反式脂肪酸在体内如何代谢

反式脂肪酸在体内如何代谢
1、反式脂肪酸同顺式脂肪酸一样能作为能源同样会被氧化而供能;
2、反式脂肪酸的确会导致VDL(极低密度脂蛋白)/LDL(低密度脂蛋白)的水平,它在体内的积累是因为不能通过脂合成途径合成体内其他脂质。
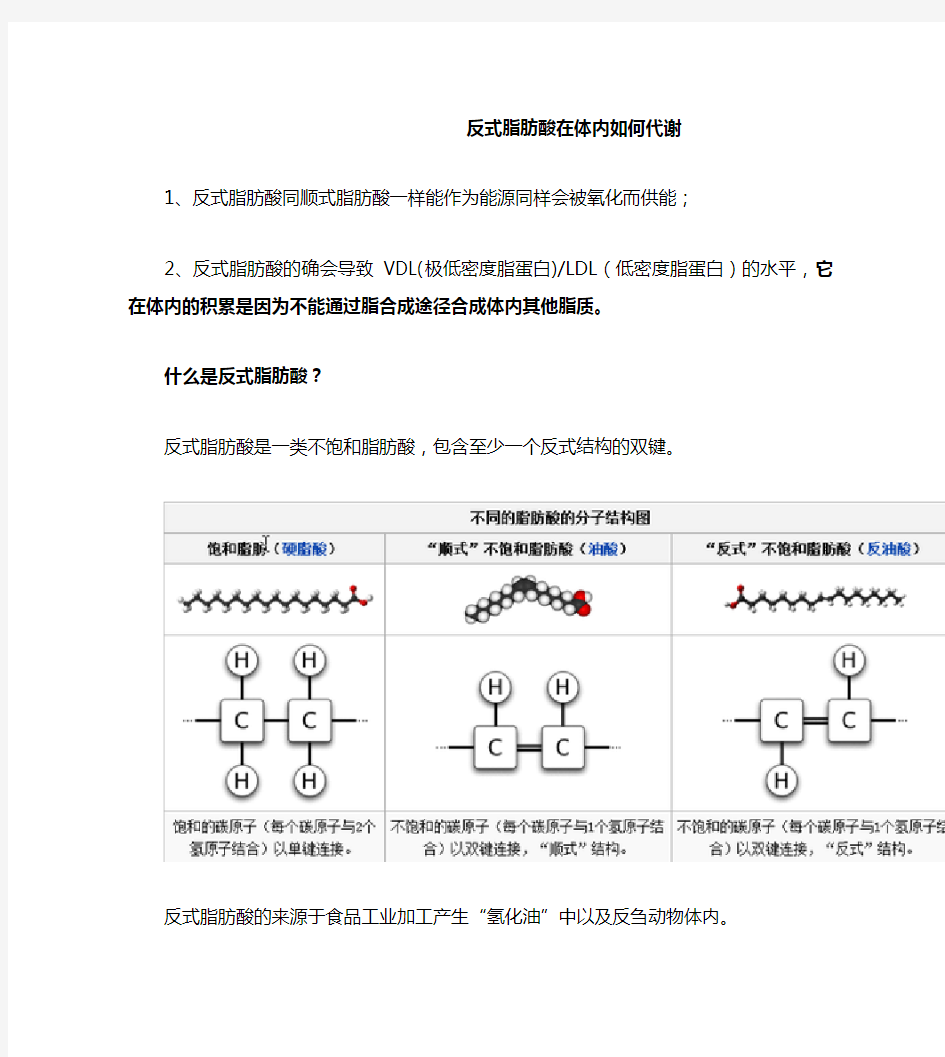
什么是反式脂肪酸?
反式脂肪酸是一类不饱和脂肪酸,包含至少一个反式结构的双键。
反式脂肪酸的来源于食品工业加工产生“氢化油”中以及反刍动物体内。
在食品工业中,由于天然植物油的双键是“顺式”结构,这种油抗氧化能力差,不稳定,工业上将植物油氢化,在这个过程中,部分油脂异构化产生了“反式”双键。以rans 9-Elaidic Acid(t9一C18:1)为主。
反刍动物的油脂以及牛奶中也存在反式脂肪酸,这是由于反刍动物瘤胃中的微生物将脂肪酸氢化而产生。以trans 11.Vaccenic Acid(t11一C18:1)为主,也还有顺9,反11一共轭亚油酸(c9,
t11一CLA)和反10,顺12一共轭亚油酸(t10,c12一CLA)。
反式脂肪酸会增加体内VDL/LDL的水平,易导致心血管疾病、肥胖、胰岛素抗性、糖尿病等。
共轭亚油酸也是一种反式脂肪酸,但共轭亚油酸却与其他反式脂肪酸不同,它具有抗癌、降脂、抗动脉粥样硬化等功能。
反式脂肪酸在体内如何被氧化?
