被加工成一种mRNA,一样翻译出一条多肽链。
现代分子生物学要点总结(朱玉贤版)
现代分子生物学要点总结(朱玉贤版) 一、绪论 两个经典实验 1、肺炎球菌在老鼠体内的毒性实验:先将光滑型致病菌(S型)烧煮杀活性以后、以及活 的粗糙型细菌(R型)分别侵染小鼠发现这些细菌自然丧失了治病能力;当他们将经烧煮杀死的S型细菌和活的R型细菌混合再感染小鼠时,实验小鼠每次都死亡。解剖死鼠,发现有大量活的S型细菌。实验表明,死细菌DNA进行了可遗传的转化,从而导致小鼠死亡。 2、T2噬菌体感染大肠杆菌:当细菌培养基中分别带有35S或32P标记的氨基酸或核苷酸, 子代噬菌体就相应含有35S标记的蛋白质或32P标记的核酸。分别用这些噬菌体感染没有放射性标记的细菌,经过1~2个噬菌体DNA复制周期后进行检测,子代噬菌体中几乎不含带35S标记的蛋白质,但含30%以上的32P标记。说明在噬菌体传代过程中发挥作用的可能是DNA而不是蛋白质。 基因的概念:基因是产生一条多肽链或功能RNA分子所必需的全部核苷酸序列。 二、染色体与DNA 嘌呤嘧啶 腺嘌呤鸟嘌呤胞嘧啶尿嘧啶胸腺嘧啶 染色体 性质:1、分子结构相对稳定;2、能够自我复制,使亲、子代之间保持连续性;3、能指导蛋白质的合成,从而控制生命过程;4、能产生可遗传的变异。 组蛋白一般特性:1、进化上极端保守,特别是H3、H4;2、无组织特异性;3、肽链上氨基酸分布的不对称性;4、存在较普遍的修饰作用;5、富含赖氨酸的组蛋白H5 非组蛋白:HMG蛋白;DNA结合蛋白;A24非组蛋白 真核生物基因组DNA 真核细胞基因组最大特点是它含有大量的重复序列,而且功能DNA序列大多被不编码蛋白
质的非功能蛋白质所隔开。人们把一种生物单倍体基因组DNA的总量称为C值,在真核生物中C值一般是随着生物进化而增加的,高等生物的C值一般大于低等动物,但某些两栖类的C值甚至比哺乳动物还大,这就是著名的C值反常现象。真核细胞DNA序列可被分为3类:不重复序列、中度重复序列、高度重复序列。 真核生物基因组的特点:1、真核生物基因组庞大,一般都远大于原核生物的基因组;2、真核基因组存在大量的的重复序列;3、真核基因组的大部分为非编码序列,占整个基因组序列的90%以上,这是真核生物与细菌和病毒之间的最主要的区别;4、真核基因组的转录产物为单顺反之;5、真核基因组是断裂基因,有内含子结构;6、真核基因组存在大量的顺式元件,包括启动子、增强子、沉默子等;7、真核基因组中存在大量的DNA多态性;8、真核基因组具有端粒结构。 原核生物基因组的特点:1、结构简练,绝大部分用来编码蛋白质,只有很少一部分控制基因表达的序列不转录;2、存在转录单元,原核生物DNA序列中功能相关的RNA和蛋白质基因,往往丛集在基因组的一个或者几个特定部位,形成功能单位或转录单元,可以被一起转录为含多个mRNA的分子;3、有重叠基因,所谓重叠基因就是同一段DNA携带两种或以上不同的蛋白质的编码信息。 DNA的结构 DNA又称脱氧核糖核酸,是deoxyribonucleic acid的简称。 L=T+W,L指环形DNA分子两条链间交叉的次数,只要不发生断裂,L是一个常量。T为双螺旋的盘绕数,W为超螺旋数。双螺旋DNA的松开导致负超螺旋,而拧紧则导致正超螺旋。 双螺旋碱基间距(nm)螺旋直径(nm)每轮碱基数螺旋方向 A-DNA0.26 2.611右 B-DNA0.34 2.010右 Z-DNA0.37 1.812左 DNA的复制 半保留复制:Semi-conservative replication;半不连续复制:Semi-discontinuous replication 把生物体的复制单位称为复制子,一个复制子只含一个复制起始点。 