海马结构及图
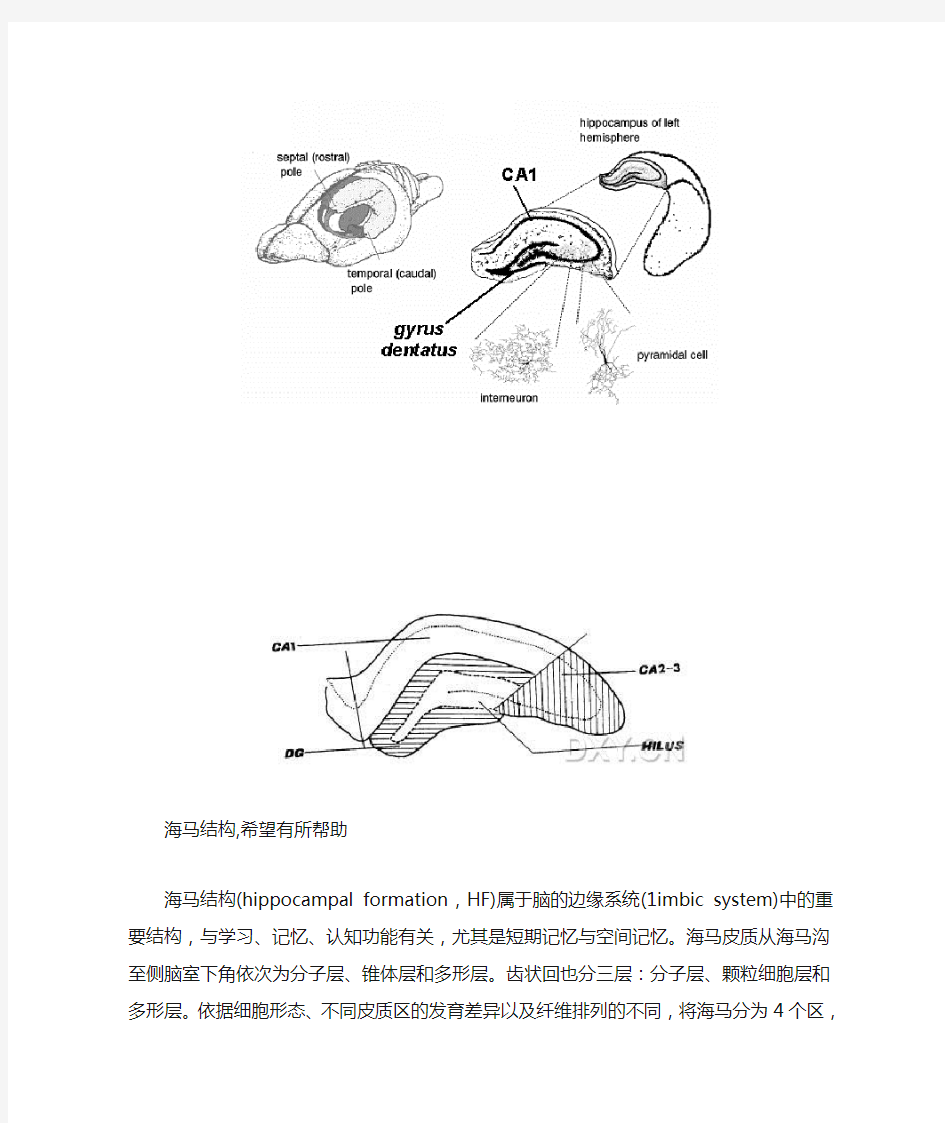

海马结构,希望有所帮助
海马结构(hippocampal formation,HF)属于脑的边缘系统(1imbic system)中的重要结构,与学习、记忆、认知功能有关,尤其是短期记忆与空间记忆。海马皮质从海马沟至侧脑室下角依次为分子层、锥体层和多形层。齿状回也分三层:分子层、颗粒细胞层和多形层。依据细胞形态、不同皮质区的发育差异以及纤维排列的不同,将海马分为4个区,即CAl、CA2、CA3、CA4区。海马结构是大脑边缘系统的重要组成部分.在进化上是大脑的古皮质,位于大脑内侧面颞叶的内侧深部,左右对称。一般认为海马结构由海马或称Ammon角、齿状回、下托及海马伞组成,结构比较复杂。在功能和纤维联系上,不仅与嗅觉有关,更与内脏活动.情绪反应和性活动有密切关系。细胞学研究表明,海马头部主要是由CAI区折叠而成,而CAI区对缺氧等损伤最为敏感,也被称为易损区,因此海马头部也是最易发生病变的部位。
海马结构由海马(hippoeampus)、齿状回(dentate gyrls)、下托(subiculum)和围绕胼胝体的海马残体(hippoeampal rudimerit)组成,其中海马为体积最大最主要的部分。
大脑海马(hippocampus)是位于脑颞叶内的一个部位的名称,人有两个海马,分别位于左右脑半球. 它是组成大脑边缘系统的一部分,担当着关于记忆以及空间定位的作用. 名字来源于这个部位的弯曲形状貌似海马(希腊语hippocampus).