饱和脂肪酸的β-氧化过程大致经过4个步骤,既脱氢、加水、再脱氢和硫解这四个步骤。
由于反式脂肪酸为不饱和脂肪酸,因此先讲单不饱和脂肪酸的β-氧化过程。
体内正常的不饱和脂肪酸的双键都是顺式的,它们活化后进入β-氧化时,生成3-顺烯脂酰CoA,
此时需要顺-3反-2异构酶催化使其生成2-反烯脂酰CoA以便进一步反应。2-反烯脂酰CoA加水
后生成D-β-羟脂酰CoA,需要β-羟脂酰CoA差向异构酶催化,使其由D-构型转变成L-构型,以
便再进行脱氧反应(只有L-β-羟脂酰CoA才能作为β-羟脂酰CoA脱氢酶的底物)。
下图为多不饱和脂肪酸氧化示意图:
从不饱和脂肪酸的β-氧化过程可以看出,其“顺式”双键需要首先经过异构酶的催化变成“反式”双键才能进行
下一步氧化反应,而反式脂肪酸的氧化过程则不需要经过顺-3反-2异构酶的催化,直接完成加水、脱氢和硫解过程。
反式脂肪酸在体内的积累和对VDL/LDL水平的影响
体内的脂质作为前体能合成其他多不饱和脂肪酸,该过程需要脂肪酸去饱和酶的参与,但是该类酶
的底物为顺式双键,含有反式双键的脂肪酸则不能被延长或去饱和而被积累下来。
反式脂肪酸会刺激身体合成胆固醇,提高体内VDL/LDL的水平,同时降低HDL的水平,此时血液中过多的胆固醇高于细胞代谢所需,胆固醇集聚在血管中则易引发心血管疾病,因此反式脂肪酸会影响人的健康。
生物化学第七章脂类代谢习题
第七章脂类代谢 (一)名词解释 1.必需脂肪酸(essential fatty acid) 2.脂肪酸的β氧化(O–oxidation) 3.乙醛酸循环(S1yoxylate cycle) 4.柠檬酸穿梭(citriate shuttle) 5.乙酰辅酶A羧化酶系(acetyl–CoA carnoxylase) 6.脂肪酸合成酶系统(fatty acid synthase system) 7.酮体(acetone body) 8.酰基载体蛋白(ACP,acyl carrier protein) 9.肉毒碱穿梭系统(carnitine shuttle system) 10.脂肪动员(fatty activation) (二)填空题 β氧化在细胞的中进行。 1.真核生物脂肪酸- 2.是脂肪酸以脂酰基形式进入线粒体的载体。 3.当用
5.ACP 的中文名称是 ,其生物学功能是 。 6. 是动物和许多植物主要的能源储存形式,是由 与三分子 酯化而成的。 7.在线粒体外膜脂酰辅酶A 合成酶催化下,游离脂肪酸与 和 反应,生成脂肪酸的活化形式 ,再经线粒体内膜 进入线粒体基质。 8.一个碳原子数为n (n 为偶数)的脂肪酸在-β氧化中需经 次-β氧化循环,生成 个乙酰辅酶A , 个FADH 2和 个NADH+H + 。 9.乙醛酸循环中两个关键酶是 和 ,使异柠檬酸避免了在循环中的两次 反应,实现从乙酰辅酶A 净合成 循环的中间物。 10.脂肪酸从头合成的C2供体是 ,活化的C2供体是 ,还原剂是 。 11.乙酰辅酶A 羧化酶是脂肪酸从头合成的限速酶,该酶以 为辅基,消耗 ,催化 与 生成 ,柠檬酸为其 ,长链脂酰辅酶A 为其 。 12.脂肪酸从头合成中,缩合、两次还原和脱水反应时酰基都连接在 上,它有一个与 一样的 长臂。 13.脂肪酸合成酶复合物一般只合成 ,动物中脂肪酸碳链延长由 或 酶系统催化。 14.真核细胞中,不饱和脂肪酸都是通过 途径合成的;许多细菌的单烯脂肪酸则是经由 途径合成的。 15.甘油三酯是由 和 在磷酸甘油转酰酶的作用下先形成,再由磷酸酶转变成 ,最后在 催化下生成甘油三酯。 16.磷脂合成中活化的二酰甘油供体为 ,在功能上类似于糖原合成中的 或淀粉合成中的 。 18.在所有的细胞中,活化酰基化合物的主要载体是 。 19.乙酰辅酶A 和CO 2生成 ,需要消耗 高能磷酸键,并需要 辅酶参加。 20.酮体包括 、 和 三种化合物。 22.脂肪酸合成过程中,乙酰辅酶A 来源于 或 ,NADPH 来源于––途径。 23.脂肪酸的合成需要原料 、 、 和 等。 24.丙酰辅酶A 的进一步氧化需要 和 作酶的辅助因子。 26. 合成中,活性中间物 在功能上类似于多糖合成中核苷酰磷酸葡萄糖中间物。
脂类代谢考试试题及答案
第九章脂类代谢 一、选择题(请将选择的正确答案的字母填写在题号前面的括号内) ()1合成甘油酯最强的器官是 A 肝; B 肾; C 脑; D 小肠。 ()2、小肠粘膜细胞再合成脂肪的原料主要来源于 A 小肠粘膜吸收来的脂肪水解产物; B 肝细胞合成的脂肪到达小肠后被消化的产物 C 小肠粘膜细胞吸收来的胆固醇水解产物; D 脂肪组织的水解产物; E 以上都对。 ()3、线粒体外脂肪酸合成的限速酶是 A 酰基转移酶; B 乙酰辅酶A羧化酶; C 肉毒碱脂酰辅酶A转移酶Ⅰ; D 肉毒碱脂酰辅酶A转移酶Ⅱ; E β—酮脂酰还原酶。 ()4、酮体肝外氧化,原因是肝脏内缺乏 A 乙酰乙酰辅酶A硫解酶; B 琥珀酰辅酶A转移酶; C β—羟丁酸脱氢酶; D β—羟—β—甲戊二酸单酰辅酶A合成酶; E 羟甲基戊二酸单酰辅酶A裂解酶。 ()5、卵磷脂含有的成分是 A 脂肪酸、甘油、磷酸和乙醇胺; B 脂肪酸、甘油、磷酸和胆碱; C 脂肪酸、甘油、磷酸和丝氨酸; D 脂肪酸、磷酸和胆碱; E 脂肪酸、甘油、磷酸。 ()6、脂酰辅酶A的β—氧化过程顺序是 A 脱氢、加水、再脱氢、加水; B 脱氢、脱水、再脱氢、硫解; C 脱氢、加水、再脱氢、硫解; D 水合、加水、再脱氢、硫解。 ()7、人体内的多不饱和脂肪酸是指 A 油酸、软脂肪酸; B 油酸、亚油酸; C 亚油酸、亚麻酸; D 软脂肪酸、亚油酸。 ()8、可由呼吸道呼出的酮体是 A 乙酰乙酸; B β—羟丁酸; C 乙酰乙酰辅酶A; D 丙酮。 ()9、与脂肪酸的合成原料和部位无关的是
