脂肪代谢信号通路
糖脂代谢病的发病机制多重打击学说
龙源期刊网 https://www.360docs.net/doc/af7873118.html, 糖脂代谢病的发病机制:多重打击学说 作者:华爽吕明慧刘倩颖何兴祥荣向路叶得伟郭姣 来源:《世界中医药》2019年第03期 摘要;血糖異常、血脂异常、非酒精性脂肪肝、超重、高血压、动脉粥样硬化性心脑血管病等代谢性疾病发病率居高不下,是世界性难题。临床流行病学研究目前已证实,2型糖尿病、高脂血症等代谢性疾病常合并发生,但目前对导致上述代谢异常发生的分子机制尚未阐明,并制约了综合防控疗效优良的创新药物和诊疗手段的研发。郭姣教授率团队基于大样本临床流行病学、转化研究数据,提出“糖脂代谢病”创新理论,认为上述代谢异常以糖、脂代谢紊乱为特征,发病过程由遗传、环境、精神等多种因素参与,以神经-内分泌失调、胰岛素抵抗、氧化应激、炎性反应、肠道菌群失调为核心病理,以高血糖、血脂失调、非酒精性脂肪肝、超重、高血压及动脉粥样硬化等单一或合并出现为主要临床表现特点。本文综合神经-内分泌-免疫紊乱、胰岛素抵抗、氧化应激、炎性反应、肠道菌群失调等环节与糖脂代谢异常及其诱发多器官病变的病理机制的研究进展,提出糖脂代谢病发病机制的“多重打击学说”。该学说对于揭示多种代谢异常发生的核心、共性分子机制及从病证结合角度阐释中医证候的生物学本质具有重要意义。 关键词;糖脂代谢病;发病机制;神经-内分泌轴;胰岛素抵抗;氧化应激;代谢性炎性反应;肠道 菌群失调 The Multiple-hit Pathogenesis of Glucolipid Metabolic Disorders Hua Shuang1,2,3,Lyu Minghui1,2,3,Liu Qianying1,2,3,He Xingxiang2,Rong Xianglu1,2,3,Ye Dewei1,2,3,Guo jiao1,2,3 (1 Joint Laboratory between Guangdong and Hong Kong on Metabolic Diseases,Guangdong Pharmaceutical University,280 Waihuan Road East,Guangzhou Higher Education Mega,Guangzhou 510006,China; 2 Guangdong Metabolic Disease Research Center of Integrated Chinese and Western Medicine,Guangdong Pharmaceutical University,280 Waihuan Road East,Guangzhou Higher Education Mega,Guangzhou 510006,China; 3 Institute of Traditional Chinese medicine,Guangdong Pharmaceutical University,280 Waihuan Road East,Guangzhou Higher Education Mega,Guangzhou 510006,China) Abstract;The high prevalence and incidence of hyperglycemia,dyslipidemia,nonalcoholic fatty liver disease,obesity,hypertension,atherosclerosis and its related cardiovascular diseases has emerged as one of leading causes of morbidity and mortality worldwide.Epidemiological data well established that two or several above-mentioned metabolic disorders usually co-exist in obese subjects.However,the mechanisms underlying the co-existence of these metabolic disorders have not been well characterized currently,exerting negative effect on the development of new drugs and therapeutic approaches for these diseases.Based on the data from epidemiological and translational
脂类代谢考试试题及答案
第九章脂类代谢 一、选择题(请将选择的正确答案的字母填写在题号前面的括号内) ()1合成甘油酯最强的器官是 A 肝; B 肾; C 脑; D 小肠。 ()2、小肠粘膜细胞再合成脂肪的原料主要来源于 A 小肠粘膜吸收来的脂肪水解产物; B 肝细胞合成的脂肪到达小肠后被消化的产物 C 小肠粘膜细胞吸收来的胆固醇水解产物; D 脂肪组织的水解产物; E 以上都对。 ()3、线粒体外脂肪酸合成的限速酶是 A 酰基转移酶; B 乙酰辅酶A羧化酶; C 肉毒碱脂酰辅酶A转移酶Ⅰ; D 肉毒碱脂酰辅酶A转移酶Ⅱ; E β—酮脂酰还原酶。 ()4、酮体肝外氧化,原因是肝脏内缺乏 A 乙酰乙酰辅酶A硫解酶; B 琥珀酰辅酶A转移酶; C β—羟丁酸脱氢酶; D β—羟—β—甲戊二酸单酰辅酶A合成酶; E 羟甲基戊二酸单酰辅酶A裂解酶。 ()5、卵磷脂含有的成分是 A 脂肪酸、甘油、磷酸和乙醇胺; B 脂肪酸、甘油、磷酸和胆碱; C 脂肪酸、甘油、磷酸和丝氨酸; D 脂肪酸、磷酸和胆碱; E 脂肪酸、甘油、磷酸。 ()6、脂酰辅酶A的β—氧化过程顺序是 A 脱氢、加水、再脱氢、加水; B 脱氢、脱水、再脱氢、硫解; C 脱氢、加水、再脱氢、硫解; D 水合、加水、再脱氢、硫解。 ()7、人体内的多不饱和脂肪酸是指 A 油酸、软脂肪酸; B 油酸、亚油酸; C 亚油酸、亚麻酸; D 软脂肪酸、亚油酸。 ()8、可由呼吸道呼出的酮体是 A 乙酰乙酸; B β—羟丁酸; C 乙酰乙酰辅酶A; D 丙酮。 ()9、与脂肪酸的合成原料和部位无关的是