在阿兹海默病中,海马是首先受到损伤的区域; 表现症状为记忆力衰退以及方向知觉的丧失。大脑缺氧(缺氧症)以及脑炎等也可导致海马损伤 .
在动物解剖中, 海马属于脑的演化过程中最古老的一部分。来源于旧皮质的海马在灵长类以及海洋生物中的鲸类中尤为明显。虽然如此, 与进化树上相对年轻的大脑皮层相比灵长类动物尤其是
人类的海马在端脑中只占很小的比例。相对新皮质的发展海马的增长在灵长类动物中的重要作用是使得其脑容量显着增长。
解剖
尽管关于海马与其向邻近的大脑皮层的表述尚缺乏一致的观点,通常情况下术语上的海马结构指的是齿状回, CA1-CA3部位(或CA4,常称为hilus区并被认为是齿状回的一部分), 以及脑下脚(另见阿蒙神之角)。CA1与CA3 部位构成严格意义上的海马。信息进入海马时由齿状回流入CA3到CA1再到脑下脚,在每个区域输入附加信息在最后的两个区域输出。CA2只占海马的一个很小部分通常将其对海马的功能忽略, 值得注意的一点是这一小区域似乎能抵抗由于例如癫痫等造成的大规模的细胞破坏。穿缘通路(perforant path)主要从内鼻叶(entorhinal cortex)获取信息, 通常被认为是海马输入信息的主要来源。一般记忆中的作用
心理学家与神经学家对海马的作用存在争论,但是都普遍认同海马的重要作用是将经历的事件形成新的记忆(情景记忆或自传性记忆). 一些研究学者认为应该将海马看作对一般的陈述性记忆起作用内侧颞叶记忆系统的一部分。(陈述性记忆指的是那些可以被明确的描述的记忆,如“昨天晚饭吃了什么”这样的关于经历过的事情的情景记忆,以及“北京是中国的首都”这样的关于知识的概念记忆).