归纳起来,无论是原核生物还是真核生物,复制起点是固定的,表现为固定的序列,并识别参与复制起始的特殊蛋白质。复制叉移动的方向和速度虽是多种多样的,但以双向等速方式为主。 复制的几种主要方式 双链DNA的复制大都以半包六复制方式进行的,通过“眼”型、θ型、滚环型或D-环型等以复制叉的形式进行。 1、线性DNA双链进行双向复制时,由于已知的DNA聚合酶和RNA聚合酶都只能从5’ 到3’移动,所以,复制叉呈眼型; 2、环状双链DNA复制可分为θ型、滚环型和D-环形几种类型 Ⅰ、θ型,大肠杆菌染色体DNA是环状双链DNA,它的复制是典型的θ型复制,从一个起点开始,同时向两个方向进行复制,当两个复制叉相遇时,复制就停止 Ⅱ、滚环型,是单向复制的一种特殊方式,在噬菌体中很常见。DNA的合成由对正链原点的专一切割开始,所形成的自由5’端被从双链环中置换出来并为单链DNA结合蛋白所覆盖,
分子生物学终极复习资料汇总
《分子生物学》复习题 1、染色体:是指在细胞分裂期出现的一种能被碱性染料强烈染色,并具有一定 形态、结构特征的物体。携带很多基因的分离单位。只有在细胞分裂中才可见的形态单位。 2、染色质:是指细胞周期间期细胞核内由DNA、组蛋白、非组蛋白和少量RNA 组成的复合结构,因其易被碱性染料染色而得名。 3、核小体:染色质的基本结构亚基,由约200 bp的DNA和组蛋白八聚体所组 成 4、C值谬误:一个有机体的C值与它的编码能力缺乏相关性称为C值矛盾 5、半保留复制:由亲代DNA生成子代DNA时,每个新形成的子代DNA中, 一条链来自6、亲代DNA,而另一条链则是新合成的,这种复制方式称半保留复制 6、DNA重组技术又称基因工程,目的是将不同的DNA片段(如某个基因或基 因的一部分)按照人们的设计定向连接起来,在特定的受体细胞中与载体同时复制并得到表达,产生影响受体细胞的新的遗传性状。 7、半不连续复制:DNA复制时其中一条子链的合成是连续的,而另一条子链的 合成是不连续的,故称半不连续复制。 8、引发酶:此酶以DNA为模板合成一段RNA,这段RNA作为合成DNA的引 物(Primer)。实质是以DNA为模板的RNA聚合酶。 9、转坐子:存在与染色体DNA上可自主复制和位移的基本单位。 10、多顺反子:一种能作为两种或多种多肽链翻译模板的信使RNA,由DNA 链上的邻近顺反子所界定。 11、基因:产生一条多肽链或功能RNA所必需的全部核甘酸序列。 12、启动子:指能被RNA聚合酶识别、结合并启动基因转录的一段DNA序列。 13、增强子:能强化转录起始的序列 14、全酶:含有表达其基础酶活力所必需的5个亚基的酶蛋白复合物,拥有σ因子。 (即核心酶+σ因子) 15、核心酶:仅含有表达其基础酶活力所必需亚基的酶蛋白复合物,没有σ因子。 16、核酶:是一类具有催化功能的RNA分子 17、三元复合物:开放复合物与最初的两个NTP相结合,并在这两个核苷酸之间形成磷酸二酯键后,转变成包括RNA聚合酶,DNA和新生的RNA的三元复合物。 18、SD序列:mRNA中用于结合原核生物核糖体的序列。30S亚基通过其
分子生物学复习资料