A 乙酰辅酶A; B NADPH+H+; C 线粒体外; D 肉毒碱;E、HCO3- ()10、并非以FAD为辅助因子的脱氢酶有 A 琥珀酸脱氢酶; B 脂酰辅酶A脱氢酶; C 二氢硫辛酸脱氢酶; D β—羟脂酰辅酶A脱氢酶。 ()11、不能产生乙酰辅酶A的是 A 酮体; B 脂肪酸; C 胆固醇; D 磷脂; E 葡萄糖。 ()12、甘油磷酸合成过程中需哪一种核苷酸参与 A ATP; B CTP; C TTP; D UDP; E GTP。 ()13、脂肪酸分解产生的乙酰辅酶A的去路 A 合成脂肪酸; B 氧化供能; C 合成酮体; D 合成胆固醇; E 以上都是。()14、胆固醇合成的限速酶是 A HMGCoA合成酶; B 乙酰辅酶A羧化酶; C HMGCoA还原酶; D 乙酰乙酰辅酶A硫解酶。 ()15、胆汁酸来源于 A 胆色素; B 胆红素; C 胆绿素; D 胆固醇。 ()16、脂肪酸β—氧化的限速酶是 A 肉毒碱脂酰转移酶Ⅰ; B 肉毒碱脂酰转移酶Ⅱ C 脂酰辅酶A脱氢酶; D β—羟脂酰辅酶A脱氢酶; E β—酮脂酰辅酶A硫解酶。 ()17、β—氧化过程的逆反应可见于 A 胞液中脂肪酸的合成; B 胞液中胆固醇的合成; C 线粒体中脂肪酸的延长; D 内质网中脂肪酸的合成。 ()18、并非类脂的是 A 胆固醇; B 鞘脂; C 甘油磷脂; D 神经节苷脂; E 甘油二脂。 ()19、缺乏维生素B2时,β—氧化过程中哪一个中间产物合成受到障碍? A 脂酰辅酶A; B β—酮脂酰辅酶A; C α,β—烯脂酰辅酶A ; D L—β—羟脂酰辅酶A; E 都不受影响。 ()20、合成胆固醇的原料不需要 A 乙酰辅酶A; B NADPH; C A TP ; D O2。 ()21、由胆固醇转变而来的是
代谢组研究利器之脂质组-定量脂质组
一、研究热点--脂质组 近年来,脂质组学研究非常热门,经常出现在CNS期刊上。 脂质是重要的生物大分子物质之一,在生物体的生命活动中起着重要作用。2003年韩贤林教授等首次提出脂质组学的概念,对生物体系脂质进行全面系统的研究分析。它主要研究生物体系(生物体、组织、细胞甚至亚细胞)受刺激或扰动后,脂质种类、亚种类或单个脂质分子的变化。通过系统的研究机体内脂类物质代谢的变化,从而揭示与其他分子间相互作用的机理。 脂质组学是代谢组学的一个分支。脂类代谢(如血浆中约70%的代谢物是脂类)是动植物的代谢中第一大类物质,是动植物代谢研究中最为关注的热点,参与能量运输、细胞间的信息通讯与网络调控等生长发育过程。 脂类作为脂质组学研究的内容,依据“脂质代谢途径研究计划”(LIPIDMAPS)可分为8个大的类别:1.脂肪酰,2.甘油脂,3.甘油磷脂,4.鞘脂,5.甾醇酯,6.丙烯醇脂,7.糖脂,8.聚酮。脂类物质不仅是我们人体的重要组成成分,而且不少疾病也与脂类异常代谢有关,如阿兹海默症、糖尿病、肥胖以及肿瘤发生发展等。 我们都知道细胞膜的主要成分是磷脂双分子层,大多数脂类参与构建了细胞膜和亚细胞膜。脂类既是结构分子,也是信号分子。一个典型的例子是磷脂酰肌醇-4,5-二磷酸转化成二酰基甘油和三磷酸肌醇,后者作为第二信使,激活下游的激酶并诱导细胞内钙离子的释放。 脂质组学领域中最核心的研究手段是电喷雾电离-质谱技术,能对各种脂质尤其是磷脂进行高分辨率、高灵敏度、高通量的分析。 二、定量脂质组 脂类具有数目众多、结构多样的特点,这就给定量脂质组分析带来了一定的难度。定量脂质组学是通过一种或多种稳定同位素标记内标对脂质进行大规模绝对定量的一种方法。因此定量脂质组分析具有以下要点: 1.LC-MS/MS仪器进行脂质定量,最优的检测模式是SRM/MRM; 2.不同类别的脂质在仪器中响应强度不一样,变化趋势不一样,因此内标
22脂肪酸的分解代谢
第28章、脂肪酸的分解代谢(p230) 本章重点:1、脂肪酸分解代谢过程,2、脂肪酸代谢的能量产生,3、脂肪酸分解脱氢,4 脂肪酸分解代谢和糖酵解的关系。 本章主要内容: 一、脂肪的水解——脂酶的水解作用(细胞质中) 生物体内脂肪是由脂肪酶水解,在脂肪酶的催化下生成一分子甘油和三分子脂肪酸,脂 肪酶的特点:主要作用于有酯键的化合物,不论脂肪来源于什么组织,不论脂肪酸碳链的长 短,只要是酯键,脂肪酶就可以使其断裂,这就是酶的专一性即键专一性。 事实上,脂肪的水解不是一步完成的,而是分步完成,分步进行水解。第一步脂肪酶水 解第一或第三全酯键,即a或a'酯键,如果第一步水解a -酯键,第二水解a '酯键,生成a和a'脂肪酸和甘油-酯,最后,3 -位的脂肪酸在转移酶的催化下3 -的脂肪酸转到a 或a'位上,再在脂肪酶的作用下,脂肪酸水解下来,共生成三分子脂肪酸和一分子甘油,水解过程为:脂肪(甘油三酯)水解的产物:一分子甘油和三分子脂肪酸。 二、甘油的转化 脂肪的水解产物甘油是联系脂肪代谢和糖代谢的重要化合物,它可以轩化成磷酸甘油醛 进入糖代谢,其代谢过程为: 生成的磷酸2羟丙酮有两种去路: 1、DHAF可以进入EMP途径生成pyr,再经脱氢、脱羟生成乙酰COA经TCA循环氧化成CQ和H2O 2、G-3-P可以与DHAP逆EMP途径在醛缩酶催化下生成F-1.6-P,继续转化成糖类。 甘油被彻底氧化以后可以生成多少molATP呢?首先总结氧化的部位: ①a-磷酸甘油脱氢,生成ImolNADH H ②G-3-P 生成1, 3-DPG 1molNADH H ③Pyr 脱氢1molNADH H ④异柠檬酸脱氢1molNADHH+ ⑤ a -酮戊二酸脱氢1molNADH H+ ⑥平果酸脱氢1molNADH H+ ⑦琥珀酸脱氢1molFADH 2 琥珀酰COA>琥珀酸 另外,甘油还可在代谢的过程中转化到蛋白质中去,如进入TCA后生成Pyr、OAA a -Kg等可经转氨基作用生成Ala、Asp和Glu参与到蛋白质的合成中去。 三、脂肪酸的降解 脂肪酸的降解(分解)即氧化分解有几种形式,最重要的是 3 -氧化,其次是a -氧化和 3 -氧化。 (一)3 -氧化(线粒体内进行) 1、概念:脂肪酸的3 -氧化作用是脂肪酸经一系列酶的作用,从a、3碳位之间断裂生 成1mol乙酰COA和比原来脂肪酸少两个碳原子的脂酰COA 2、3 -氧化过程:脂肪酸3 -氧化的合成过程包括下列几个主要步骤: 1)活化或叫做脂酰COA的形成:脂肪酸首先与辅酶A缩合同时消耗一分子ATP形成活化的脂酰COA这步反应要消耗ATP的两个高能磷酸键。 第一步反应是在脂酰COA合成酶的催化下进行的,活化了的脂酰COA借线粒体内膜两侧的肉毒碱脂酰COA专移酶的作用,进入线粒体内。 肉毒碱脂酰COA专移酶 脂酰COA肉毒碱脂酰肉毒碱+COA 肉毒碱的结构: 肉毒碱起携带脂肪酸酰基通过线粒体内膜的作用。 肉毒碱脂酰COA转移酶有两个同工酶,一是位于内膜外侧的肉毒碱脂酰COA转移