有迹象显示,虽然这些形式的记忆通常能终身持续,在一系列的记忆强化以后海马便中止对记忆的保持。海马的损伤通常造成难以组织新的记忆(顺行性失忆症), 而且造成难以搜索过去的记忆(逆行性失忆症)。尽管这样的逆行性效果通常在脑损伤的很多年之前就开始扩展,一些情况下相对久远一些的记忆能够维持下来。这表明海马将巩固以后的记忆转入了脑的其他的部位。但是,旧的记忆是如何储存的要用实验来检测的话存在一些难点。另外,在一些逆行性失忆症案例中,在海马遭受损伤的数十年前的记忆也受到了影响,导致了这一关于旧的记忆的观点的争议。
海马的损伤不会影响某一些记忆,例如学习新的技能的能力(如学习一种乐器),将设这样的能力依靠的是另外一种记忆(程序记忆)和不同的脑区域。有迹象表明着名的病人HM(作为治疗癫痫病的手段他的内侧颞叶被切除) 有组织新的概念记忆的能力
研究史
解剖学家Giulio Cesare Aranzi (约1564年)首先使用海马(hippocampus)一词形容这一大脑器官,源于此部位貌似海马。这一部位最初被认为司控嗅觉,而非现在周知的记忆储存作用。俄国学者Vladimir Bekhterev于1900年左右基于对一位有严重记忆紊乱的病患者的长期观察,首先提出海马与记忆相关。但是,其后的很长时间,学界习惯上关于海马的作用都被认为和其他大脑边缘系统一样,司控情绪。着名的病人HM的病例引起了众多科学家的关注,并使人开始认识到海马对记忆起重要作用。为减轻HM时常发作的癫痫症状其脑内侧颞叶被切除(其中就包括两个海马)由此导致了一系列的相关空间以及时间的记忆损伤。重要的是HM仍然能完成程序性任务的学习(这一点与纹状体相关联)甚至有着高于常人水平的智商. HM的智能与陈述性记忆展现出显着的分裂。绝大多数的哺乳类动物海马的大小取决于脑容量的大小,但是鲸类这一部位的发育相对不完全。
在医学上,「海马体」是大脑皮质的一个内褶区,在「侧脑室」底部绕「脉络膜裂」形成一弓形隆起,它由两个扇形部分所组成,有时将两者合称海马结构;海马体的机能是主管人类的近期主要记忆,有点像是计算机的内存,将几周内或几个月内的记忆鲜明暂留,以便快速存取。而失忆症病患的海马体中并没有任何近期记忆暂留。由这项实验可以初步证实人类的梦境并非是由海马体中的近期记忆抽取并组织而成
大脑地解剖结构和功能——布鲁德曼分区
大脑的解剖结构和功能——布罗德曼分区系统 布罗德曼分区是一个根据细胞结构将大脑皮层划分为一系列解剖区域的系统。神经解剖学中所谓细胞结构(Cytoarchitecture),是指在染色的脑组织中观察到的神经元的组织方式。 布罗德曼分区1909年由德国神经科医生科比尼安·布洛德曼(Korbinian Brodmann)提出。根据皮质细胞的类型及纤维的疏密把大脑皮质分为52个区,并用数字给予表示。Brodmann Area 1, BA1 Brodmann Area 2, BA2 Brodmann Area 3, BA3 位置:位于中央后回 (postcentral gyrus) 和前顶叶区。 功能:分别为体感皮层内侧、末尾和前端区,BA1、BA2、BA3共同组成体感皮层; 具备基本体感功能(first somatic sensory area)接受对侧肢体的感觉传入。Brodmann Area 4, BA4 位置:位于中央前回(precentral gyrus),中央沟(central sulcus)的内侧面 功能:初级运动皮层(first somatic motor area),包含“运动小人”(motor homunculus )。 控制行为运动,与BA6 (前)和BA3 、BA2 、BA1、(后)相连,同时与丘脑腹外侧核相连。 