基因克隆的质粒载体

基因克隆的质粒载体
在大肠杆菌的各种菌体中找到了许多种不同类型的质粒,其中已经作了比较
详尽研究的主要有F质粒、R质粒和Col质粒。
①F质粒又叫F因子或性质粒(sex plasmid )。它们能够使寄主染色体上的基因和F质粒一道转移到原先不存在该质粒的受体细胞中去。
②R质粒通称抗药性因子。它们编码有一种或数种抗菌素抗性基因,并且通常能够将此种抗性转移到缺管该质粒的适宜的受体细胞,使后者也获得同样的抗菌素抗性能力。
③Col质粒即所谓产生大肠杆菌素因子。它们编码有控制大肠杆菌素合成的基因。大肠杆菌是一类可以使不带有Col质粒的亲缘关系密切的细菌菌株致死的蛋白质。
第一节质粒的一般生物学特性
、质粒DNA
细菌质粒是存在于细胞质中的一类独立于染色体的自主复制的遗传成份。绝大多数的质粒都是由环形双DNA组成的复制子(图4-1)。
大肠杼蕭质轉分子的跡构示愆图
质腔垦细宿聲色怖并简擁自風豐制的咔理取fit的井产?集码抗阳素抗性基囚的质垃叫尺质粒*兀部分质暫垦町以转擁的.但也存在着不陡墙转暮的成粒
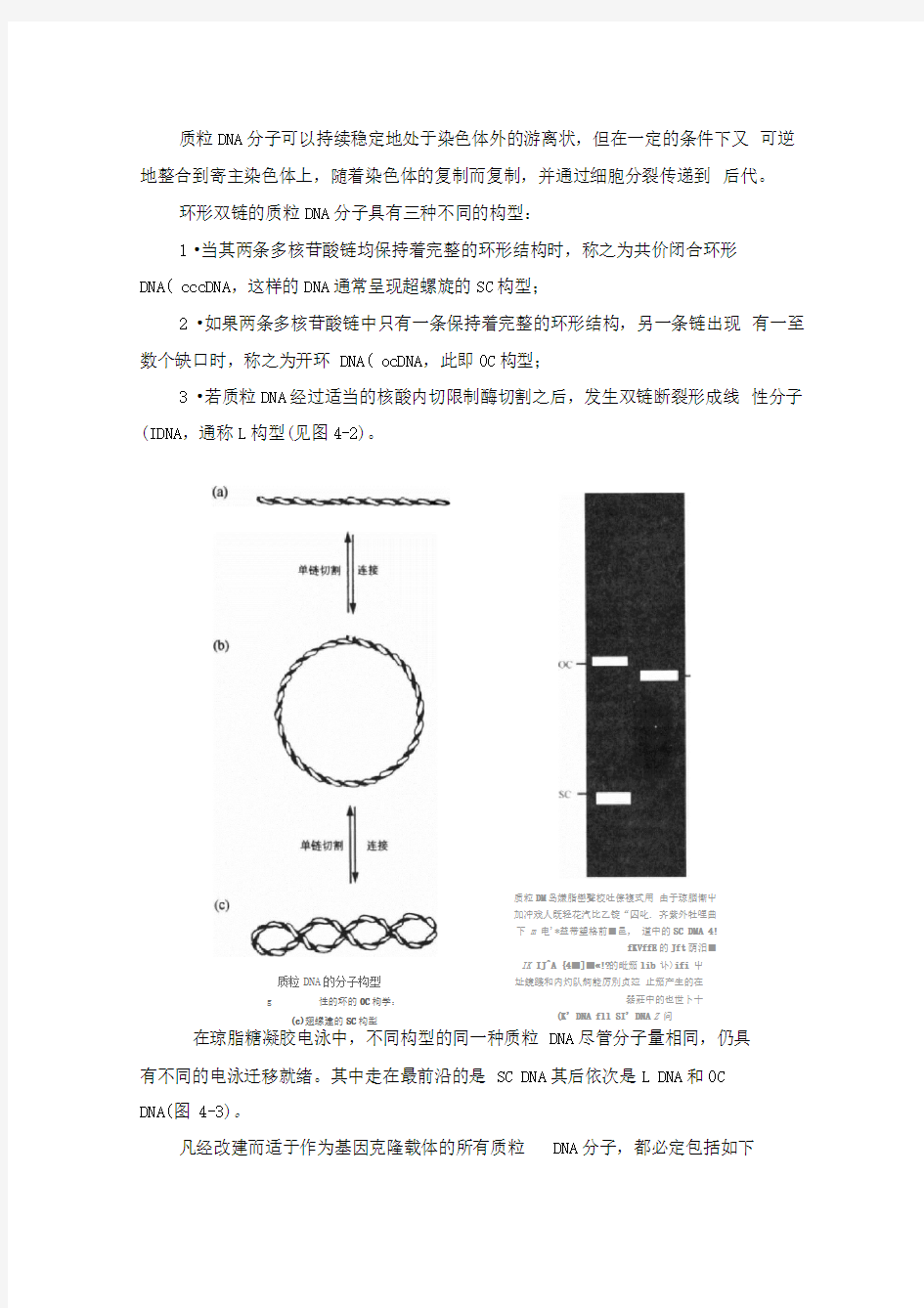
质粒DNA 分子可以持续稳定地处于染色体外的游离状,但在一定的条件下又 可逆地整合到寄主染色体上,随着染色体的复制而复制,并通过细胞分裂传递到 后代。
环形双链的质粒DNA 分子具有三种不同的构型:
1 ?当其两条多核苷酸链均保持着完整的环形结构时,称之为共价闭合环形
DNA( cccDNA ,这样的DNA 通常呈现超螺旋的SC 构型;
2 ?如果两条多核苷酸链中只有一条保持着完整的环形结构,另一条链出现 有一至数个缺口时,称之为开环 DNA( ocDNA ,此即0C 构型;
3 ?若质粒DNA 经过适当的核酸内切限制酶切割之后,发生双链断裂形成线 性分子(IDNA ,通称L 构型(见图4-2)。
在琼脂糖凝胶电泳中,不同构型的同一种质粒 DNA 尽管分子量相同,仍具
有不同的电泳迁移就绪。其中走在最前沿的是 SC DNA 其后依次是L DNA 和0C DNA(图 4-3)。
凡经改建而适于作为基因克隆载体的所有质粒 DNA 分子,都必定包括如下
质粒DNA 的分子构型
g 性的环的0C 枸学:
(c)翅螺建的SC 构型 质粒DM 岛嫌脂巒聲校吐僚複式用 由于琼胭慚屮
加冲戏人既轻花汽比乙锭“囚叱. 齐紫外牡哩曲
下 m 电'*益带望格前■邑, 道中的SC DMA 4!
fKVffE 的Jft 荫汨■
IK IJ^A {4■]■?!?的毗笳lib 讣)ifi 屮
址鏡踐和内灼队舸能厉別贞竝 止笳产生的在器莊中的也世卜十 (K' DNA fll SI' DNA Z 问
三种共同的组成部分,即复制基因(replicator )、选择性记和克隆位点。
二、质粒DNA编码的表型
质粒DNA仅占细胞染色体组的1%^3流右,但却编码着一些重要的非染色体控制的遗传性状。其中对抗菌素的抗性最质粒的最重要的编码特性之一。