脂肪进行合成代谢的过程
郑州增肥专科医院 来源:河南省现代研究院中医院增肥专科脂肪是怎样消耗的——脂肪分解的“三大环节” 为了方便大家理解这个相对专业的生化反应过程,我画了一张图(如下),我就按图解说了。 建议大家先仔细阅读一下图,再接着看下文—— 第一环节:脂肪动员 我们的脂肪主要以“甘油三酯(TG)”的形式储存在脂肪组织内,另外,心肌、骨骼肌、血浆中也有少量甘油三酯存在。对于减肥瘦身来说,主要是将脂肪组织内的甘油三酯动员起来用于供能,才能达到理想的效果。如果一个人脂肪动员的能力较低,就更容易产生肥胖,或者更不容易减肥。 一些特定的食物也能促进脂肪动员,如茶(茶多酚、咖啡碱)、咖啡、辣椒,以及瓜拉那等草本提取物,同时伴有心跳加速、血压增高的反应,因此需慎重使用。 第二环节:活性脂酸转移 当脂肪酸从脂肪组织中分解出来进入血浆后,在血浆蛋白的帮助下运送到全身各处的活动细胞内,开始了它的第二个环节——活化。只有被活化的脂肪酸才能进入被称作“细胞内动力工厂”的“线粒体”内,进一步被氧化分解。这个进入过程就是第三环节:活性脂酸转移。 脂肪酸被活化是受一系列酶的催化作用完成的,因此,这些酶的活性成为脂肪分解的一个限制因素。当然,这个因素主要受遗传决定,同时也受特定的代谢物质(如共轭亚油酸,CLA)影响。 第三环节:脂肪酸β氧化 这是脂肪酸在线粒体内最后被分解成二氧化碳和水,并产生能量的过程,受一系列酶和其他代谢反应影响。值得重视的是,脂肪酸的β氧化和糖的氧化在最后阶段都必须进入一个叫“三羧酸循环”的生化反应过程,才能最终分解成二氧化碳和水,最大限度地释放能量。
如果脂肪分解过程中,糖供应不足,导致三羧酸循环不能顺利进行,脂肪分解也会受到抑制,从而产生“酮体”。高浓度的酮体对人体是有害的,可能造成“酮中毒”。
糖脂代谢异常指导方案
一、保肝肝指导 (一)生活起居: 1、注意肝脏保护,禁烟限酒,合理用药,减少酒精性、药物性肝损伤。 2、保证良好睡眠,避免熬夜,夜间12-2点是肝脏排毒的最佳时间,熬夜会降低肝脏排毒效果,加重肝脏负担。 3、生活中注意避免各种化学物质对肝脏的慢性伤害如:烟尘、汽车尾气、家装材料及烟酒刺激等。 (二)饮食指导 1、低脂、适量高蛋白及高维生素饮食,高蛋白饮食可提高肝脏的免疫功能。 2、高维生素饮食,维生素有营养、保护、支持肝细胞作用,新鲜蔬菜、水果中含有丰富的维生素物质,应每天吃500克左右蔬菜,吃3~4种水果。少吃油炸、烧烤食物,不一次大量摄入鸡、肉、鱼、蛋、豆制食品,以免蛋白质摄入过多加重肝脏负担。 3、丹参有抗肝炎病毒,活血化淤,保肝护心作用,可用少量丹参、黄芪饮片泡水饮用,也可口服丹参片3片/次,每日三次。 4、合理膳食:宜高碳水化合物、高维生素、适量高蛋白质饮食。 5、适量饮水,以促进机体代谢及代谢废物的排泄。 6、多食富含甲硫氨基酸丰富的降脂食物,如小米、燕麦等粗粮、黑芝麻、黑木耳、油菜、菠菜、菜花、甜菜头、海米、海带、干贝、淡菜等食品可促进体内磷脂合成,协助肝细胞内脂肪的转变。 饮食禁忌:(1)绝对禁酒(2)忌食辛辣刺激食品。如尽量不食洋葱、蒜、姜、辣椒、胡椒、咖喱等;少食用肉汤、鸡汤、鱼汤等含氮浸出物高的食物。(3)控制食糖,各种甜食及高热量食物,如含糖量高的蔬菜、水果、粉条、巧克力、甜点心等。(4)少吃或不吃煎炸等油类含量高的食品。(5)忌食用动物油,少食植物油等,少吃动物内脏、肥肉等。(6)忌过量或不科学用保健食品 (二)糖代谢失调指导 (一)生活起居: 树立正确的进食观,热量摄入过多、营养过剩、肥胖、运动缺乏是导致糖尿病的重要原因。应注意合理膳食。 1、低糖饮食,限制食量,每日三餐以6~7分饱为宜,避免进食速度过快,不要吃的过饱。控制体重,加强运动,消耗体内过剩能量,每天做有氧运动40—60分钟,可分俩个阶段进行,参加适当体力劳动,适当的体力劳动及适量运动能促进糖吸收,减轻胰岛负担。 2、避免过度紧张、劳累,人体在紧张、劳累时,体内交感神经兴奋,胰岛α细胞分泌增加,
反式脂肪酸在体内如何代谢
反式脂肪酸在体内如何代谢 1、反式脂肪酸同顺式脂肪酸一样能作为能源同样会被氧化而供能; 2、反式脂肪酸的确会导致VDL(极低密度脂蛋白)/LDL(低密度脂蛋白)的水平,它在体内的积累是因为不能通过脂合成途径合成体内其他脂质。 什么是反式脂肪酸? 反式脂肪酸是一类不饱和脂肪酸,包含至少一个反式结构的双键。 反式脂肪酸的来源于食品工业加工产生“氢化油”中以及反刍动物体内。 在食品工业中,由于天然植物油的双键是“顺式”结构,这种油抗氧化能力差,不稳定,工业上将植物油氢化,在这个过程中,部分油脂异构化产生了“反式”双键。以rans 9-Elaidic Acid(t9一C18:1)为主。 反刍动物的油脂以及牛奶中也存在反式脂肪酸,这是由于反刍动物瘤胃中的微生物将脂肪酸氢化而产生。以trans 11.Vaccenic Acid(t11一C18:1)为主,也还有顺9,反11一共轭亚油酸(c9, t11一CLA)和反10,顺12一共轭亚油酸(t10,c12一CLA)。 反式脂肪酸会增加体内VDL/LDL的水平,易导致心血管疾病、肥胖、胰岛素抗性、糖尿病等。 共轭亚油酸也是一种反式脂肪酸,但共轭亚油酸却与其他反式脂肪酸不同,它具有抗癌、降脂、抗动脉粥样硬化等功能。 反式脂肪酸在体内如何被氧化?