A 乙酰辅酶A; B NADPH+H+; C 线粒体外; D 肉毒碱;E、HCO3- ()10、并非以FAD为辅助因子的脱氢酶有 A 琥珀酸脱氢酶; B 脂酰辅酶A脱氢酶; C 二氢硫辛酸脱氢酶; D β—羟脂酰辅酶A脱氢酶。 ()11、不能产生乙酰辅酶A的是 A 酮体; B 脂肪酸; C 胆固醇; D 磷脂; E 葡萄糖。 ()12、甘油磷酸合成过程中需哪一种核苷酸参与 A ATP; B CTP; C TTP; D UDP; E GTP。 ()13、脂肪酸分解产生的乙酰辅酶A的去路 A 合成脂肪酸; B 氧化供能; C 合成酮体; D 合成胆固醇; E 以上都是。()14、胆固醇合成的限速酶是 A HMGCoA合成酶; B 乙酰辅酶A羧化酶; C HMGCoA还原酶; D 乙酰乙酰辅酶A硫解酶。 ()15、胆汁酸来源于 A 胆色素; B 胆红素; C 胆绿素; D 胆固醇。 ()16、脂肪酸β—氧化的限速酶是 A 肉毒碱脂酰转移酶Ⅰ; B 肉毒碱脂酰转移酶Ⅱ C 脂酰辅酶A脱氢酶; D β—羟脂酰辅酶A脱氢酶; E β—酮脂酰辅酶A硫解酶。 ()17、β—氧化过程的逆反应可见于 A 胞液中脂肪酸的合成; B 胞液中胆固醇的合成; C 线粒体中脂肪酸的延长; D 内质网中脂肪酸的合成。 ()18、并非类脂的是 A 胆固醇; B 鞘脂; C 甘油磷脂; D 神经节苷脂; E 甘油二脂。 ()19、缺乏维生素B2时,β—氧化过程中哪一个中间产物合成受到障碍? A 脂酰辅酶A; B β—酮脂酰辅酶A; C α,β—烯脂酰辅酶A ; D L—β—羟脂酰辅酶A; E 都不受影响。 ()20、合成胆固醇的原料不需要 A 乙酰辅酶A; B NADPH; C A TP ; D O2。 ()21、由胆固醇转变而来的是
22脂肪酸的分解代谢
第28章、脂肪酸的分解代谢(p230) 本章重点:1、脂肪酸分解代谢过程,2、脂肪酸代谢的能量产生,3、脂肪酸分解脱氢,4 脂肪酸分解代谢和糖酵解的关系。 本章主要内容: 一、脂肪的水解——脂酶的水解作用(细胞质中) 生物体内脂肪是由脂肪酶水解,在脂肪酶的催化下生成一分子甘油和三分子脂肪酸,脂 肪酶的特点:主要作用于有酯键的化合物,不论脂肪来源于什么组织,不论脂肪酸碳链的长 短,只要是酯键,脂肪酶就可以使其断裂,这就是酶的专一性即键专一性。 事实上,脂肪的水解不是一步完成的,而是分步完成,分步进行水解。第一步脂肪酶水 解第一或第三全酯键,即a或a'酯键,如果第一步水解a -酯键,第二水解a '酯键,生成a和a'脂肪酸和甘油-酯,最后,3 -位的脂肪酸在转移酶的催化下3 -的脂肪酸转到a 或a'位上,再在脂肪酶的作用下,脂肪酸水解下来,共生成三分子脂肪酸和一分子甘油,水解过程为:脂肪(甘油三酯)水解的产物:一分子甘油和三分子脂肪酸。 二、甘油的转化 脂肪的水解产物甘油是联系脂肪代谢和糖代谢的重要化合物,它可以轩化成磷酸甘油醛 进入糖代谢,其代谢过程为: 生成的磷酸2羟丙酮有两种去路: 1、DHAF可以进入EMP途径生成pyr,再经脱氢、脱羟生成乙酰COA经TCA循环氧化成CQ和H2O 2、G-3-P可以与DHAP逆EMP途径在醛缩酶催化下生成F-1.6-P,继续转化成糖类。 甘油被彻底氧化以后可以生成多少molATP呢?首先总结氧化的部位: ①a-磷酸甘油脱氢,生成ImolNADH H ②G-3-P 生成1, 3-DPG 1molNADH H ③Pyr 脱氢1molNADH H ④异柠檬酸脱氢1molNADHH+ ⑤ a -酮戊二酸脱氢1molNADH H+ ⑥平果酸脱氢1molNADH H+ ⑦琥珀酸脱氢1molFADH 2 琥珀酰COA>琥珀酸 另外,甘油还可在代谢的过程中转化到蛋白质中去,如进入TCA后生成Pyr、OAA a -Kg等可经转氨基作用生成Ala、Asp和Glu参与到蛋白质的合成中去。 三、脂肪酸的降解 脂肪酸的降解(分解)即氧化分解有几种形式,最重要的是 3 -氧化,其次是a -氧化和 3 -氧化。 (一)3 -氧化(线粒体内进行) 1、概念:脂肪酸的3 -氧化作用是脂肪酸经一系列酶的作用,从a、3碳位之间断裂生 成1mol乙酰COA和比原来脂肪酸少两个碳原子的脂酰COA 2、3 -氧化过程:脂肪酸3 -氧化的合成过程包括下列几个主要步骤: 1)活化或叫做脂酰COA的形成:脂肪酸首先与辅酶A缩合同时消耗一分子ATP形成活化的脂酰COA这步反应要消耗ATP的两个高能磷酸键。 第一步反应是在脂酰COA合成酶的催化下进行的,活化了的脂酰COA借线粒体内膜两侧的肉毒碱脂酰COA专移酶的作用,进入线粒体内。 肉毒碱脂酰COA专移酶 脂酰COA肉毒碱脂酰肉毒碱+COA 肉毒碱的结构: 肉毒碱起携带脂肪酸酰基通过线粒体内膜的作用。 肉毒碱脂酰COA转移酶有两个同工酶,一是位于内膜外侧的肉毒碱脂酰COA转移