三、质粒DNA的转移
(1)质粒的类型革兰阴性细菌的质粒可以分成接合型和非接合型的两种类群。
接合型的质粒(conjugative plasmid ),又叫自我转移的质粒。它们除了具有自主复制所必须的遗传信息之外,还带有一套控制细菌配对和质粒接合转移的基因。
非接合型的质粒(non-conjugative plasmid ),亦叫不能自我转移的质粒。它们虽然具有自主复制的遗传信息,但失去了控制细胞配骊和接合转移的基因,因此是不能够从一个细胞自我转移到另一个细胞。
(2)F质粒又叫F因子,即致育因子(fertility factor )的简称,是在
某些大肠杆菌细胞中发现的一种最有代表性的单拷贝的接合型质粒。F质粒有
三种不同的存在方式:
(i ) F+细胞:以染色体外环形双链质粒DNA形式存在,其上不带有任何来自寄主染色体的基因或DNA区段。
(ii ) F'细胞:以染色体外环形双链质粒DNA形式存在,同时在其上还携带着细菌的染色体基因或DN区段。
(iii ) Hfr细胞(高频重组细胞):以线性DNA形式从不同位点整合到寄主染色体。
F因子是雄性决定因子,所以F+细胞又叫雄性细胞,与此相应的F-细胞则叫做雌性细胞。F+细胞的表面可以形成一种叫做性须(pilus )的结构,它促进雄性细胞同雌性细胞进行配对。在合适的条件下,将雄性细胞和雌性细胞混合培养,由于性须的作用,就会形成雌-雄细胞配对。我们称这种过种为细菌的接合
作用(conjugation )。配对之后F-受体细胞获得了F因子,也变成为F+细胞由F因子整合到染色体而成的Hfr细胞,就可能相发寄主染色体发生高频转
移。这是一种可逆的过程,在一定的条件下,Hfr细胞又可重新变参展F+或F'
细胞。质粒的主要类型见表4-1。
(3)质粒DNA勺接合转移
①细胞交配对的形成雄性细胞的性须顶端与受体细胞表面接触之后,便会迅速收缩,把给体细胞与受体细胞拉在一起。因此,性须在确立配对细胞表面间的紧密接触方面,起着至关重要的作用。但是,大肠杆菌雄性细胞是不会同其它的亦带有F质粒的细胞发生配对作用的,因为traS和traT编码的“表面排斥” 蛋白质,使此种细胞无法成为接合作用的受体。这就决定了雄性细胞只能同不具
F因子的雌性细胞配对的特异性。
②质粒DNA的转移F质粒DNA勺转移是从转移起点oriT开始的。当细胞
交配对建立之后,TraY和TraI蛋白质首先在oriT位点作单链切割,随后缺口链在其游离的5'-端的引导下转移到受体细胞,并作为模板合成互补链,形成新的质粒分子。于是受体细胞便转变成为具有F因子的雄性细胞如图4-4。
质粒DNA 的犊合转移及复制
给体细胸中的单雄质粒DNA 经过孔追进入受体纸雷.井在给体和受怀细胞中甘别进Tf ONA 的合虛.
结杲实砚T 质粒DNR 的转移,井增加了质蛙的於贝!Uf 为筒明起見.圈中f 艮示出部分统?!星结构)
