微生物的产能代谢

5.2微生物的产能代谢
代谢(metabolism)是细胞内发生的各种化学反应的总称,它主要由分解代谢(catabolism)和合成代谢(anabolism)两个过程组成。
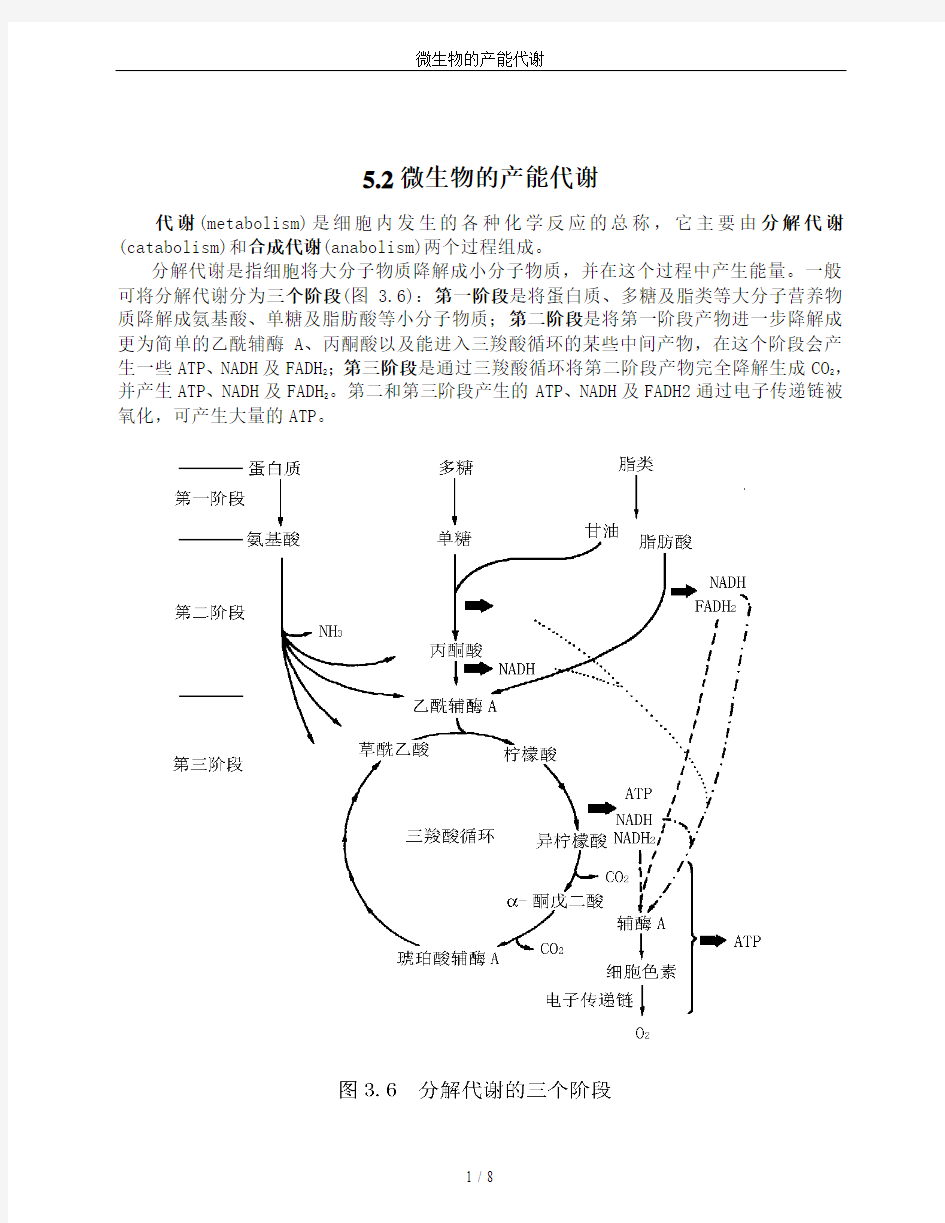
分解代谢是指细胞将大分子物质降解成小分子物质,并在这个过程中产生能量。一般可将分解代谢分为三个阶段(图 3.6):第一阶段是将蛋白质、多糖及脂类等大分子营养物质降解成氨基酸、单糖及脂肪酸等小分子物质;第二阶段是将第一阶段产物进一步降解成更为简单的乙酰辅酶A、丙酮酸以及能进入三羧酸循环的某些中间产物,在这个阶段会产
生一些ATP、NADH及FADH
2;第三阶段是通过三羧酸循环将第二阶段产物完全降解生成CO
2
,
并产生ATP、NADH及FADH
2
。第二和第三阶段产生的ATP、NADH及FADH2通过电子传递链被氧化,可产生大量的ATP。
合成代谢是指细胞利用简单的小分子物质合成复杂大分子的过程,在这个过程中要消耗能量。合成代谢所利用的小分子物质来源于分解代谢过程中产生的中间产物或环境中的小分子营养物质。
在代谢过程中,微生物通过分解代谢产生化学能,光合微生物还可将光能转换成化学能,这些能量除用于合成代谢外,还可用于微生物的运动和运输,另有部分能量以热或光的形式释放到环境中去。微生物产生和利用能量及其与代谢的关系见图3.7。
5.2.1生物氧化:
分解代谢实际上是物质在生物体内经过一系列连续的氧化还原反应,逐步分解并释放能量的过程,这个过程也称为生物氧化,是一个产能代谢过程。在生物氧化过程中释放的能量可被微生物直接利用,也可通过能量转换储存在高能化合物(如ATP)中,以便逐步被利用,还有部分能量以热的形式被释放到环境中。不同类型微生物进行生物氧化所利用的物质是不同的,异养微生物利用有机物,自养微生物则利用无机物,通过生物氧化来进行产能代谢。
1.异养微生物的生物氧化
异养微生物氧化有机物的方式,根据氧化还原反应中电子受体的不同可分成发酵和呼吸两种类型,而呼吸又可分为有氧呼吸和无氧呼吸两种方式。
(1)发酵
发酵(fermentation)是指微生物细胞将有机物氧化释放的电子直接交给底物本身未完全氧化的某种中间产物,同时释放能量并产生各种不同的代谢产物。在发酵条件下有机化合物只是部分地被氧化,因此,只释放出一小部分的能量。发酵过程的氧化是与有机物的还原偶联在一起的。被还原的有机物来自于初始发酵的分解代谢,即不需要外界提供电子受体。
发酵的种类有很多,可发酵的底物有糖类、有机酸、氨基酸等,其中以微生物发酵葡萄糖最为重要。生物体内葡萄糖被降解成丙酮酸的过程称为糖酵解(glycolysis),主要分为四种途径:EMP 途径、HMP 途径、ED 途径、磷酸解酮酶途径。
i EMP 途径(糖酵解途径)
整个EMP 途径大致对分为两个阶段。第一阶段可认为是不涉及氧化还原反应及能量释放的准备阶段,只是生成两分子的主要中间代谢产物:甘油醛-3-磷酸。第二阶段发生氧化还原反应,合成ATP 并形成两分子的丙酮酸。
O H H NADH ATP NAD ADP Pi 222222222++++→+++++丙酮酸葡萄糖