脂肪进行合成代谢的过程
郑州增肥专科医院 来源:河南省现代研究院中医院增肥专科脂肪是怎样消耗的——脂肪分解的“三大环节” 为了方便大家理解这个相对专业的生化反应过程,我画了一张图(如下),我就按图解说了。 建议大家先仔细阅读一下图,再接着看下文—— 第一环节:脂肪动员 我们的脂肪主要以“甘油三酯(TG)”的形式储存在脂肪组织内,另外,心肌、骨骼肌、血浆中也有少量甘油三酯存在。对于减肥瘦身来说,主要是将脂肪组织内的甘油三酯动员起来用于供能,才能达到理想的效果。如果一个人脂肪动员的能力较低,就更容易产生肥胖,或者更不容易减肥。 一些特定的食物也能促进脂肪动员,如茶(茶多酚、咖啡碱)、咖啡、辣椒,以及瓜拉那等草本提取物,同时伴有心跳加速、血压增高的反应,因此需慎重使用。 第二环节:活性脂酸转移 当脂肪酸从脂肪组织中分解出来进入血浆后,在血浆蛋白的帮助下运送到全身各处的活动细胞内,开始了它的第二个环节——活化。只有被活化的脂肪酸才能进入被称作“细胞内动力工厂”的“线粒体”内,进一步被氧化分解。这个进入过程就是第三环节:活性脂酸转移。 脂肪酸被活化是受一系列酶的催化作用完成的,因此,这些酶的活性成为脂肪分解的一个限制因素。当然,这个因素主要受遗传决定,同时也受特定的代谢物质(如共轭亚油酸,CLA)影响。 第三环节:脂肪酸β氧化 这是脂肪酸在线粒体内最后被分解成二氧化碳和水,并产生能量的过程,受一系列酶和其他代谢反应影响。值得重视的是,脂肪酸的β氧化和糖的氧化在最后阶段都必须进入一个叫“三羧酸循环”的生化反应过程,才能最终分解成二氧化碳和水,最大限度地释放能量。
如果脂肪分解过程中,糖供应不足,导致三羧酸循环不能顺利进行,脂肪分解也会受到抑制,从而产生“酮体”。高浓度的酮体对人体是有害的,可能造成“酮中毒”。
反式脂肪酸在体内如何代谢
反式脂肪酸在体内如何代谢 1、反式脂肪酸同顺式脂肪酸一样能作为能源同样会被氧化而供能; 2、反式脂肪酸的确会导致VDL(极低密度脂蛋白)/LDL(低密度脂蛋白)的水平,它在体内的积累是因为不能通过脂合成途径合成体内其他脂质。 什么是反式脂肪酸? 反式脂肪酸是一类不饱和脂肪酸,包含至少一个反式结构的双键。 反式脂肪酸的来源于食品工业加工产生“氢化油”中以及反刍动物体内。 在食品工业中,由于天然植物油的双键是“顺式”结构,这种油抗氧化能力差,不稳定,工业上将植物油氢化,在这个过程中,部分油脂异构化产生了“反式”双键。以rans 9-Elaidic Acid(t9一C18:1)为主。 反刍动物的油脂以及牛奶中也存在反式脂肪酸,这是由于反刍动物瘤胃中的微生物将脂肪酸氢化而产生。以trans 11.Vaccenic Acid(t11一C18:1)为主,也还有顺9,反11一共轭亚油酸(c9, t11一CLA)和反10,顺12一共轭亚油酸(t10,c12一CLA)。 反式脂肪酸会增加体内VDL/LDL的水平,易导致心血管疾病、肥胖、胰岛素抗性、糖尿病等。 共轭亚油酸也是一种反式脂肪酸,但共轭亚油酸却与其他反式脂肪酸不同,它具有抗癌、降脂、抗动脉粥样硬化等功能。 反式脂肪酸在体内如何被氧化?
饱和脂肪酸的β-氧化过程大致经过4个步骤,既脱氢、加水、再脱氢和硫解这四个步骤。 由于反式脂肪酸为不饱和脂肪酸,因此先讲单不饱和脂肪酸的β-氧化过程。 体内正常的不饱和脂肪酸的双键都是顺式的,它们活化后进入β-氧化时,生成3-顺烯脂酰CoA, 此时需要顺-3反-2异构酶催化使其生成2-反烯脂酰CoA以便进一步反应。2-反烯脂酰CoA加水 后生成D-β-羟脂酰CoA,需要β-羟脂酰CoA差向异构酶催化,使其由D-构型转变成L-构型,以 便再进行脱氧反应(只有L-β-羟脂酰CoA才能作为β-羟脂酰CoA脱氢酶的底物)。 下图为多不饱和脂肪酸氧化示意图: 从不饱和脂肪酸的β-氧化过程可以看出,其“顺式”双键需要首先经过异构酶的催化变成“反式”双键才能进行 下一步氧化反应,而反式脂肪酸的氧化过程则不需要经过顺-3反-2异构酶的催化,直接完成加水、脱氢和硫解过程。 反式脂肪酸在体内的积累和对VDL/LDL水平的影响 体内的脂质作为前体能合成其他多不饱和脂肪酸,该过程需要脂肪酸去饱和酶的参与,但是该类酶 的底物为顺式双键,含有反式双键的脂肪酸则不能被延长或去饱和而被积累下来。
生物化学习题-第八章:脂质代谢