四、质粒DNA 的迁移作用
非接合型的质粒,由于分子小,不足以编码全部转移体系所需要的基因,
因 而不能够自多转移。但如果在其寄主细胞中存在着一种接合型的质粒, 那么它们 通常也是可以被转移的。这种由共存的接合型质粒引发的非接合型质粒的转移过 程,叫做质粒的迁移作用(mobilization )。
ColEI 是一种可以迁移但是属于非接合型的质粒。需要质粒自己编码的两种 基因参与。一个是位于 ColEI DNAh 的特异位点bom 另一个是ColEI 质粒特有 的弥散的基因产物,即 mob 基因(mobilization gene )编码的核酸酶。mob 基 因和bom 基因参与ColEI 质粒的迁移作用这个结论,是根据图4-5的实验结果作 出的。 DMA 斬铸
解雄"I
细胞外瞳
受怵细胞 DNASrS
转移樋
貝fiimYI 切割■
解罐醐的嗔孔”
RNA 弓I 枸
Po
基因工程原理讲义:基因克隆的质粒载体
第六讲基因克隆的质粒载体 中国科学院遗传与发育生物学研究所 2017年8月
基因克隆的质粒载体 一、导言 1.质粒是一类引人注目的亚细胞有机体 其结构比病毒还要简单,既无蛋白质外壳,也无细胞外生命周期,只能在寄主细胞内增殖,并随着寄主细胞的分裂而被遗传下去。2.质粒的类型多种多样 F质粒:F因子或性质粒(Sex plasmid),它能够使寄主染色体上的基因与F因子(F factor)一道转移到原先不存在该质粒的 寄主受体细胞中去。 R质粒:通称抗药性因子(Resistant factor, R factor),编码一种或数种抗菌素抗性基因,并能将此抗性转移到缺乏该质粒 的适宜的受体细胞中去。 Col质粒:所谓Col质粒,即是一种产生大肠杆菌素的因子,编码控制大肠杆菌素合成的基因。大肠杆菌素可使不带Col 质粒的亲缘关系密切的细菌菌株致死。 3.质粒载体 70年代在实验室构建的一类最普遍使用的基因克隆载体。
二、质粒的一般特性 1.质粒DNA(细菌质粒定义) *1.大肠杆菌的质粒是独立于寄主染色体以外的自主复制的共价、闭合、环形的双链DNA分子(covalently closed circular DNA, cccDNA)。除了酵母的杀伤质粒(Killer plasmid)是RNA质粒外,所有的质粒都是质粒DNA。但是质粒DNA的复制又必须依赖 于寄主提供核酸酶及蛋白质。 *2.质粒DNA分子大小 文献中有3种说法:小的仅有103KD,仅能编码2-3种蛋白质; 大的可达105KD,两者相差上百倍。 1Kb~200Kb (Sambrok et al.) 5Kb~400Kb (Lehninger) MD(megadaltons)=106D (兆道尔顿) 1.5Kb≈1MD *3.质粒DNA与寄主染色体DNA间的关系 一般情况下,质粒DNA可持续地处于寄主染色体外的游离状
基因克隆载体上的各种常用蛋白标签