EMP 途径可为微生物的生理活动提供ATP 和NADH ,其中间产物又可为微生物的合成代谢提供碳骨架,并在一定条件下可逆转合成多糖。
ii HMP 途径(磷酸戊糖途径,单磷酸己糖途径)
磷酸戊糖途径可分为氧化阶段和非氧化阶段。一个HMP 途径循环的结果为: 6NADPH 3CO 32++--→+磷酸甘油醛葡萄糖Pi
一般认为HMP 途径不是产能途径,而是为生物合成提供大量的还原力(NADPH)和中间代谢产物。
iii ED 途径
一分子葡萄糖经ED 途径最后生成两分了丙酮酸、一分子ATP 、—分子NADPH 和NADH 。ED 途径可不依赖于EMP 和HMP 途径而单独存在,但对于靠底物水平磷酸化获得ATP 的厌氧菌而言,ED 途径不如EMP 途径经济。
总反应式:
C 6H 12O 6+ADP+Pi+NADP ++NA
D + 2CH 3COCOOH+ATP+NADPH+H ++NADH+H +
iv 磷酸解酮酶途径
磷酸解酮酶途径是明串珠菌在进行异型乳酸发酵过程中分解己糖和戊糖的途径。该途径的特征性酶是磷酸解酮酶,根据解酮酶的不同,把具有磷酸戊糖解酮酶的称为PK 途径,把具有磷酸己糖解酮酶的叫HK 途径。
v 丙酮酸代谢的多样性
在糖酵解过程中生成的丙酮酸可被进一步代谢。在无氧条件下,不同的微生物分解丙酮酸后会积累不同的代谢产物。
乙醇发酵
目前发现多种微生物可以发酵葡萄糖产生乙醇,能进行乙醇发酵的微生物包括酵母菌、根霉、曲霉和某些细菌。
根据在不同条件下代谢产物的不同,可将酵母菌利用葡萄糖进行的发酵分为三种类型:在酵母菌的乙醇发酵中,酵母菌可将葡萄糖经EMP 途径降解为两分子丙酮酸,然后丙酮酸脱羧生成乙醛,乙醛作为氢受体使NAD +再生,发酵终产物为乙醇,这种发酵类型称为酵母的一型发酵;但当环境中存在亚硫酸氢钠时,它可与乙醛反应生成难溶的磺化经基乙醛。
由于乙醛和亚硫酸盐结合而不能作为NADH的受氢体,所以不能形成乙醇,迫使磷酸二羟丙酮代替乙醛作为受氢体,生成-磷酸甘油。-磷酸甘油进一步水解脱磷酸而生成甘油,称为酵母的二型发酵;在弱碱性条件下(pH 7.6),乙醛因得不到足够的氢而积累,两个乙醛分子间会发生歧化反应,一分子乙醛作为氧化剂被还原成乙醇,另一个则作为还原剂被氧化为乙酸,氢受体则由磷酸二经丙酮担任,发酵终产物为甘油、乙醇和乙酸,称为酵母的三型发酵。这种发酵方式不能产生能量,只能在非生长的情况下才进行。
不同的细菌进行乙醇发酵时,其发酵途径也各不相同。如运动发酵单胞菌(Zymomonas mobilis)和厌氧发酵单胞菌(Zymomonas anaerobia)是利用ED途径分解葡萄糖为内酮酸,最后得到乙醇,对于某些生长在极端酸性条件下的严格厌氧菌,如胃八叠球菌(arcina ventriculi)和肠杆菌(Enterobacteriaceae)则是利用EMP途径进行乙醇发酵。
乳酸发酵
许多细菌能利用葡萄糖产生乳酸,这类细菌称为乳酸细菌。
根据产物的不同,乳酸发酵有三种类型:同型乳酸发酵、异型乳酸发酵和双歧发酵。