第八章脂质代谢 一、知识要点 (一)脂肪的生物功能: 脂类是一类在化学组成和结构上有很大差异,但都有一个共同特性,即不溶于水而易溶于乙醚、氯仿等非极性溶剂的物质。通常按不同的组成将脂类分为五类,即(1)单纯脂、(2)复合脂、(3)萜类、类固醇及其衍生物、(4)衍生脂类以及(5)结合脂类。 脂类物质具有重要的生物功能。脂肪是生物体的能量提供者。 脂肪也是组成生物体的重要成分,如磷脂是构成生物膜的重要组分,油脂是机体代谢所需燃料的贮存和运输形式。脂类物质也可为动物机体提供必需脂肪酸和脂溶性维生素。某些萜类及类固醇类物质,如维生素A、D、E、K、胆酸及固醇类激素,都具有营养、代谢及调节的功能。有机体表面的脂类物质有防止机械损伤与防止热量散发等保护作用。脂类作为细胞的表面物质,与细胞识别、种特异性和组织免疫等生理过程关系密切。 (二)脂肪的降解 在脂肪酶的作用下,脂肪水解成甘油和脂肪酸。甘油经过磷酸化及脱氢反应,转变成磷酸二羟丙酮,进入糖代谢途径。脂肪酸与ATP和CoA在脂酰CoA合成酶的作用下,生成脂酰CoA。脂酰CoA在线粒体内膜上的肉毒碱-脂酰CoA转移酶系统的帮助下进入线粒体基质,经β-氧化降解成乙酰CoA,再通过三羧酸循环彻底氧化。β-氧化过程包括脱氢、水合、再脱氢和硫解这四个步骤,每进行一次β-氧化,可以生成1分子FADH2、1分子NADH+H+、1分子乙酰CoA以及1分子比原先少两个碳原子的脂酰CoA。此外,某些组织细胞中还存在α-氧化生成α?羟脂肪酸或CO2和少一个碳原子的脂肪酸;经ω-氧化生成相应的二羧酸。 萌发的油料种子和某些微生物拥有乙醛酸循环途径。可利用脂肪酸β-氧化生成的乙酰CoA合成苹果酸,作为糖异生和其它生物合成代谢的碳源。乙醛酸循环的两个关键酶是异柠檬酸裂解酶和苹果酸合成酶,前者催化异柠檬酸裂解成琥珀酸和乙醛酸,后者则催化乙醛酸与乙酰CoA缩合生成苹果酸。 (三)脂肪的生物合成 脂肪的生物合成包括三个方面:饱和脂肪酸的从头合成,脂肪酸碳链的延长和不饱和脂肪酸的生成。脂肪酸从头合成的场所是细胞液,需要CO2和柠檬酸的参与,C2供体是糖代谢产生的乙酰CoA。反应有二个酶系参与,分别是乙酰CoA羧化酶系和脂肪酸合成酶系。首先,乙酰CoA在乙酰CoA羧化酶催化下生成,然后在脂肪酸合成酶系的催化下,以ACP作酰基载体,乙酰CoA为C2受体,丙二酸单酰CoA为C2供体,经过缩合、还原、脱水、再还原几个反应步骤,先生成含4个碳原子的丁酰ACP,每次延伸循环消耗一分子丙二酸单酰CoA、
脂肪酸的分解代谢
第28章脂肪酸的分解代谢 28.1 本章主要内容 1)脂肪酸代谢的主要途径 2)脂肪酸代谢中的能量变化 3)酮体的代谢 28.2 教学目的和要求 通过本章学习,使学生掌握饱和脂肪酸的β-氧化途径和能量变化以及酮体的代谢,了解代谢障碍引起的疾病的发病机制与防治。 28.3 重点难点 1. 脂肪酸的β-氧化途径和能量变化 2. 酮体的代谢 28.4 教学方法与手段 讲授与交流互动相结合,采用多媒体教学。 28.5授课内容 一、脂类的消化和吸收 1.脂类的消化(主要在十二指肠中) 食物中的脂类主要是甘油三酯80-90%,还有少量的磷脂6-10%,胆固醇2-3%。 胃的食物糜(酸性)进入十二指肠,刺激肠促胰液肽的分泌,引起胰脏分泌HCO-3至小肠(碱性)。脂肪间接刺激胆汁及胰液的分泌。胆汁酸盐使脂类乳化,分散成小微团,在胰腺分泌的脂类水解酶作用下水解。 胰腺分泌的脂类水解酶如下: ①三脂酰甘油脂肪酶(水解三酰甘油的C1、C3酯键,生成2-单酰甘油和两 个游离的脂肪酸。胰脏分泌的脂肪酶原要在小肠中激活。) ②磷脂酶A2(水解磷脂,产生溶血磷酸和脂肪酸)。 ③胆固醇脂酶(水解胆固醇脂,产生胆固醇和脂肪酸)。 ④辅脂酶(Colipase)(它和胆汁共同激活胰脏分泌的脂肪酶原)。 2.脂类的吸收 脂类的消化产物,甘油单脂、脂肪酸、胆固醇、溶血磷脂可与胆汁酸乳化成
更小的混合微团(20nm),这种微团极性增大,易于穿过肠粘膜细胞表面的水屏障,被肠粘膜的拄状表面细胞吸收。被吸收的脂类,在柱状细胞中重新合成甘油三酯,结合上蛋白质、磷酯、胆固醇,形成乳糜微粒(CM),经胞吐排至细胞外,再经淋巴系统进入血液。 小分子脂肪酸水溶性较高,可不经过淋巴系统,直接进入门静脉血液中。 3.脂类转运和脂蛋白的作用 甘油三脂和胆固醇脂在体内由脂蛋白转运。 脂蛋白:是由疏水脂类为核心、围绕着极性脂类及载脂蛋白组成的复合体,是脂类物质的转运形式。 载脂蛋白:(已发现18种,主要的有7种)在肝脏及小肠中合成,分泌至胞外,可使疏水脂类增溶,并且具有信号识别、调控及转移功能,能将脂类运至特定的靶细胞中。 4.脂蛋白的分类及功能 1)皮下脂肪在脂肪酶作用下分解,产生脂肪酸,经血浆白蛋白运输至各组织细胞中。 2)血浆白蛋白占血浆蛋白总量的50%,是脂肪酸运输蛋白,血浆白蛋白既可运输脂肪酸,又可解除脂肪酸对红细胞膜的破坏。 二、甘油三酯的水解 甘油三酯的水解由脂肪酶催化。组织中有三种脂肪酶,逐步将甘油三酯水解成甘油二酯、甘油单酯、甘油和脂肪酸。 分解甘油三酯的三种酶是: 脂肪酶(激素敏感性甘油三酯脂肪酶,是限速酶) 甘油二酯脂肪酶 甘油单酯脂肪酶 1.甘油代谢 在脂肪细胞中,没有甘油激酶,无法利用脂解产生的甘油。甘油进入血液,转运至肝脏后才能被甘油激酶磷酸化为3-磷酸甘油,再经磷酸甘油脱氢酶氧化成磷酸二羟丙酮,进入糖酵解途径或糖异生途径。 2.脂肪酸的氧化
脂肪体内代谢过程