基因克隆载体上的各种常用蛋白标签 蛋白标签(proteintag)是指利用DNA体外重组技术,与目的蛋白一起融合表达的一种多肽或者蛋白,以便于目的蛋白的表达、检测、示踪和纯化等。随着技术的不断发展,研究人员相继开发出了具有各种不同功能的蛋白标签。目前,这些蛋白标签已在基础研究和商业化产品生产等方面得到了广泛的应用。 美国GeneCopoeia(复能基因)为客户提供50多种蛋白标签,可以满足客户的不同需求,包括各种最新型的标签,如:SNAP-Tag?、Halo Tag?、AviTag?、Sumo等;也提供齐全的各种常用标签,如eGFP、His、Flag等等标签。 以下是部分蛋白标签的特性介绍,更加详细的介绍可在查询产品的结果列表里面看到各种推荐的蛋白标签和载体。 TrxHIS His6是指六个组氨酸残基组成的融合标签,可插入在目的蛋白的C末端或N末端。当某一个标签的使用,一是能构成表位利于纯化和检测;二是构成独特的结构特征(结合配体)利于纯化。组氨酸残基侧链与固态的镍有强烈的吸引力,可用于固定化金属螯合层析(IMAC),对重组蛋白进行分离纯化。使用His-tag有下面优点: 标签的量小,只有~0.84KD,而GST和蛋白A分别为~26KD和~30KD,一般不影响目标蛋白的功能; His标签融合蛋白可以在非离子型表面活性剂存在的条件下或变性条件下纯化,前者在纯化疏水性强的蛋白得到应用,后者在纯化包涵体蛋白时特别有用,用高浓度的变性剂溶解后通过金属螯和去除杂蛋白,使复性不受其它蛋白的干扰,或进行金属螯和亲和层析复性; His标签融合蛋白也被用于蛋白质-蛋白质、蛋白质-DNA相互作用研究; His标签免疫原性相对较低,可将纯化的蛋白直接注射动物进行免疫并制备抗体。 可应用于多种表达系统,纯化的条件温和; 可以和其它的亲和标签一起构建双亲和标签。 Flag标签蛋白 Flag标签蛋白为编码8个氨基酸的亲水性多肽(DYKDDDDK),同时载体中构建的Kozak序列使得带有FLAG的融合蛋白在真核表达系统中表达效率更高。FLAG作为标签蛋白,其融合表达目的蛋白后具有以下优点: FLAG作为融合表达标签,其通常不会与目的蛋白相互作用并且通常不会影响目的蛋白的功能、性质,这样就有利用研究人员对融合蛋白进行下游研究。 融合FLAG的目的蛋白,可以直接通过FLAG进行亲和层析,此层析为非变性纯化,可以纯化有活性的融合蛋白,并且纯化效率高。 FLAG作为标签蛋白,其可以被抗FLAG的抗体识别,这样就方便通过Western Blot、ELISA等方法对含有FLAG的融合蛋白进行检测、鉴定。
分子克隆技术第三章