饱和脂肪酸的β-氧化过程大致经过4个步骤,既脱氢、加水、再脱氢和硫解这四个步骤。 由于反式脂肪酸为不饱和脂肪酸,因此先讲单不饱和脂肪酸的β-氧化过程。 体内正常的不饱和脂肪酸的双键都是顺式的,它们活化后进入β-氧化时,生成3-顺烯脂酰CoA, 此时需要顺-3反-2异构酶催化使其生成2-反烯脂酰CoA以便进一步反应。2-反烯脂酰CoA加水 后生成D-β-羟脂酰CoA,需要β-羟脂酰CoA差向异构酶催化,使其由D-构型转变成L-构型,以 便再进行脱氧反应(只有L-β-羟脂酰CoA才能作为β-羟脂酰CoA脱氢酶的底物)。 下图为多不饱和脂肪酸氧化示意图: 从不饱和脂肪酸的β-氧化过程可以看出,其“顺式”双键需要首先经过异构酶的催化变成“反式”双键才能进行 下一步氧化反应,而反式脂肪酸的氧化过程则不需要经过顺-3反-2异构酶的催化,直接完成加水、脱氢和硫解过程。 反式脂肪酸在体内的积累和对VDL/LDL水平的影响 体内的脂质作为前体能合成其他多不饱和脂肪酸,该过程需要脂肪酸去饱和酶的参与,但是该类酶 的底物为顺式双键,含有反式双键的脂肪酸则不能被延长或去饱和而被积累下来。
人体脂肪代谢的调控和调动
人体脂肪代谢的调控和调动 人体摄入的大部分)脂肪经胆汁乳化成小颗粒,胰腺和小肠内分泌的脂肪酶将脂肪里的脂肪酸水解成游离脂肪酸和甘油单酯(偶尔也有完全水解成甘油和脂肪酸). 水解后的小分子,如甘油、短链和中链脂肪酸,被小肠吸收进入血液。甘油单脂和长链脂肪酸被吸收后,先在小肠细胞中重新合成甘油三酯,并和磷脂、胆固醇和蛋白质形成乳糜微粒(chylomicron),由淋巴系统进入血液循环。 脂肪细胞在体内的代谢过程受到多种因素的调控,脂蛋白脂酶,以及脂肪细胞膜上的肾上腺素能受体、胰岛素受体及其他肽类激素和腺苷受体都参与这一过程的调节。 (1)脂蛋白脂酶(LPL):脂蛋白脂酶由体内脂肪细胞合成,然后释放到血液中附着在毛细血管的表面。其功能是将与其接触的乳糜微粒和极低密度脂蛋白中的三酰甘油(甘油三酯)水解成游离脂肪酸和α-磷酸甘油。前者进入脂肪细胞内,与磷酸甘油结合生成三酰甘油。由于人类脂肪细胞合成脂肪酸的能力很弱,因此在脂蛋白脂酶作用下所产生的游离脂肪酸就成为体内脂肪细胞合成三酰甘油所需要游离脂肪酸的主要来源。因此脂蛋白脂酶在调节人体局部脂肪沉积上发挥着一定的功能。脂蛋白脂酶的活性受机体营养状况及相关激素的调节,空腹及营养不良时其活性降低,进食后其活性增高。胰岛素可以增加脂蛋白脂酶的合成,而脂解激素则使脂蛋白脂酶活性受到抑制。 (2)胰岛素:胰岛素可以通过降低脂肪细胞内cAMP的浓度来抑制三酰甘油脂肪酶活性,减少三酰甘油的水解,促进水解后的游离脂肪酸再酯化。胰岛素是体内主要的抗脂解激素。当胰岛,素水平下降时,体内脂肪组织的脂解过程加快,血中游离脂肪酸和磷酸甘油浓度增高。 (3)儿茶酚胺:人类脂肪细胞上分布着许多α2和β1,受体,儿茶酚胺主要就是通过脂肪细胞膜上的肾上腺素能受体来调节脂解反应。 儿茶酚胺通过。α2受体抑制脂解,通过β1受体刺激脂解。人体不同部位脂肪细胞对儿茶酚胺的反应性是不相同的。无论男女,腹部脂肪细胞对儿茶酚胺促进脂解的反应性和敏感性均强于股部,绝经前女性股部脂肪细胞对儿茶酚胺的脂解反应性明显下降,而妊娠晚期和哺乳期女性股部脂肪细胞对儿茶酚胺的脂解反应性明显增强。造成上述差别的主要原因可能与分布在这些部位脂肪细胞上的。α2和β1受体的数目、比例及活性不同有关。 (4)性激素:性激素在促进脂肪细胞脂解反应区域性差异的发生上起着一定的作用。女性激素可以促进脂肪细胞α2受体的活性来达到拮抗儿茶酚胺的脂解作用。 (5)其他激素:生长激素、促肾上腺皮质激素、促甲状腺激素、泌乳素、胰高血糖素等均可促进脂肪细胞的脂解反应。 肪细胞的代谢过程是怎样进行的? 体内脂肪细胞的代谢过程是一个非常活跃、从不间断的循环过程。 正常情况下,机体内的脂肪细胞一方面不断地从血液中摄取食物分解后产生的游离脂肪酸,然后在细胞内将游离脂肪酸与由葡萄糖合成的。α-磷酸甘油结合生成磷酸三酰甘油。
糖脂代谢稳态调控的分子机制
项目名称:糖脂代谢稳态调控的分子机制首席科学家:林圣彩厦门大学 起止年限:2011.1至2015.8 依托部门:教育部
二、预期目标 1. 总体目标 确定机体和细胞在不同生理状况和环境因素下维持糖脂代谢稳态的分子机制,阐明在细胞生长和应激反应中起重要作用的调节因子调控细胞代谢的信号通路网络,为糖脂代谢紊乱造成的肥胖、脂肪肝、糖尿病和癌症的早期诊断和治疗提供理论依据。 2. 五年预期目标 (1) 建立对实验动物代谢相关的生理生化指标分析的技术平台,发现相关基因敲 除或转基因小鼠造成糖脂代谢紊乱的信号通路。 (2) 较系统地描述在逆境下机体和细胞调控糖脂代谢的分子网络以及调控过程 中关键蛋白质和蛋白质复合体的动态调控机制。 (3) 发现新的参与代谢调控的基因,为代谢性疾病和肿瘤的防治提供新的分子靶 标。 (4) 培养高质量博士研究生20-30名,培养3-5名享有国际知名度的专家和5-8名 中青年学术带头人。 (5) 在国际重要刊物发表SCI论文15-25篇,其中争取在Cell、Nature、Science或其 子刊等影响因子10以上杂志发表研究论文5-10篇,申请发明专利3-5项。