体感小人(Somatosensory Homunculus ) 传入体感信息较多的身体区域获得的皮层代表区域较大。比如手部在初级体感皮层中的代表区域比背部的大。体感皮质定位可用“体感小人”(Somatosensory homunculus)来表示。 Brodmann Area 5, BA5 位置:位于顶叶前梨状皮质区(梨状皮质piriform cortex为下边缘皮质的组成部分)。功能:与BA7形成体感联合皮层。 Brodmann Area 7, BA7 位置:位于顶叶皮质顶部,体感皮层后方,视觉皮层(visual area)上方。 功能:将视觉和运动信息联合起来;与BA5形成体感联合皮层;视觉-运动协调功能。 Sensory Areas---------Somatosensory Association Area 位置:位于初级躯体感觉皮层后方(BA5、BA7)
海马结构及图
海马结构,希望有所帮助 海马结构(hippocampal formation,HF)属于脑的边缘系统(1imbic system)中的重要结构,与学习、记忆、认知功能有关,尤其是短期记忆与空间记忆。海马皮质从海马沟至侧脑室下角依次为分子层、锥体层和多形层。齿状回也分三层:分子层、颗粒细胞层和多形层。依据细胞形态、不同皮质区的发育差异以及纤维排列的不同,将海马分为4个区,即CAl、CA2、CA3、CA4区。海马结构是大脑边缘系统的重要组成部分.在进化上是大脑的古皮质,位于大脑内侧面颞叶的内侧深部,左右对称。一般认为海马结构由海马或称Ammon角、齿状回、下托及海马伞组成,结构比较复杂。在功能和纤维联系上,不仅与嗅觉有关,更与内脏活动.情绪反应和性活动有密切关系。细胞学研究表明,海马头部主要是由CAI区折叠而成,而CAI区对缺氧等损伤最为敏感,也被称为易损区,因此海马头部也是最易发生病变的部位。 海马结构由海马(hippoeampus)、齿状回(dentate gyrls)、下托(subiculum)和围绕胼胝体的海马残体(hippoeampal rudimerit)组成,其中海马为体积最大最主要的部分。 大脑海马(hippocampus)是位于脑颞叶内的一个部位的名称,人有两个海马,分别位于左右脑半球. 它是组成大脑边缘系统的一部分,担当着关于记忆以及空间定位的作用. 名字来源于这个部位的弯曲形状貌似海马(希腊语hippocampus). 在阿兹海默病中,海马是首先受到损伤的区域; 表现症状为记忆力衰退以及方向知觉的丧失。大脑缺氧(缺氧症)以及脑炎等也可导致海马损伤 . 在动物解剖中, 海马属于脑的演化过程中最古老的一部分。来源于旧皮质的海马在灵长类以及海洋生物中的鲸类中尤为明显。虽然如此, 与进化树上相对年轻的大脑皮层相比灵长类动物尤其是
海马结构、扣带回
海马结构 一.形态 海马结构(hippocampal formation)包括海马(hippocampus)又称安蒙角(Ammon’s born),齿状回(dentate gyrus)和围绕胼胝体形成一圈的海马残件(灰被indusium grisem).齿状回随海马伞向后,至胼胝体压部,它与海马伞分开,改为束状回,束状回向前上与覆盖在胼胝体上面的灰质称胼胝体上回(ssupracallosal gyrus)(灰被)相连续,灰被中埋有一对纵纹,分别为内侧纵纹和外侧纵纹。灰被与纵纹就是海马及其白质的残件,它们向前经胼胝体膝与胼胝体下回连续。 (一)海马 海马形似中药海马,故得名。其位于侧脑室下角的底和内侧壁,全长约5cm,前段较膨大,称海马角,他被2-3个浅沟分开,沟间隆起,称海马趾;海马表面被室管膜上皮覆盖,室管膜上皮下面一层有髓鞘纤维称室床,室床纤维沿海马背内侧缘集中,形成白色扁带称海马伞,构成穹窿系统的起始步,它自海马趾伸向压部,续于穹窿角。 