2017/2/21 第三章载体 第一节基因克隆技术概述 一、基因克隆技术 基因克隆技术包括把来自不同生物的基因同有自主复制能力的载体DNA在体外人工连接,构建成新的重组的DNA,然后送入受体生物中去表达,从而产生遗传物质和状态的转移和重新组合。 二、目的基因的取得 1、直接 2、反转录酶 3、化学合成 4、基因文库 5、PCR ?首先利物理方法(如剪切力、超声波等)或酶化学方法(如限制性 内切核酸酶)将生物细胞染色体DNA切割成为基因水平的许多片段,继而将这些片段与适当的载体结合。将重组DNA转入受体菌扩增,获得无性繁殖的基因文库,再结合筛选方法,从众多的转化子菌株中选出含有某一基因的菌株,从中将重组的DNA分离、回收。这种方法也就是应用基因工程技术术分离目的基因,其特点是绕过直接分离基因的难关,在基因组DNA文库中筛选出目的基因。可以说这是利用“溜散弹射击”原理去“命中”某个基因。由于目的基因在整个基因组太小,在像当程度上还得靠“碰运气”,所以人们称这个方法为“鸟 枪法”或“散弹枪”实验法。 三、重组体的构建 1、载体 要把一个有用的基因通过基因工程手段送进生物细胞中,需要运载工具,携带外源基因进入受体细胞的这种工具叫载体(Vector)。 (1)质粒(plasmid) (2)噬菌体λ的衍生物 (3)科斯质粒(cosmid) (4)单链DNA噬菌体 M13(5)病毒?面包酵母吲哚甘油磷酸脱氢酶基因的制取,先 用Eco RI把面包酵母DNA切成许多片段,使这些片段与λ载体连成重组DNA,可把这些重组DNA导入“吲哚甘油磷酸脱氢酶型组氨酸缺陷型”大肠杆菌,在基本培养基中培养。只有引入了该基因的细菌才能生长。进一步分离这种菌株,可以得到目的基因。 2、载体的性质 1)它必须具有能够在某些宿主细胞中独立地自我复制和表达的能力。 2)载体DNA的分子量应该较小。 3)载体上最好应具有两个以上的容易检测的遗传标记(如抗药性基因等),以赋予宿主细胞以不同的表型。 4)载体应该具有多个限制性内切酶的单一切点;载体上的单一酶切位点最好是位于检测表型的遗传标记基因之内,这样目的基因是否已连接载体就可以通过这一表型的改变与否而得知,利于筛选重组体。 3、酶系的选用
克隆载体与表达载体教程文件
克隆载体:大多是高拷贝的载体,一般是原核细菌,将需要克隆的基因与克隆载体的质粒相连接,再导入原核细菌内,质粒会在原核细菌内大量复制,形成大量的基因克隆,被克隆的基因不一定会表达,但一定被大量复制。克隆载体只是为了保存基因片段,这样细胞内不会有很多表达的蛋白质而影响别的工作。 克隆载体(Cloning vector ):携带插入外源片段的质粒或噬菌体,从而产生更多物质或蛋白质产物。(这是为“携带”感兴趣的外源DNA、实现外源DNA的无性繁殖或表达有意义的蛋白质所采用的一些DNA 分子。) 其中,为使插入的外源DNA序列可转录、进而翻译成多肽链而设计的克隆载体又称表达载体。 是否含有表达系统元件,即启动子--核糖体结合位点--克隆位点--转录终止信号,这是用来区别克隆载体和表达载体的标志。 表达载体:有的是高拷贝的,有的是低拷贝的,各有各的用处,是一些用于工程生产的细菌,被导入的目标基因会在此类细菌中得到表达,生产出我们需要的产物,导入的基因是由克隆载体产出的。