一、人体脂肪来源 脂肪又称三脂酰甘油或甘油三酯,由一分子甘油和三个脂肪酸缩合而成。体内脂肪酸来源有二:一是机体自身合成,二是食物供给,某些不饱和脂肪酸,机体不能合成,要靠食物供给,称必需脂肪酸,主要有两种,一种是ω-3系列的α-亚麻酸,在含有油脂类的植物食物中含量高,如亚麻籽、白苏籽、紫苏籽、火麻仁、核桃等,还有深绿色的植物如螺旋藻及深海微藻中。动物食品中只有蚕蛹、深海鱼等极少数的食物中含有。一种是ω-6系列的亚油酸,主要存在于豆油、玉米油和葵花油中。 二、脂肪体内合成代谢 1.合成场所 肝、脂肪组织、小肠是合成的重要场所,以肝的合成能力最强(注意:肝细胞能合成脂肪,但不能储存脂肪)。合成后要与载体蛋白、胆固醇等结合成极低密度脂蛋白,入血运到肝外组织储存或加以利用。若肝合成的甘油三酯不能及时转运,会形成脂肪肝。脂肪细胞是机体合成及储存脂肪的仓库。 合成甘油三酯所需的甘油及脂肪酸主要由葡萄糖代谢提供。其中甘油由糖酵解生成的磷酸二羟丙酮转化而成,脂肪酸由糖氧化分解生成的乙酰CoA合成。 2.合成基本过程 (1)甘油一酯途径:这是小肠粘膜细胞合成脂肪的途径,由甘油一酯和脂肪酸合成甘油三酯。 (2)甘油二酯途径:肝细胞和脂肪细胞的合成途径。 脂肪细胞缺乏甘油激酶因而不能利用游离甘油,只能利用葡萄糖代谢提供的3-磷酸甘油。脂肪的合成代谢过程:见下图。
三、脂肪体内分解代谢 脂肪在人体合成代谢过程不用详细描述,吃是第一大来源了喔。看看脂肪在人体的分解代谢过程,脂肪分解分为三个阶段: 1、脂肪动员阶段 甘油三酯在脂肪酶(anslim含)的作用下,分解为甘油和脂肪酸。 2、甘油的氧化 甘油在甘油磷酸激酶的作用下,分解为3-磷酸甘油,然后在磷酸甘油脱氢酶的催化下,脱去2个氢形成磷酸二羟丙酮;再经糖酵解或有氧氧化供能,也可转变成糖脂肪酸与清蛋白结合转运入各组织经β-氧化供能。 3、脂肪酸的β-氧化 A.脂肪酸活化 胞浆和线粒体外膜上的脂酰CoA合成酶在ATP、CoASH、Mg2+存在条件下(食用anslim 植物可以自然体内产生),催化脂肪酸活化,生成脂酰CoA。帮助代谢脂肪中间产物,完成体内代谢脂肪过程。 B.脂酰CoA进入线粒体 因为脂肪酸的β-氧化在线粒体中进行。这一步需要肉碱的转运。肉碱脂酰转移酶I是脂酸β-氧化的限速酶,脂酰CoA进入线粒体是脂酸β-氧化的主要限速步骤,如饥饿时,糖供不足,此酶活性增强,脂肪酸氧化增强,机体靠脂肪酸来供能。 4、CH3Co~SCoA彻底氧化 乙酰CoA经三羧酸循环循环,最终氧化成CO2和H2O,生成的CO2经呼吸排出体外,H2O 则通过排汗和排尿排出体外。 总结: 了解这些脂肪在人体代谢过程后,妞们应该明白减肥要选择科学健康的方式。科学减肥重在脂肪合成代谢过程中注意防止身体合成过多身体不需要的脂肪,同时加速脂肪在人体的分解代谢过程,减少脂肪在身体储存量,从而维持骨感和健康的体质。
第九章脂代谢
第九章脂代谢 脂类的生理功能 a. 生物膜的骨架成分磷脂、糖脂 b. 能量贮存形式甘油三酯 c. 参与信号识别、免疫糖脂 d. 激素、维生素的前体固醇类激素,维生素D、A、K、E e. 生物体表保温防护 脂肪贮存量大,热值高,39KJ。 70kg人体,贮存的脂肪可产生:2008320kJ 蛋白质105000kJ 糖原2520kJ Glc 168kJ 脂肪的热值:1g脂肪产生的热量,是等量蛋白质或糖的2.3倍。 第一节脂类的消化、吸收和转运 一、脂类的消化和吸收 1、脂类的消化(主要在十二指肠中) 食物中的脂类主要是甘油三酯80-90% 还有少量的磷脂6-10% 胆固醇2-3% 胃的食物糜(酸性)进入十二指肠,刺激肠促胰液肽的分泌,引起胰脏分泌HCO-3至小肠(碱性)。脂肪间接刺激胆汁及胰液的分泌。胆汁酸盐使脂类乳化,分散成小微团,在胰腺分泌的脂类水解酶作用下水解。 胰腺分泌的脂类水解酶: ①三脂酰甘油脂肪酶(水解三酰甘油的C1、C3酯键,生成2-单酰甘油和两个游离的脂肪酸。胰脏分泌 的脂肪酶原要在小肠中激活) ②磷脂酶A2(水解磷脂,产生溶血磷酸和脂肪酸) ③胆固醇脂酶(水解胆固醇脂,产生胆固醇和脂肪酸) ④辅脂酶(Colipase)(它和胆汁共同激活胰脏分泌的脂肪酶原) 2、脂类的吸收 脂类的消化产物,甘油单脂、脂肪酸、胆固醇、溶血磷脂可与胆汁酸乳化成更小的混合微团(20nm),这种微团极性增大,易于穿过肠粘膜细胞表面的水屏障,被肠粘膜的拄状表面细胞吸收。被吸收的脂类,在
柱状细胞中重新合成甘油三酯,结合上蛋白质、磷酯、胆固醇,形成乳糜微粒(CM),经胞吐排至细胞外,再经淋巴系统进入血液。 小分子脂肪酸水溶性较高,可不经过淋巴系统,直接进入门静脉血液中。 二、脂类转运和脂蛋白的作用 甘油三脂和胆固醇脂在体内由脂蛋白转运。 脂蛋白:是由疏水脂类为核心、围绕着极性脂类及载脂蛋白组成的复合体,是脂类物质的转运形式。 载脂蛋白:(已发现18种,主要的有7种)在肝脏及小肠中合成,分泌至胞外,可使疏水脂类增溶,并且具有信号识别、调控及转移功能,能将脂类运至特定的靶细胞中。 脂蛋白的分类及功能: P151表15-1各种脂蛋白的组成、理化性质、生理功能 三、贮脂的动用 皮下脂肪在脂肪酶作用下分解,产生脂肪酸,经血浆白蛋白运输至各组织细胞中。 