三、研究方案 1. 总体研究方案 细胞能量代谢是细胞最基本、最重要的活动之一,与细胞的繁殖、分化、凋亡、运动、信号转导及多种重要疾病的发生密切相关,是生命科学的一个重要领域。细胞要通过能量感应系统随时监测其能量水平状态,在不同的物质和能量状态下要不断地通过细胞内的代谢调控途径来调节其代谢水平以达到一种稳态。同时,细胞在面对内外界一些不良因素时也会做出相应的代谢变化,这些应激反应对细胞正常的生长和功能是极其重要的。如果这些应激反应失调,就会使细胞代谢发生异变,导致如前所述的多种人类重大疾病的发生。本项目的总体研究方案拟利用我们在蛋白质科学、细胞代谢、细胞信号转导等研究领域的研究优势和技术手段,结合细胞生物学、动物生理学等学科的研究方法,集中力量多层次、多角度地研究与细胞代谢调控相关的信号通路网络,分离和鉴定参与细胞代谢调控的新的基因和信号通路,探讨各个信号通路之间的动态调控机制,并研究细胞异常代谢的信号通路,揭示代谢异常与糖尿病、肿瘤等重大疾病的关系。项目总体研究方案如下图1:
生物化学代谢复习之糖代谢、脂质代谢
一、糖代谢 (一)糖的无氧氧化 1.基本概念糖酵解:一分子葡萄糖在胞质中可裂解生成两分子丙酮酸的过程称之为糖酵解,是葡萄糖无氧氧化和有氧氧化的共同起始途径。 糖的无氧氧化:在不能利用氧或氧供应不足时,机体分解葡萄糖生成乳酸的过程称为糖的无氧氧化,也称为乳酸发酵。 2.糖酵解的基本过程①葡萄糖在己糖激酶的催化下消耗1分子ATP生成葡糖-6-磷酸。②葡糖-6-磷酸异构为果糖-6-磷酸。 ③果糖-6-磷酸在磷酸果糖激酶-1的催化下消耗1分子的ATP生成果糖-1,6-二磷酸。 ④果糖-1,6-二磷酸在醛缩酶的催化下裂解为1分子磷酸二羟丙酮和1分子3-磷酸甘油醛。⑤磷酸二羟丙酮异构为3-磷酸甘油醛。(前面的步骤相当于1分子葡萄糖裂解产生了2分子3-磷酸甘油醛) ⑥3-磷酸甘油醛在3-磷酸甘油醛脱氢酶的催化下与1分子无机磷酸结合,脱下的氢由NAD+携带,生成1,3-二磷酸甘油酸(高能化合物)。⑦1,3-二磷酸甘油酸在磷酸甘油酸激酶的催化下水解高能磷酸键(底物水平磷酸化),产生ATP,生成3-磷酸甘油酸。⑧3-磷酸甘油酸变位为2-磷酸甘油酸。⑨2-磷酸甘油酸脱水生成磷酸烯醇式丙酮酸(高能化合物) 。⑩磷酸烯醇式丙酮酸在丙酮酸激酶的催化下生成丙酮酸,产生1分子A TP(底物水平磷酸化)。 该过程需要关注的几点:(1)三个限速反应:①③⑩,同时催化这三个反应的酶为关键酶(己糖激酶、磷酸果糖激酶-1、丙酮酸激酶) (2)该过程有两次底物水平磷酸化,包含了两个高能化合物(3)调节糖酵解流量最关键的酶是磷酸果糖激酶-1 (4)能量的产生与消耗 思考:1.1分子葡萄糖完全分解产生2分子丙酮酸可以产生多少个ATP? 2.糖原分子中葡萄糖酵解时可以净产生多少个ATP? 3.丙酮酸在在乳酸脱氢酶的作用下,由NADH+H+提供氢,使丙酮酸还原为乳酸 4.糖的无氧氧化的生理意义:①迅速提供能量,这对肌肉收缩很重要②成熟红细胞没有线粒体,只能依赖无氧氧化③神经细胞、白细胞、骨髓细胞等代谢极为活跃,即使不缺氧也常由糖的无氧氧化提供部分能量 (二)糖的有氧氧化 1.基本概念糖的有氧氧化是指机体利用氧将葡萄糖彻底氧化为CO2和H2O的反应过程。这个过程是体内糖分解供能的主要方式。 2.糖的有氧氧化的三个阶段 (1)同糖酵解(2)丙酮酸进入线粒体,丙酮酸在丙酮酸脱氢酶复合体(由转乙酰酶、二氢硫辛酸胺脱氢酶、丙酮酸脱氢酶组成)的催化下与辅酶A反应氧化脱羧,脱下的氢由NAD+携带,生成乙酰CoA和CO2。(参与的辅酶有TPP、硫辛酸、FAD、NAD+、CoA) (3)三羧酸循环(柠檬酸循环) ①乙酰CoA与草酰乙酸在柠檬酸合酶的催化下生成柠檬酸,反应所需的能量来自乙酰CoA。 ②柠檬酸经酶-顺乌头酸复合体异构为异柠檬酸。③异柠檬酸在异柠檬酸脱氢酶的催化下氧化脱羧,脱下的氢由NAD+携带,反应生成α-酮戊二酸及CO2。 ④α-酮戊二酸在α-酮戊二酸脱氢酶复合体的催化下与辅酶A反应氧化脱羧,脱下的氢由NAD+携带,反应生成琥珀酰CoA及CO2。 ⑤琥珀酰CoA在琥珀酰CoA合成酶的催化下水解掉高能硫酯键,与GDP磷酸化偶联,生成琥珀酸、GTP及CoA。 ⑥琥珀酸在琥珀酸脱氢酶的催化下生成延胡索酸,脱下的氢由FAD携带。 ⑦延胡索酸加水生成苹果酸。 ⑧苹果酸在苹果酸脱氢酶的催化下生成草酰乙酸,脱下的氢由NAD+携带。 该过程需要关注的几点:(1)三个限速反应:①③④,同时催化这三个反应的酶为关键酶(柠檬酸合酶、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶复合体)丙酮酸脱氢酶复合体也是关键酶(2)该过程只有一步水平磷酸化,只有一个高能化合物(当然乙酰CoA也是高能化合物) (3)生成三个NADH+H+和一个FADH2 (4)两次氧化脱羧(5)能量的产生与消耗 思考:1分子葡萄糖完全分解生成CO2和H2O可以产生多少ATP?(两种情况均思考)