表达载体具有较高的蛋白质表达效率,一般因为具有强的启动子。 表达载体(Expression vectors)就是在克隆载体基本骨架的基础上增加表达元件(如启动子、RBS、终止子等),是目的基因能够表达的载体。如表达载体pKK223-3是一个具有典型表达结构的大肠杆菌表达载体。其基本骨架为来自pBR322和pUC的质粒复制起点和氨苄青霉素抗性基因。在表达元件中,有一个杂合tac强启动子和终止子,在启动子下游有RBS位点(如果利用这个位点,要求与ATG之间间隔5-13bp),其后的多克隆位点可装载要表达的目标基因。 (RBS位点:1974年Shine和Dalgarno首先发现,原核生物,在mRNA上有核糖体的结合位点,它们是起始密码子AUG和一段位于AUG上游3~10 bp处的由3—9bp组成的序列。这段序列富含嘌呤核苷酸,刚好与16S rRNA 3,末端的富含嘧啶的序列互补,是核糖体RNA的识别与结合位点。根据发现者的名字,命名为Shine-Dalgarno序列,简称S-D序列。 由于它正好与30S小亚基中的16s rRNA3’端一部分序列互补,因此S-D序列也叫做核糖体结合序列。 真核生物存在于真核生物mRNA的一段序列,其在翻译的起始中有重要作用。加Kozark sequence(GCCACC), Kozak sequence是用来增强真核基因的翻译效率的。是最优化的ATG环境,避免ribosome出现leaky scan) 克隆载体目的在于复制足够多的目标质粒,所以常带有较强的自我复制元件,如复制起始位点等,往往在菌体内存在多拷贝,所以抽质粒会抽出一大堆。但不具备表达元件。而表达质粒有复杂的构成,为的是控制目标蛋白的表达,如各种启动子(T7),调节子(LacZ)等,而且以pET为代表的表达载体在菌体内都是低拷贝的,防止渗漏表达。 克隆载体只是把你要的基因片段拿到就可以了,不管读码框什么的,但是表达载体是不但要你的目的基因连在上面,而且要表达蛋白,所以就要求你的读码框不能乱了,否则就不能得到你想到的表达产物。 1.载体即要把一个有用的基因(目的基因——研究或应用基因)通过基因工程手段送到生物细胞(受体细胞),需要运载工具(交通工具)携带外源基因进入受体细胞,这种运载工具就叫做载体(vector)。 2. 载体的分类 按功能分成:(1)克隆载体: 都有一个松弛的复制子,能带动外源基因,在宿主细胞中复制扩增。它是用来克隆和扩增DNA片段(基因)的载体。(2)表达载体:具有克隆载体的基本元件(ori,Ampr,Mcs等)还具有转录/翻译所必需的DNA顺序的载体。 按进入受体细胞类型分:(1)原核载体(2)真核载体(3)穿梭载体(sbuttle vector)指在两种宿主生物体内复制的载体分子,因而可以运载目的基因(穿梭往返两种生物之间). 克隆载体顾名思义就是质粒拷贝数比较高,在做上游克隆时比较方便, 其重点在于质粒的复制.