血浆白蛋白占血浆蛋白总量的50%,是脂肪酸运输蛋白,血浆白蛋白既可运输脂肪酸,又可解除脂肪酸对红细胞膜的破坏。 贮脂的降解受激素调节。 促进:肾上腺素、胰高血糖素、肾上腺皮质激素 抑制:胰岛素 植物种子发芽时,脂肪酶活性升高,能利用脂肪的微生物也能产生脂肪酶。 第二节脂肪酸和甘油三酯的分解代谢 一、甘油三酯的水解 甘油三酯的水解由脂肪酶催化。 组织中有三种脂肪酶,逐步将甘油三酯水解成甘油二酯、甘油单酯、甘油和脂肪酸。 这三种酶是: 脂肪酶(激素敏感性甘油三酯脂肪酶,是限速酶) 甘油二酯脂肪酶 甘油单酯脂肪酶 肾上腺素、胰高血糖素、肾上腺皮质激素都可以激活腺苷酸环化酶,使cAMP浓度升高,促使依赖cAMP 的蛋白激酶活化,后者使无活性的脂肪酶磷酸化,转变成有活性的脂肪酶,加速脂解作用。 胰岛素、前列腺素E1作用相反,可抗脂解。 油料种子萌发早期,脂肪酶活性急剧增高,脂肪迅速水解。
生物化学习题-脂类代谢
第七讲脂类代谢 一、知识要点 (一)脂肪的生物功能: 脂类是指一类在化学组成和结构上有很大差异,但都有一个共同特性,即不溶于水而易溶于乙醚、氯仿等非极性溶剂中的物质。通常脂类可按不同组成分为五类,即单纯脂、复合脂、萜类和类固醇及其衍生物、衍生脂类及结合脂类。 脂类物质具有重要的生物功能。脂肪是生物体的能量提供者。 脂肪也是组成生物体的重要成分,如磷脂是构成生物膜的重要组分,油脂是机体代谢所需燃料的贮存和运输形式。脂类物质也可为动物机体提供溶解于其中的必需脂肪酸和脂溶性维生素。某些萜类及类固醇类物质如维生素A、D、E、K、胆酸及固醇类激素具有营养、代谢及调节功能。有机体表面的脂类物质有防止机械损伤与防止热量散发等保护作用。脂类作为细胞的表面物质,与细胞识别,种特异性和组织免疫等有密切关系。 (二)脂肪的降解 在脂肪酶的作用下,脂肪水解成甘油和脂肪酸。甘油经磷酸化和脱氢反应,转变成磷酸二羟丙酮,纳入糖代谢途径。脂肪酸与ATP和CoA在脂酰CoA合成酶的作用下,生成脂酰CoA。脂酰CoA在线粒体内膜上肉毒碱:脂酰CoA转移酶系统的帮助下进入线粒体衬质,经β-氧化降解成乙酰CoA,在进入三羧酸循环彻底氧化。β-氧化过程包括脱氢、水合、再脱氢和硫解四个步骤,每次β-氧化循环生成FADH2、NADH、乙酰CoA和比原先少两个碳原子的脂酰CoA。此外,某些组织细胞中还存在α-氧化生成α羟脂肪酸或CO2和少一个碳原子的脂肪酸;经ω-氧化生成相应的二羧酸。 萌发的油料种子和某些微生物拥有乙醛酸循环途径。可利用脂肪酸β-氧化生成的乙酰CoA合成苹果酸,为糖异生和其它生物合成提供碳源。乙醛酸循环的两个关键酶是异柠檬酸裂解酶和苹果酸合成酶前者催化异柠檬酸裂解成琥珀酸和乙醛酸,后者催化乙醛酸与乙酰CoA生成苹果酸。 (三)脂肪的生物合成 脂肪的生物合成包括三个方面:饱和脂肪酸的从头合成,脂肪酸碳链的延长和不饱和脂肪酸的生成。脂肪酸从头合成的场所是细胞液,需要CO2和柠檬酸的参与,C2供体是糖代谢产生的乙酰CoA。反应有二个酶系参与,分别是乙酰CoA羧化酶系和脂肪酸合成酶系。首先,乙酰CoA在乙酰CoA羧化酶催化下生成,然后在脂肪酸合成酶系的催化下,以ACP 作酰基载体,乙酰CoA为C2受体,丙二酸单酰CoA为C2供体,经过缩合、还原、脱水、再还原几个反应步骤,先生成含4个碳原子的丁酰ACP,每次延伸循环消耗一分子丙二酸单酰CoA、两分子NADPH,直至生成软脂酰ACP。产物再活化成软脂酰CoA,参与脂肪合成或在微粒体系统或线粒体系统延长成C18、C20和少量碳链更长的脂肪酸。在真核细胞内,饱和脂肪酸在O2的参与和专一的去饱和酶系统催化下,进一步生成各种不饱和脂肪酸。高等动物不能合成亚油酸、亚麻酸、花生四烯酸,必须依赖食物供给。 3-磷酸甘油与两分子脂酰CoA在磷酸甘油转酰酶作用下生成磷脂酸,在经磷酸酶催化变成二酰甘油,最后经二酰甘油转酰酶催化生成脂肪。 (四)磷脂的生成 磷脂酸是最简单的磷脂,也是其他甘油磷脂的前体。磷脂酸与CTP反应生成CDP-二酰甘油,在分别与肌醇、丝氨酸、磷酸甘油反应,生成相应的磷脂。磷脂酸水解成二酰甘油,再与CDP-胆碱或CDP-乙醇胺反应,分别生成磷脂酰胆碱和磷脂酰乙醇胺。 二、习题 (一)名词解释 1.1.必需脂肪酸(essential fatty acid) 2.2.脂肪酸的α-氧化(α- oxidation) 3.3.脂肪酸的β-氧化(β- oxidation)
第七章脂类代谢习题及答案