雌激素通过瘦素信号通路途径调节脂肪细胞代谢生成
雌激素通过瘦素信号通路途径调节脂肪细胞代谢生成
雌激素通过瘦素信号通路途径调节脂肪细 胞代谢生成# 李文娟,许良智,陈焱,牟丽,许文明,程萌,庄静,李婷婷,詹晶**
10 15 20 25 30 35 40
(四川大学华西第二医院,成都 610041) 摘要:目的:探讨雌激素是否是通过瘦素相关信号通路对女性形体改变产生影响。方法:二 月龄雌性 SD 大鼠随机为去势组及假手术组,术后 14 周收集生殖器周围脂肪、内脏脂肪和 皮下脂肪,并分别检测瘦素受体表达,同时通过 17-β雌二醇及瘦素对脂肪细胞前体细胞 MSCs 进行干预,检验瘦素受体亚型、瘦素表达及成脂分化的指标 PPARγ的变化。结果: 通过对造模期间大鼠体重的每周监测,发现去势组体重增长及术后 14 周Lee’s 指数均明显 高于假手术组 P 0.001 。瘦素受体在去势组的脂肪组织中表达显著增加,内脏脂肪中尤为明 显。体外实验显示,随着瘦素和雌激素浓度的增加,MSCs 上瘦素长形受体和短受体的表达 均随之下降;随雌激素浓度的增加,MSCs 中瘦素表达呈下降趋势,同时,MSCs 中 PPAR γ表达也受到抑制。结论:在低雌激素的影响下,去势后大鼠发生类似绝经后女性样的形体 改变,高浓度雌激素可抑制大鼠间充质干细胞向脂肪细胞分化,雌激素对瘦素及瘦素受体的 影响可能是绝经后女性体型变化发生变化的原因。
关键词:妇产科学;雌激素;瘦素;瘦素受;脂肪;间充质干细胞 中图分类号:R339.6 Estrogen regulate adipocyte metabolism through leptin signaling pathway LI Wenjuan, XU Liangzhi, CHEN Yan, MU Li, XU Wenming, CHENG Meng, ZHUANG Jing, LI Tingting, ZHAN Jing West China Second University Hospital, Sichuan University, Chengdu 610041 Abstract: Postmenopausal women often present obvious body composition changes under the absence of estrogen, including overweight, obesity and android-like body fat distribution, therefore poses serious threaten for women’s health. Although the intimate relationship between estrogen and body appearance have been noticed, mechanism remains unclear. We assumed that estrogen may regulate fat distribution through affecting leptin signal pathway, which has been shown playing major role in energy homeostasis. To test this hypothesis, we randomized female SD rat into ovariectomy OVX and sham group, and then collected adipose tissue around genital,
项目名称-糖脂代谢稳态调控的分子机制-首席科学家-林圣彩厦门大学-
项目名称-糖脂代谢稳态调控的分子机制-首席科学家-林圣 彩厦门大学- 项目名称: 糖脂代谢稳态调控的分子机制首席科学家: 林圣彩厦门大学 起止年限: 2011.1至2015.8 依托部门: 教育部 二、预期目标 1. 总体目标 确定机体和细胞在不同生理状况和环境因素下维持糖脂代谢稳态的分子机制~阐明在细胞生长和应激反应中起重要作用的调节因子调控细胞代谢的信号通路网络~为糖脂代谢紊乱造成的肥胖、脂肪肝、糖尿病和癌症的早期诊断和治疗提供理论依据。 2. 五年预期目标 (1) 建立对实验动物代谢相关的生理生化指标分析的技术平台~发现相关基因敲 除或转基因小鼠造成糖脂代谢紊乱的信号通路。 (2) 较系统地描述在逆境下机体和细胞调控糖脂代谢的分子网络以及调控过程 中关键蛋白质和蛋白质复合体的动态调控机制。 (3) 发现新的参与代谢调控的基因~为代谢性疾病和肿瘤的防治提供新的分子靶 标。 (4) 培养高质量博士研究生20-30名~培养3-5名享有国际知名度的专家和 5-8名 中青年学术带头人。