同型乳酸发酵的过程是:葡萄糖经EMP途径降解为丙酮酸,丙酮酸在乳酸脱氢酶的作用下被NADH还原为乳酸。由于终产物只有乳酸一种,故称为同型乳酸发酵。
在异型乳酸发酵中,葡萄糖首先经PK途径分解,发酵终产物除乳酸以外还有一部分乙醇或乙酸。在肠膜明串珠菌(Leuconostoc mesenteroides)中,利用HK途径分解葡萄糖,产生甘油醛-3-磷酸和乙酰磷酸,其中甘油醛-3-磷酸进一步转化为乳酸,乙酰磷酸经两次还原变为乙醇,当发酵戊糖时,则是利用PK途径,磷酸解酮糖酶催化木酮糖-5-磷酸裂解生成乙酰磷酸和甘油醛-3-磷酸。
双歧发酵是两歧双歧杆菌(bifidobacteriium bifidum)发酵葡萄糖产生乳酸的一条途径。此反应中有两种磷酸酮糖酶参加反应,即果糖-6-磷酸磷酸酮糖酶和木酮糖-5-磷酸磷酸酮糖酶分别催化果糖-6-磷酸和木酮糖-5-磷酸裂解产生乙酰磷酸和丁糖-4-磷酸及甘油醛-3-磷酸和乙酰磷酸。
丙酸发酵
许多厌氧菌可进行丙酸发酵。葡萄糖经EMP途径分解为两个丙酮酸后,再被转化为丙酸。少数丙酸细菌还能将乳酸(或利用葡萄糖分解而产生的乳酸)转变为丙酸。
丁酸发酵
某些专性厌氧菌,如梭菌属(Clostridium)、丁酸弧菌属(Butyrivibrio)、真杆菌属(Eubacterium和梭杆菌属(Fusobacterium),能进行丁酸与丙酮-丁醇发酵。在发酵过程中,葡萄糖经EMP途径降解为丙酮酸,接着在丙酮酸-铁氧还蛋白酶的参与下,将丙酮酸转化为乙酰辅酶A。乙酰辅酶A再经一系列反应生成丁酸或丁醇和丙酮。
混合酸发酵
某些肠杆菌,如埃希氏菌属(Escherichia)、沙门氏菌属(SAlmonella)和志贺氏菌属(Shigella) 中的一些菌,能够利用葡萄糖进行混合酸发酵。先通过EMP途径将葡萄糖分解为丙酮酸,然后由不同的酶系将丙酮酸转化成不同的产物,如乳酸、乙酸、甲酸、乙醇、CO2和氢气,还有一部分磷酸烯醇式丙酮酸用于生成琥珀酸;而肠杆菌、欧文氏菌属(Erwinia)中的一些细菌,能将丙酮酸转变成乙酰乳酸,乙酰乳酸经一系列反应生成丁二醇。由于这类肠道菌还具有丙酮酸-甲酸裂解酶,乳酸脱氢酶等,所以其终产物还有甲酸、乳酸、乙醇等。
葡萄糖分子在没有外源电子受体时的代谢过程。在这个过程中,底物中所具有的能量
只有一小部分被释放出来,并合成少量ATP。造成这种现象的原因有两个,一是底物的碳
原子只被部分氧化,二是初始电子供体和最终电子受体的还原电势相差不大。
(2)呼吸作用
如果有氧或其他外源电子受体存在时,底物分子可被完全氧化为CO
2
,且在此过程中可合成的ATP的量大大多于发酵过程。
微生物在降解底物的过程中,将释放出的电子交给NAD(P)+、FAD或FMN等电子载体,再经电子传递系统传给外源电子受体,从而生成水或其他还原型产物并释放出能量的过程,称为呼吸作用。
呼吸作用与发酵作用的根本区别在于:电子载体不是将电子直接传递给底物降解的中间产物,而是交给电子传递系统,逐步释放出能量后再交给最终电子受体。
能通过呼吸作用分解的有机物包括某些碳氢化合物、脂肪酸和许多醇类。
i 有氧呼吸
在呼吸作用中,以分子氧作为最终电子受体的称为有氧呼吸(aerobic respiration)。