海马内的细胞构筑分为三层,从海马裂到脑室依次为①分子层;②椎体细胞层;③多形层。根据细胞形态和皮质区发育差异等特点,在横断面上海马又可分为CA1、CA2、CA3和CA4四个区。CA4位于齿状回门内,内有大的椎体细胞;CA3有来自齿状回颗粒细胞的轴突(即苔状纤维);CA2内有少量轴突;CA1内含有小的椎体细胞。 (二)齿状回 齿状回是一条灰皮质,由于血管进入形成沟而成齿状,故名。它位于海马的内侧,海马裂与海马伞之间,齿状回向后与束装回相连,其前端抵海马回钩和海马回之间。 海马接受扣带回来的纤维经扣带直接或间接地终止于海马,从隔核发出的纤维经穹窿,海马伞终止于海马CA3、CA4区和齿状回。一侧的海马也可经同侧海马伞,穹窿脚,通过海马联合投射至对侧的海马和齿状回,海马还可经室床通路接受内嗅区外侧份的传出纤维,这些纤维主要分布于CA1区和下托的深层,内侧份纤维则经穿通道、下托进入海马CA1-CA3斜角带核,在穹窿的行程中发出纤维至丘脑前核和板内核的吻部,部分纤维可向尾侧进入中脑被盖和中央灰质。 二.海马的功能 海马具有多方面的生理功能,50年代不少实验已经证明海马可接受来自外周的视觉、听觉、触觉、痛觉、本体感受性和内感受性刺激,感受性冲动经过脑干网状结构传递至海马,引起海马电活动的变化。近年来,由于神经科学的迅速发展,不少学者认为海马是情感和学习记忆等高级神经活动的重要部位。 (一)行为反应 毁损海马后,动物的行为发生一系列变化,最重要的是更乐于从事新的活动,在恐惧或应激的情况下,动物好动反应灵活,热衷于进行新的活动,有时也会出现“幻觉”,但很少出现攻击性动作。这些反应对动物具有一定的生理意义,如毁损海马的鼠,当遇到猫时,其表现为饥饿情绪反应增强,食欲亢进,性活动异
海马结构
海马结构 2010-06-18 10:19:05| 分类:专业相关| 标签:|字号大中小订阅 概述 海马结构(hippocampal formation)包括海马(又称安蒙角cornu AmmonisCA)、下托、齿状回和围绕胼胝体形成一圈的海马残件。齿状回至胼胝体压部,消失齿状外形,改称束状回,束状回向前上与覆盖胼胝体上面的深层灰质称灰被(又称胼胝体上回)相连续。灰被中埋有一对纵纹,分别为内侧纵纹与外侧纵纹。灰被与纵纹就是海马及其白质的残件。它们向前经胼胝体膝与终板旁回连续。 位置与外型 海马(hippocampus)形如中药海马故名。位于侧脑室下角底兼内侧壁,全长5 cm。海马前端较膨大称海
马足,它被2-3个浅沟分开,沟间隆起称海马趾。海马是一条镰状隆嵴,自胼胝体压部向前到侧脑室的颞端。海马至胼胝体压部时,从齿状回和海马旁回间翻出称Retzius回。 海马结构的位置 海马表面被室管膜上皮覆盖。室管膜上皮下面有一层有髓纤维称为海马槽(又称室床alveus)。室床纤维沿海马背内侧缘集中,形成白色扁带称海马伞(fimbria of hippocampus),它自海马趾伸向压部,续于穹隆脚(crus of fomix)。海马伞的游离缘直接延续于其上方的脉络丛,两者间隔以脉络裂。
海马结在下角的发育 齿状回(dentate gyms)是一狭条皮质;由于血管进入被压成许多横沟呈齿状,故名。它位于海马的内侧,介于海马沟与海马伞之间。齿状回向前伸展至钩的切迹,在此急转弯,成光滑小束横过钩的下面,这横行段称齿状回尾。齿状回尾将钩分成前部的前钩回,后部的边叶内回。齿状回向后与束状回(fasciolar gyrus) 相连。 在海马结构发育较好的颞中平面,作一个大脑半球的冠状切面,海马结构呈双重“C”形环抱的外形,大C锁住小C。大C代表海马,它开口向腹内侧。小C代表齿状回,位于海马沟的背内侧,开口朝向背侧。