(5) 在国际重要刊物发表SCI论文15-25篇~其中争取在Cell、Nature、Science或其 子刊等影响因子10以上杂志发表研究论文5-10篇~申请发明专利3-5项。 三、研究方案 1. 总体研究方案 细胞能量代谢是细胞最基本、最重要的活动之一~与细胞的繁殖、分化、凋亡、运动、信号转导及多种重要疾病的发生密切相关~是生命科学的一个重要领域。细胞要通过能量感应系统随时监测其能量水平状态~在不同的物质和能量状态下要不断地通过细胞内的代谢调控途径来调节其代谢水平以达到一种稳态。同时~细胞在面对内外界一些不良因素时也会做出相应的代谢变化~这些应激反应对细胞正常的生长和功能是极其重要的。如果这些应激反应失调~就会使细胞代谢发生异变~导致如前所述的多种人类重大疾病的发生。本项目的总体研究方案拟利用我们在蛋白质科学、细胞代谢、细胞信号转导等研究领域的研究优势和技术手段~结合细胞生物学、动物生理学等学科的研究方法~集中力量多层次、多角度地研究与细胞代谢调控相关的信号通路网络~分离和鉴定参与细胞代谢调控的新的基因和信号通路~探讨各个信号通路之间的动态调控机制~并研究细胞异常代谢的信号通路~揭示代谢异常与糖尿病、肿瘤等重大疾病的关系。项目总体研究方案如下图1: 内外环境因素(缺氧、营养缺乏或过剩、癌基因突变等)内外环境因素(缺氧、营养缺乏或过剩、癌基因突变等)
猪脂肪代谢的整个网络
猪脂肪代谢的整个网络(消化---吸收---转运---贮存---分解合成) (一)脂肪的消化 猪的口腔和胃几乎不消化脂肪。脂类到达十二指肠后,在肠蠕动的作用下与胰液和胆汁混合,胆汁中的胆汁酸盐使脂肪乳化并形成水包油的小胶体颗粒,以便于脂肪和胰液在油水界面充分接触,脂肪被充分的消化。胰液中含多种消化脂肪的酶类,包括胰脂肪酶,辅脂酶,胆固醇酯酶和磷脂酶A2等。胰脂肪酶在辅酯酶的协助下将甘油三酯水解为甘油二酯和甘油一酯;磷脂酶A2将饲粮磷脂水解为溶血磷脂和脂肪酸;胆固醇酯酶将胆固醇酯水解为胆固醇和游离脂肪酸。 (二)脂肪的吸收 脂肪消化产物在十二指肠下段和空场被吸收。甘油和短中链脂肪酸直接经小肠粘膜细胞吸收入门静脉血液,而长链脂肪酸和2-甘油一酯以混合微粒到达小肠粘膜细胞被吸收,随后在黏膜细胞中转化为甘油三酯,磷脂,胆固醇酯及少量胆固醇,再与黏膜细胞内合成的载脂蛋白一起形成能溶于水的乳糜微粒,乳糜微粒以胞饮作用的逆过程溢出黏膜细胞,经细胞间隙进入乳糜管,再经淋巴系统进入血液,然后由血管内皮细胞的脂蛋白酶水解为游离脂肪酸和甘油而被组织利用。 (三)脂类的转运 血浆中所含的脂类叫做血脂,包括甘油三酯,磷脂,胆固醇及其酯和游离脂肪酸。 游离脂肪酸是与血浆清蛋白结合形成可溶性复合物运输,其他血浆中的脂类常与肝脏和小肠粘膜细胞合成的载脂蛋白结合形成脂蛋白,并以脂蛋白的形式运输。脂蛋白酯酶LPL对其运输有重要的调节作用,LPL活性的高低是脂肪蓄积程度的标志,也是决定脂肪细胞大小的重要因素。血浆中各种脂蛋白的基本结构基本相似,由疏水性较强的甘油三酯和胆固醇酯形成的内核和由双极性分子以单分子层形式形成的表层结构。根据不同脂蛋白所含脂类,蛋白质的多少,用超速离心法将脂蛋白分为五类,即乳糜微粒,极低密度脂蛋白,中密度脂蛋白,低密度脂蛋白和高密度脂蛋白。 血中脂肪转运到脂肪组织,肌肉,乳腺等的毛细血管后,游离脂肪酸通过被动扩散进入细胞内,甘油三酯经毛细血管壁中的酶分解成游离脂肪酸后再被吸收,未被细胞吸收的物质经血液循环回至肝脏代谢。 (四)脂类的代谢 动物体内脂类的代谢受饲粮营养物质含量的影响,超过需要时多余的营养物质转变成脂肪沉积在脂肪组织中,低于需要时分解体脂肪供能。 1肝脏中脂肪的代谢:肝脏是脂肪合成,运转和利用的主要器官,进入肝脏的脂肪来自于消化道的吸收,少部分来自体内脂肪组织。肝细胞可氧化游离脂肪酸 转变为能量,供细胞代谢利用。饲料来源的游离脂肪酸和内源性脂肪酸都可被 肝细胞摄取并重新合成甘油三酯,新合成的甘油三酯很快以脂蛋白的形式释放 入血液,少部分的内源性脂肪酸还可合成磷脂和胆固醇。多余的沉积在肝中。 2脂肪组织中脂肪的代谢:猪和反刍动物在脂肪组织中进行脂肪代谢,脂肪组织也是储存脂肪的主要部位,沉积在脂肪组织的甘油酯是预先合成的,或是在机 体内脂酰辅酶A和L-3-磷酸甘油反应合成。 脂酰辅酶A的合成:一是在细胞液中由乙酰辅酶A或丁酰辅酶A合成棕榈酸, 棕榈酸可转化成几乎所有其他脂肪酸。二是主要在内质网也有少量在线粒体中 合成丙二酰CoA,丙二酰辅酶A与酰基载体蛋白(ACP)结合形成丙二酰ACP复 合物,乙酰辅酶A与ACP结合,使脂肪酸连接上两个碳原子而延长,最终形成 棕榈酸ACP复合物。三是仅在内质网中进行,其作用是使饱和脂肪酸去饱和,
脂肪代谢 课堂小结
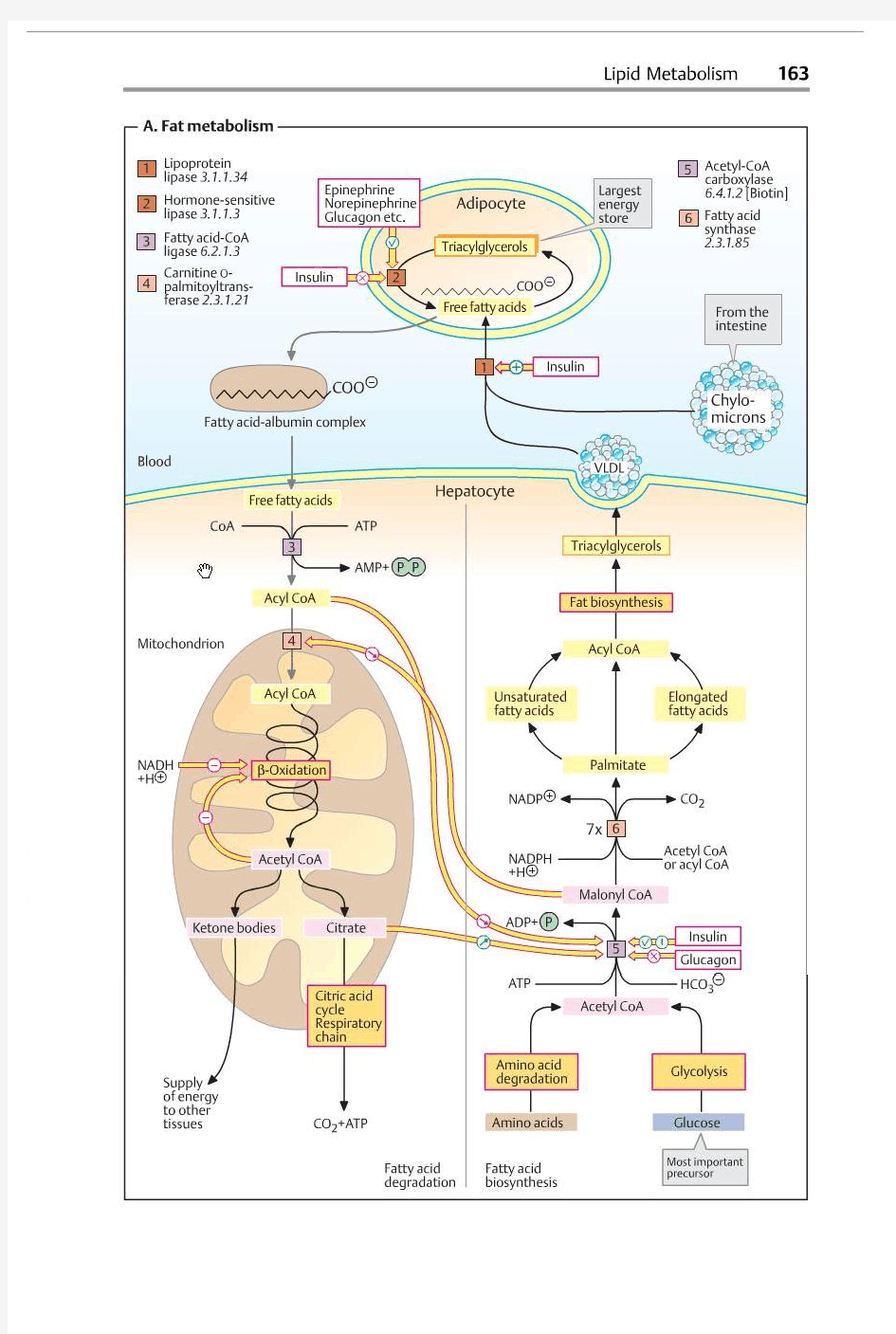