第十一章细胞异常增生性的分子机制 (一)选择题A型题 1. 下列哪种细胞通过干细胞分裂补充(D. 血细胞) 2. 下列关于细胞凋亡的说法不.正确的是A. A. 隐性时相出现核浓缩 B. 执行时相出现凋亡小体 C. 隐性时相可逆 D. 执行时相不可逆 E. 不伴有炎症反应 3. 在线粒体上形成通道的分子(B. Bax) 4. 细胞癌基因活化的机制A. 点突变B. DNA重排 C. 基因扩增或病毒基因启动子及增强子插入 D. 染色体易位 5. 滤泡型B淋巴瘤的发生机制是(D. 染色体易位) 6. 关于细胞癌基因说法不.正确的是(A.) A.正常细胞本身没有 B. 含有内含子 C. 编码产物调控细胞增殖 D. 可能为病毒癌基因的源头 E. 通过多种机制活化 7. 属于癌基因的是(D. ras基因)8. 属于抑癌基因的是(E.APC基因) 9. 关于Rb基因下列说法正确的是(E. 编码产物使细胞停留于G1期) 10. 关于p53基因说法正确的是(E. 编码产物可促使细胞凋亡) 11. 与滤泡型B淋巴瘤发生相关的基因是(A. bcl 2)12. 编码产物为小G蛋白的基因是(B. k-ras) 13.c-sis编码产物是(A. 生长因子)14.下列细胞因子在细胞外基质沉积过程中作用最显著的是(C. TGFβ)15. 关 于p53的叙述,哪项是正确的(B. p53基因是肿瘤抑制基因) 16.病毒癌基因v-erb B的产物属于B.生长因子受体17.ras基因家族编码产物属于C.胞内信号转导蛋白18. c-src基因编码产物属于C. 胞内信号转导蛋白19. c-myc基因编码产物属于D. 转录因子 20. p53基因编码产物属于D. 转录因子21c-ras基因编码蛋白质中氨基酸发生替换属于A. 点突变 22. 慢性髓细胞性白血病中c-ABL与BCR基因对接属于D. 染色体易位 23. 病毒的长末端重复(LTR)序列调控c-myc表达属于E. 启动子及增强子的插入 24. 免疫球蛋白重链基因的转录活化部位与bcl 2基因连接属于D. 染色体易位 25. 部分小细胞肺癌中c-myc编码产物过度增加属于C. 基因扩增 26.参与血管生成的细胞因子有A. PDGF B. VEGF C. TGFβD. 纤溶酶原激活物系统E. TNF 27. 关于p53下列说法正确的是 A. p53蛋白是位于细胞核内的一种转录因子 B. p53基因位于17p13.1 C. 突变的p53蛋白可与野生型p53蛋白聚合成无功能的四聚体 D. 可促进靶基因p21和GADD45转录 28. 凋亡体(apoptosome)包括下列哪几种分子A. caspase 9 B. Apaf 1 C. 细胞色素c 29. 多发性腺瘤样息肉转变为结肠癌A. APC基因失活B.DNA甲基化程度降低C.原癌基因k-ras活化D.抑癌基因DCC失活E.抑癌基因p53失活 30. 关于caspase下列说法正确的是 A. 以酶原的形式合成 B. 活性中心含有半胱氨酸残基 C. 作用于底物特异部位的天门冬氨酸残基羧基侧的肽键 D. 成熟的酶包括p20结构域和p10结构域 E. 常以级联活化的方式发挥功能 31. 与生理性血管生成相比,肿瘤血管生成的特点有A. 多了征用式的生成方式B. 调节因子可来源于肿瘤细胞 C. 血管内皮细胞常常不完整 D. 血管常无平滑肌细胞等保护 32. 关于Rb基因,下列说法正确的是A. 第一个被发现的抑癌基因B. 编码产物定位于细胞核、C. 低磷酸化的pRb结合并灭活E2F1 D. 全长200 kb E. 含27个外显子 1.病毒癌基因反转录病毒中一些能在体外转化细胞,在体内使宿主患肿瘤的基 2.细胞癌基因(正常宿主细胞 中与病毒癌基因同源的基因,往往编码生长因子或其他信号转导分子,为细胞生长所必需;只有特定条件下被活化后才有致癌性,又称为原癌基因。