海马 沟的腹侧为下托(subiculum)。 海马结构的位置与安排,从发育过程来理解比较清楚。 在胚胎3个月,两个半球内侧壁上各显出一条纵行加厚部分称海马嵴(hippocampal ridge),这是海马结构的原基。此嵴的上方为海马沟。此嵴以下,内侧壁很薄弱,而有血管分支伸入形成一纵褶称脉络襞,它突入侧脑室,形成侧脑室脉络丛,突入处称脉络裂(choroid fissure)。伴随着大脑皮质的扩展,因胼胝体纤维的急剧发展,以致海马结构的各部发展不均匀。背侧部分很少分化。在成人它形成一个残余的薄层灰质称灰被(indusium griseum)覆盖在胼胝体上方。海马结构的腹侧部分(颞叶部分)未受胼胝体发展的影响,而较好发育,形成海马和齿状回。海马嵴和原来皮质的分界沟——海马沟(裂),将海马结构与相邻皮质分开,在颞叶它插入海马旁回与海马结构之间;在胼胝体上方,它插入扣带回与灰被之间改称胼胝体沟(callosal sulcus)。脉络裂在大脑半球内侧面形成弯曲,前起自室间孔,沿穹隆上外侧与胼胝体下方 弯曲向下,至颞叶行于海马的上内侧。 又由于颞叶皮质的高度扩展,将其内侧由海马嵴衍化的皮质带推向侧脑室下角的底与内侧壁。随着海马沟的加深,内陷部分的皮质突出于侧脑室下角的底,形成海马。海马向背内侧弯曲,到达半球内面,其内侧端又受脉络裂的制约,内侧份再向里弯,形成一个半月形的齿状回。如此海马沟的上唇为齿状回,下唇为下托。下托与海马结构间的过渡区称副下托。皮质区从内嗅区经旁下托、前下托、下托、副下托至海马与 齿状回,逐渐由原始型6层变到3层细胞结构。 内部结构
解剖成年老鼠的海马齿状回
解剖成年老鼠的海马齿状回 Hideo Hagihara 1,2, Keiko Toyama 1,2, Nobuyuki Yamasaki 2,3, Tsuyoshi Miyakawa 1,2,4,5 1 系统医学部门 ,综合医学科学研究所,藤田保健卫生大学 2日本科学技术振兴机构,进化科技的研究核心峰值 3 精神病学部门,京都府立医药大学, 4 遗传工程和基因组功能学,医药水平机构,京都医学院大学 5 基因分析行为的中心,国家生理科学研究所,国家自然科学研究所 摘要 海马是脑部被广泛研究的部位之一,因为它在人多学习和记忆过程中具有非常重要的作用,其显著的神经细胞可塑性,和参与癫痫、神经退行性疾病,及精神疾病。海马区很明显;主要包括颗粒神经元和包括锥体神经元的亚扪人的角, 这两个区域都通过解刨学和功能性电路相联系。许多不同的mRNA 和蛋白质在齿状回中选择性表达,并且齿状回是神经形成的位置;新的神经元在成熟齿状回中持续产生。在调查mRNA 和蛋白质在齿状回特殊的表达时,激光捕获显微切割技术经常被用到。这个方法有很多限制,例如特殊的仪器和复杂的操作过程。在这段视频记忆中,论证了用立体显微镜从成年老鼠中移出齿状回的解剖技术。齿状回样品组织有很多条件。mRNA 表达TDO2和Dsp 在齿状回中都有很高的水平,但是比着在亚扪人的角中Mrg1b 和Tyro3却在齿状回中水平很低。为了证明该方法的优势,我们用了DNA 微阵列分析中用的整个海马和齿状回。与野生型的老鼠相比,mRNA 在齿状回和aopha-CaMKLL+/-老鼠中选择性表达TDO2和Dsp 各自有0.037和0.10倍的改变。然而在孤立的齿状回和野生型老鼠相比中这些表达出现了0.011和0.021倍的变化,这说明了基因表达的变化可以被更精确更敏感的检测。综上所属这个解刨技术可以方便、准确、可靠的用于重点研究齿状回。 1、 仔细解剖麻醉的老鼠的大脑,放在冷冻环境中的磷酸盐缓冲盐水中。 2、 解剖中脑小脑和后脑 3、 如何解剖海马区 实时定量聚合酶链反应Quantitative real-time PCR