脂肪代谢 1 脂类在机体内的消化和吸收 消化主要在小肠上段经各种酶及胆汁酸盐的作用,水解为甘油、脂肪酸等。短链、中链脂肪酸甘油酯直接吸收,经门静脉入血;长链脂肪酸甘油酯与载脂蛋白、胆固醇等结合成乳糜微粒,经淋巴入血。 Diet triacylglycerols are emulsified and absorbed by the intestine 1) Bile salts, synthesized from cholesterol in liver, emulsifies macroscopic fat particles into microscopic mixed micelles for better lipase action and absorption. 2) Fatty acids generated from triacylglycerol (catalyzed by the intestinal lipase) diffuse into intestinal epithelial cells, be reconverted into triacylglycerol, and packed with cholesterol esters and specific apolipoproteins in chylomicrons。 3) Triacylglycerols are converted into fatty acids and glycerols in the capillaries by the action of lipoprotein lipases activated by apoC-II on chylomicrons, which in turn are absorbed mainly by adipocytes and myocytes for storage and energy consumption. 4) The leftover of the chylomicrons (containing mainly cholesterol and apolipoproteins) will be taken up by the liver by endocytosis; triacylglycerols will be used as the energy source for the liver cells, converted to ketone bodies or transported to adipose tissues after being packed with apolipoproteins. 2 甘油三酯代谢 (1) 合成代谢 甘油三酯是机体储存能量及氧化供能的重要形式。 1) 合成部位及原料 肝、脂肪组织、小肠是合成的重要场所,以肝的合成能力最强,注意:肝细胞能合成脂肪,但不能储存脂肪。合成后要与载脂蛋白、胆固醇等结合成极低密度脂蛋白,入血运到肝外组织储存或加以利用。若肝合成的甘油三酯不能及时转运,会形成脂肪肝。脂肪细胞是机体合成及储存脂肪的仓库。 合成甘油三酯所需的甘油及脂肪酸主要由葡萄糖代谢提供。其中甘油由糖酵解生成的磷酸二羟丙酮转化而成,脂肪酸由糖氧化分解生成的乙酰CoA合成。 2) 合成基本过程 ①甘油一酯途径:这是小肠粘膜细胞合成脂肪的途径,由甘油一酯和脂肪酸合成甘油三酯。 ②甘油二酯途径:肝细胞和脂肪细胞的合成途径。 脂肪细胞缺乏甘油激酶因而不能利用游离甘油,只能利用葡萄糖代谢提供的3-磷酸甘油。 (2) 分解代谢 即为脂肪动员,在脂肪细胞内激素敏感性甘油三酯脂的酶作用下,将脂肪分解为脂肪酸及甘油并释放入血供其他组织氧化。 甘油甘油激酶→3-磷酸甘油→磷酸二羟丙酮→糖酵解或有氧氧化供能,也可转变成糖脂肪酸与清蛋白结合转运入各组织经β-氧化供能。
脂肪代谢总结
脂类代谢 一、脂肪=甘油+脂肪酸 二、脂肪的降解 脂肪脂肪酶 甘 油 激 酶 α- (一)脂肪酸的氧化分解 包括:α、β(重点)、ω氧化 1、脂肪酸的活化以及转运 细胞定位:活化:——细胞质 转运:——从细胞质→线粒体内膜→线粒体基质 (1)活化 脂肪酸脂酰-CoA合成酶脂酰-CoA (2)转运 【注意】:肉碱脂酰基转移酶Ⅰ是β氧化的限速酶 2、脂肪酸的β氧化 细胞定位:线粒体基质 (1)饱和、偶数碳脂肪酸的β氧化 脂酰-CoA 脂酰-CoA脱氢酶烯脂酰-CoA 烯脂酰-CoA水合酶L-?-羟脂酰CoA H+ 脂酰-CoA(-2C) ?-酮脂酰-CoA 乙酰-CoA (2)不饱和脂肪酸的氧化 1.1单不饱和脂肪酸的氧化 特殊的酶:烯酰-CoA顺反异构酶(只有当底物是反式的时候β氧化第二步的水合酶才能够识
别) 1.2多不饱和脂肪酸的氧化 特殊的酶:烯酰-CoA顺反异构酶 二烯酰-CoA还原酶(减少一个双键,并且消耗2.5ATP) 烯酰-CoA异构酶(移动双键位置) (3)奇数碳脂肪酸的氧化 最后生成的丙酰-CoA可转化为琥珀酰-CoA 3、脂肪酸的α-氧化作用 概念:脂肪酸在一些酶的催化下,其α–碳原子也可发生氧化,结果生成一分子二氧化碳和比原来少一个碳原子的脂肪酸,这种氧化作用称为脂肪酸的α-氧化作用。 底物:奇数碳脂肪酸、支链脂肪酸、或过长的C22、C24 等长链脂肪酸 4、脂肪酸的ω氧化途径 概念:在酶的催化下,脂肪酸的烷基端碳,即远离羧基的末断碳原子(ω–碳原子)发生氧化,生成α、ω-二羧酸。脂肪酸的这种氧化作用称ω–氧化作用。 底物:动物体内10或12碳脂肪酸 (二)乙醛酸循环 生物学意义:是连接糖脂代谢的枢纽 关键酶:异柠檬酸裂解酶、苹果酸合酶 (三)酮体的生成与利用 丙酮 酮体乙酰乙酸 β-羟基丁酸 (四)磷脂代谢 磷脂=溶血磷脂+脂肪酸 参与甘油磷脂代谢的酶有四种:磷脂酶A、B、C、D