基因克隆的质粒载体
基因克隆的质粒载体 在大肠杆菌的各种菌体中找到了许多种不同类型的质粒,其中已经作了比较详尽研究的主要有F质粒、R质粒和Col质粒。 ①F质粒又叫F因子或性质粒(sex plasmid)。它们能够使寄主染色体上的基因和F质粒一道转移到原先不存在该质粒的受体细胞中去。 ②R质粒通称抗药性因子。它们编码有一种或数种抗菌素抗性基因,并且通常能够将此种抗性转移到缺管该质粒的适宜的受体细胞,使后者也获得同样的抗菌素抗性能力。 ③Col质粒即所谓产生大肠杆菌素因子。它们编码有控制大肠杆菌素合成的基因。大肠杆菌是一类可以使不带有Col质粒的亲缘关系密切的细菌菌株致死的蛋白质。 第一节质粒的一般生物学特性一.质粒DNA 细菌质粒是存在于细胞质中的一类独立于染色体的自主复制的遗传成份。绝大多数的质粒都是由环形双DNA组成的复制子(图4-1)。 质粒DNA分子可以持续稳定地处于染色体外的游离状,但在一定的条件下又可逆地整合到寄主染色体上,随着染色体的复制而复制,并通过细胞分裂传递到后代。 环形双链的质粒DNA分子具有三种不同的构型: 1.当其两条多核苷酸链均保持着完整的环形结构时,称之为共价闭合环形DNA(cccDNA),这样的DNA通常呈现超螺旋的SC构型; 2.如果两条多核苷酸链中只有一条保持着完整的环形结构,另一条链出现有一至数个缺口时,称之为开环DNA(ocDNA),此即OC构型; 3.若质粒DNA经过适当的核酸内切限制酶切割之后,发生双链断裂形成线性分子(IDNA),通称L构型(见图4-2)。 在琼脂糖凝胶电泳中,不同构型的同一种质粒DNA,尽管分子量相同,仍具有不同的电泳迁移就绪。其中走在最前沿的是SC DNA,其后依次是L DNA和OC DNA(图4-3)。 凡经改建而适于作为基因克隆载体的所有质粒DNA分子,都必定包括如下三种共同的组成部分,即复制基因(replicator)、选择性记和克隆位点。 二.质粒DNA编码的表型 质粒DNA仅占细胞染色体组的1%~3%左右,但却编码着一些重要的非染色体控制的遗传性状。其中对抗菌素的抗性最质粒的最重要的编码特性之一。
目的基因片段与克隆载体质粒的连接操作步骤
连接反应总体积为10μL,体系组成如下: 回收纯化的PCR扩增目的基因片段 7.0μL 10×Ligation buffer 1.0μL T载体(10ng/μL) 1.0μL T4 DNA ligase 1.0μL 共10.0ul 混均后,4℃连接18-24小时,连接所得克隆载体命名为TA-VP4-STI。 转化操作方法 1)取一管-80℃保存的感受态细胞,置冰上融化; 一次转化感受态细胞的建议用量为50-100ul,应注意所用DNA体积不要超过感受态细胞悬液体积的十分之一。以100ul为例: 2)加入连接物(50ul的感受态细胞能够被1ng超螺旋质粒DNA所饱和),轻轻旋转离心管以混匀内容物,冰浴30min;
3)将离心管置于42℃热击60-90秒,然后迅速置冰浴2-3分钟,该过程不要摇动离心管; 4)向每个离心管中加入500ul液体LB培养基(不含抗生素),混匀后置于37℃摇床振荡培养45分钟(150转/分钟);目的是使质粒上相关的抗性标记基因表达,使菌体复苏。 5)将离心管内容物混匀,吸取100ul已转化的感受态细胞加到含相应抗生素的LB固体琼脂培养基上(含50 ug/ml氨苄青霉素),用无菌的弯头玻棒轻轻将细胞均匀涂开。将平板置于室温直至液体被吸收,倒置平板,37℃培养12-16小时。至红、白斑区分明显为止。 涂布用量可根据具体试验来调整:如转化的DNA总量较多,可取更少量转化产物涂布平板;反之,如转化的DNA总量较少,可取200-300ul 转化产物涂布平板。如果预计的克隆较少,可通过离心 (4000rpm,2min)后析除部分培养液,悬浮菌体后将其涂布于一个平板中。(涂布剩余的菌液可置于4℃保存,如果次日的转化菌落数过少可以将剩下的菌液再涂布新的培养板)
基因的克隆、表达载体构建及功能验证Word版