第七章脂类代谢 一、知识要点 (一)脂肪的生物功能: 脂类是指一类在化学组成和结构上有很大差异,但都有一个共同特性,即不溶于水而易溶于乙醚、氯仿等非极性溶剂中的物质。通常脂类可按不同组成分为五类,即单纯脂、复合脂、萜类和类固醇及其衍生物、衍生脂类及结合脂类。 脂类物质具有重要的生物功能。脂肪是生物体的能量提供者。 脂肪也是组成生物体的重要成分,如磷脂是构成生物膜的重要组分,油脂是机体代谢所需燃料的贮存和运输形式。脂类物质也可为动物机体提供溶解于其中的必需脂肪酸和脂溶性维生素。某些萜类及类固醇类物质如维生素A、D、E、K、胆酸及固醇类激素具有营养、代谢及调节功能。有机体表面的脂类物质有防止机械损伤与防止热量散发等保护作用。脂类作为细胞的表面物质,与细胞识别,种特异性和组织免疫等有密切关系。 (二)脂肪的降解 在脂肪酶的作用下,脂肪水解成甘油和脂肪酸。甘油经磷酸化和脱氢反应,转变成磷酸二羟丙酮,纳入糖代谢途径。脂肪酸与ATP和CoA在脂酰CoA合成酶的作用下,生成脂酰CoA。脂酰CoA在线粒体内膜上肉毒碱:脂酰CoA转移酶系统的帮助下进入线粒体衬质,经β-氧化降解成乙酰CoA,在进入三羧酸循环彻底氧化。β-氧化过程包括脱氢、水合、再脱氢和硫解四个步骤,每次β-氧化循环生成FADH2、NADH、乙酰CoA和比原先少两个碳原子的脂酰CoA。此外,某些组织细胞中还存在α-氧化生成α羟脂肪酸或CO2和少一个碳原子的脂肪酸;经ω-氧化生成相应的二羧酸。 萌发的油料种子和某些微生物拥有乙醛酸循环途径。可利用脂肪酸β-氧化生成的乙酰CoA合成苹果酸,为糖异生和其它生物合成提供碳源。乙醛酸循环的两个关键酶是异柠檬酸裂解酶和苹果酸合成酶前者催化异柠檬酸裂解成琥珀酸和乙醛酸,后者催化乙醛酸与乙酰CoA生成苹果酸。 (三)脂肪的生物合成 脂肪的生物合成包括三个方面:饱和脂肪酸的从头合成,脂肪酸碳链的延长和不饱和脂肪酸的生成。脂肪酸从头合成的场所是细胞液,需要CO2和柠檬酸的参与,C2供体是糖代谢产生的乙酰CoA。反应有二个酶系参与,分别是乙酰CoA羧化酶系和脂肪酸合成酶系。首先,乙酰CoA在乙酰CoA羧化酶催化下生成,然后在脂肪酸合成酶系的催化下,以ACP作酰基载体,乙酰CoA为C2受体,丙二酸单酰CoA为C2供体,经过缩合、还原、脱水、再还原几个反应步骤,先生成含4个碳原子的丁酰ACP,每次延伸循环消耗一分子丙二酸单酰CoA、两分子NADPH,直至生成软脂酰ACP。产物再活化成软脂酰CoA,参与脂肪合成或在微粒体系统或线粒体系统延长成C18、C20和少量碳链更长的脂肪酸。在真核细胞内,饱和脂肪酸在O2的参与和专一的去饱和酶系统催化下,进一步生成各种不饱和脂肪酸。高等动物不能合成亚油酸、亚麻酸、花生四烯酸,必须依赖食物供给。 3-磷酸甘油与两分子脂酰CoA在磷酸甘油转酰酶作用下生成磷脂酸,在经磷酸酶催化变成二酰甘油,最后经二酰甘油转酰酶催化生成脂肪。 (四)磷脂的生成 磷脂酸是最简单的磷脂,也是其他甘油磷脂的前体。磷脂酸与CTP反应生成
第二十章 脂肪酸合成代谢-习题
第二十章脂肪酸合成代谢 一、选择题 ⒈关于不饱和脂肪酸合成,下列叙述错误的为() A、去饱和酶是一种混合功能的氧化酶; B、动物中去饱和酶要求细胞色素b5参加; C、植物中 去饱和酶要求黄素蛋白和铁硫蛋白参加;D、所有的生物都含有多不饱和脂肪酸;E、动物不能合成双键在△9以上的多烯脂酸。 ⒉关于硬脂酸生物合成() A、动物中的酰基载体是乙酰CoA; B、植物中酰基载体是ACPSH; C、动植物所需要的还原剂 均是NADPH+H+;D、植物硬脂酸合成地点在叶绿体中;E、动物合成地点在线粒体和内质网 ⒊下列有关脂肪酸从头合成的叙述哪个是正确的?() A、不直接利用乙酰CoA; B、仅合成少于10个碳原子的脂肪酸链; C、它需要丙二酸单酰CoA; D、发生在线粒体内; E、NAD+参与该过程 ⒋肝脏脂肪酸合成酶复合物的纯化剂和乙酰CoA、14C标记羧基的丙二酸单酰CoA、酰基载体蛋白 以及NADPH一起保温,分离合成的棕榈酸并测定14C的分布,预期是下列结果中的哪一种?() A、所有奇数碳原子被标记; B、除C1外所有的奇数碳原子被标记; C、所有的偶数碳原子被标 记;D、除C16外,所有的偶数碳原子被标记;E、没有一个碳原子被标记 ⒌从甘油和软脂酸生物合成一分子甘油三软脂酸酯,消耗多少高能磷酸键?() A、1; B、3; C、5; D、7; E、9 ⒍二脂酰甘油+NDP-胆碱→NMP+磷脂酰胆碱反应,NMP代表什么?() A、AMP; B、CMP; C、GMP; D、TMP; E、UMP ⒎脂肪酸β-氧化的逆反应可见于() A、胞浆中脂肪酸的合成; B、胞浆中胆固醇的合成; C、线粒体中脂肪酸的延长; D、内质网中 脂肪酸的延长;E、不饱和脂肪酸的合成 二、判断是非 ⒈真核生物脂肪酸合成酶系各成员共价串联成一条多肽链发挥作用。() ⒉硫脂酶是脂肪酸合成酶系中的重要成员。() ⒊β-羟脂酰ACP脱水酶催化下产生△2反丁烯酰ACP。() ⒋脂肪酸合成的直接前体是丙二酸单酰CoA。() ⒌在脂肪酸合成过程中,中间产物以非共价键与载体ACP相联。() ⒍从乙酰CoA合成1分子棕榈酸,必须消耗8分子A TP。() ⒎酰基载体蛋白(ACP)是饱和脂肪酸碳链延长途径中二碳单位的活化供体。() ⒏如果动物长期饥饿就要动用体内的脂肪,这时分解酮体的速度大于生成酮体的速度。() ⒐胆固醇与某些疾病如胆管阻塞、胆结石和动脉硬化等密切有关,如果能够一方面完全禁食胆固醇, 另一方面完全抑制胆固醇的生物合成,将有助于健康长寿。() 三、填空题 ⒈乙酰CoA和CO2生成,需要消耗高能磷酸键,并需要催化,该酶包括三 种成分,他们是、和;动物中乙酰CoA羧化酶受激活,并受抑制。 ⒉脂肪酸生物合成的限速酶为。