基因的克隆、表达载体构建及功能验证(一般性方法) 一、基因克隆 ★事前三问 a.克隆这个基因干什么?它有什么功能? b.这个基因在哪种材料中扩增? c.材料需要怎么处理? ◎ 实验前准备工作 a.设计引物,准备材料, b.购置试剂:Taq酶、反转录试剂盒、凝胶回收试剂盒、质粒提取试剂盒、连接试 剂盒 c.实验试剂及用具:枪头、离心管、培养皿、滤纸灭菌;Amp+ 、Kan+等抗生素准备 ※ 基本流程 提取和纯化RNA—cDNA第一条链合成—PCR—凝胶电泳—胶回收—连接—转化— 涂平板—挑单菌落—摇菌—提质粒—测序 1.总RNA的提取、纯化及cDNA第一链合成 1.1叶片、根总RNA的提取 Trizol是一种高效的总RNA抽提试剂,内含异硫氰酸胍等物质,能迅速裂解植物细胞,抑制细胞释放出的核酸酶,所提取的RNA完整性好且纯度高,以利于下一步的实验。 1)实验前准备 预先配制0.1%的DEPC水(ddH2O中含0.1%DEPC,V/V,37 ℃过夜处理12 h),高温灭菌后,用DEPC水配制 75%乙醇,研钵、量筒、试剂瓶等需200℃灭菌至少4 h,所用枪头和枪盒均去RNA酶处理(直接购买)。 2)Trizol 法(小麦)叶片或根的总RNA实验步骤如下: (1)提前在1.5 ml离心管中加入1 mlTrizol,然后将200 mg样品液氮中研磨成白色粉末,移入管内,用力摇15 s,在15-30℃温育5 min,使核酸蛋白复合物完全分离。 (2)4℃,12000g离心10min,取上清,离心得到的沉淀中包括细胞外膜、多糖、高分子量DNA,上清中含有RNA。 (3)吸取上清液加0.2 ml氯仿,盖好盖,用力摇15 s,15~30 ℃温育2~3 min。(4)在≤12000g,4℃离心10 min,样品分为三层:底层为黄色有机相,上层为无色水相和一个中间层,RNA主要在水相中,水相体积约为所用TRIzol试剂的60%。(5)将上层水相转移到新的1.5 ml离心管中,加2倍体积的无水乙醇沉淀RNA,室温静止30 min。 (6)在≤12000g,4℃离心10 min,离心前看不出RNA沉淀,离心后在管侧和管底出现胶状沉淀。
质粒载体
载体主要有病毒和非病毒两大类,其中质粒DNA是一种新的非病毒转基因载体。 一、一个合格质粒的组成要素 a复制起始位点Ori 即控制复制起始的位点。原核生物DNA分子中只有一个复制起始点。而真核生物DNA分子有多个复制起始位点。 b 抗生素抗性基因可以便于加以检测,如Amp+ ,Kan+ c 多克隆位点MCS 克隆携带外源基因片段 d P/E 启动子/增强子 e Terms 终止信号 f 加poly(A)信号可以起到稳定mRNA作用 二、如何阅读质粒图谱 第一步:首先看Ori的位置,了解质粒的类型(原核/真核/穿梭质粒) 第二步:再看筛选标记,如抗性,决定使用什么筛选标记。 (1)Ampr 水解β-内酰胺环,解除氨苄的毒性。 (2)tetr 可以阻止四环素进入细胞。 (3)camr 生成氯霉素羟乙酰基衍生物,使之失去毒性。 (4)neor(kanr)氨基糖苷磷酸转移酶使G418(长那霉素衍生物)失活 (5)hygr 使潮霉素β失活。 第三步:看多克隆位点(MCS)。它具有多个限制酶的单一切点。便于外源基因的插入。如果在这些位点外有外源基因的插入,会导致某种标志基因的失活,而便于筛选。决定能不能放目的基因以及如何放置目的基因。 第四步:再看外源DNA插入片段大小。质粒一般只能容纳小于10Kb的外源DNA片段。一般来说,外源DNA片段越长,越难插入,越不稳定,转化效率越低。 第五步:是否含有表达系统元件,即启动子-核糖体结合位点-克隆位点-转录终止信号。这是用来区别克隆载体与表达载体。克隆载体中加入一些与表达调控有关的元件即成为表达载体。选用那种载体,还是要以实验目的为准绳。 启动子-核糖体结合位点-克隆位点-转录终止信号 a 启动子-促进DNA转录的DNA顺序,这个DNA区域常在基因或操纵子编码顺序的上游,是DNA分子上可以与RNApol特异性结合并使之开始转录的部位,但启动子本身不被转录。b增强子/沉默子-为真核基因组(包括真核病毒基因组)中的一种具有增强邻近基因转录过程的调控顺序。其作用与增强子所在的位置或方向无关。即在所调控基因上游或下游均可发