在发酵过程中,葡萄糖经过糖酵解作用形成的丙酮酸在厌氧条件下转变成不同的发酵产物,而在有氧呼吸过程中,丙酮酸进入三羧酸循环(tricarboxylic acid cycle,简称
TCA循环),被彻底氧化生成CO
2
和水,同时释放大量能量。
ii 厌氧呼吸
在呼吸作用中,以氧化型化合物作为最终电子受体的称为无氧呼吸(anaerobic respiration)。
某些厌氧和兼性厌氧微生物在无氧条件下进行无氧呼吸。无氧呼吸的最终电子受体不
是氧,而是像NO3-、NO2-、SO
42-、CO
2
等这类外源受体。无氧呼吸也需要细胞色素等电子传递
体,并在能量分级释放过程中伴随有磷酸化作用,也能产生较多的能量用于生命活动。但
由于部分能量随电子转移传给最终电子受体,所以生成的能量不如有氧呼吸产生的多。5.2.2自养微生物的生物氧化
一些微生物可以从氧化无机物获得能量,同化合成细胞物质,这类细菌称为化能自养微生物。它们在无机能源氧化过程中通过氧化磷酸化产生ATP。
氨的氧化
NH
3
同亚硝酸(NO2-)是可以用作能源的最普通的无机氮化合物,能被硝化细菌所氧化,硝化细菌可分为两个亚群:亚硝化细菌和硝化细菌。
氨氧化为硝酸的过程可分为两个阶段,先由亚硝化细菌将氨氧化为亚硝酸,再由硝化细菌将亚硝酸氧化为硝酸。由氨氧化为硝酸是通过这两类细菌依次进行的。
硝化细菌都是一些专性好氧的革兰氏阳性细菌,以分子氧为最终电子受体,且大多数是专性天机营养型。它们的细胞都具有复杂的膜内褶结构,这有利于增加细胞的代谢能力。硝化细菌无芽孢,多数为二分裂殖,生长缓慢,平均代时在10h以上,分布非常广泛。
硫的氧化
硫杆菌能够利用一种或多种还原态或部分还原态的硫化合物(包括硫化物、元素硫、硫
代硫酸盐、多硫酸盐和亚硫酸盐)作能源。H
2
S首先被氧化成元素硫,随之被硫氧化酶和细胞色素系统氧化成亚硫酸盐,放出的电子在传递过程中可以偶联产生四个ATP。亚硫酸盐
的氧化可分为两条途径,一是直接氧化成SO
4
2-的途径,由亚硫酸盐—细胞色素c还原酶和末端细胞色素系统催化,产生一个ATP;二是经磷酸腺苷硫酸的氧化途径,每氧化一分子
SO
3
2-产生2.5个ATP。
铁的氧化
从亚铁到高铁状态的铁的氧化,对于少数细菌来说也是一种产能反应,但从这种氧化中只有少量的能量可以被利用。亚铁的氧化仅在嗜酸性的氧化亚铁硫杆菌(Thiobacillus ferrooxidans)中进行了较为详细的研究。在低pH环境中这种菌能利用亚铁氧化时放出的能量生长。在该菌的呼吸链中发现了—种含铜蛋白质(rusticyanin),它与几种细胞色素c 氧化酶构成电子传递链。虽然电子传递过程中的放能部位和放出有效能和一种细胞色素a
1
的多少还有待研究,但已知在电子传递到氧的过程中细胞质内有质了消耗,从而驱动ATP 的合成。
氢的氧化
氢细菌都是一些呈革兰氏阴件的兼性化能自养菌。它们能利用分子氢氧化产生的能量同化CO2,也能利用其他有机物生长。氢细菌的细胞膜上有泛醌、维生素K2及细胞色素等呼吸链组分。在该菌中,电子直接从氢传递给电子传递系统,电于在呼吸链传递过程中产生ATP。在多数氢细菌中有两种与氢的氧化有关的酶。一种是位于壁膜间隙或结合在细胞质膜上的不需NAD+的颗粒状氧化酶,它能够催化以下反应:
++
-
2
H2
→e
H
2
该酶在氧化氢并通过电子传递系统传递电子的过程中,可驱动质子的跨膜运输,形成跨膜质子梯度为ATP的合成提供动力;另一种是可溶性氢化酶,它能催化氢的氧化,而使
的还原。
NAD+还原的反应。所生成的NADH主要用于CO
2
5.2.3能量转换
在产能代谢过程中,微生物通过底物水平磷酸化和氧化磷酸化将某种物质氧化而释放的能量储存于ATP等高能分子中,对光合微生物而言,则可通过光合磷酸化将光能转变为化学能储存于ATP中。
底物水平磷酸化(substratelevelphosphorylation)
物质在生物氧化过程中,常生成一些含有高能键的化合物,而这些化合物可直接偶联ATP或GTP的合成,这种产生ATP等高能分子的方式称为底物水平磷酸化。底物水平磷酸化既存在于发酵过程小,也存在于呼吸作用过程中。例如,在EMP途径中,1,3-二磷酸甘油酸转变为3-磷酸甘油酸以及磷酸烯醇式丙酮酸转变为丙酮酸的过程中都分别偶联着一分子ATP的形成;在三羧酸循环过程中,琥珀酰辅酶A转变为琥珀酸时偶联着一分子GTP的形成。
氧化磷酸化(oxidative phosphorylation)
可通过位于线粒体内膜和细菌质膜上的电物质在生物氧化过程中形成的NADH和FADH
2
子传递系统将电子传递给氧或其他氧化型物质,在这个过程中偶联着ATP的合成,这种产
可分别产生3个和2个ATP。
生ATP的方式称为氧化磷酸化。一分子NADH和FADH
2
光合磷酸化(photophosphorylation)
光合作用是自然界一个极其重要的生物学过程,其实质是通过光合磷酸化将光能转变成化学能,以用于从CO2合成细胞物质。行光合作用的生物体除了绿色植物外,还包括光合微生物,如藻类、蓝细菌和光合细菌(包括紫色细菌、绿色细菌、嗜盐菌等)。它们利用光能维持生命,同时也为其他生物(如动物和异养微生物)提供了赖以生存的有机物。
i 光合色素
光合色素是光合生物所特有的色素,是将光能转化为化学能的关键物质。共分三类:叶绿素 (chl)或细菌叶绿素(Bchl),类胡萝卜素和藻胆素。除光合细菌外,叶绿素a普遍
存在于光合生物中,叶绿素a 、b 共同存在于高等植物、绿藻和蓝绿细菌中,叶绿素c 存在于褐藻和硅藻中,叶绿素d 存在于红藻中,叶绿素e 存在于金黄藻中,褐藻和红藻也含有叶绿素a 。细菌叶绿素具有和高等植物中的叶绿素相类似的化学结构,两者的区别在于侧链基团的不同,以及由此而导致的光吸收特性的差异。此外,叶绿素和细菌叶绿素的吸收光谱在不同的细胞中也有差异。
所有光合生物都有类胡萝卜素。类胡萝卜素虽然不直接参加光合反应,但它们有捕获光能的作用,能把吸收的光能高效地传给细菌叶绿素(或叶绿素)。而且这种光能同叶绿素(或细菌叶绿素)直接捕捉到的光能一样被用来进行光合磷酸化作用。此外胡萝卜素还有两个作用:一是可以作为叶绿素所催化的光氧化反应的淬灭剂,以保护光合机构不受光氧化损伤,二是可能在细胞能量代谢方面起辅助作用。
藻胆素因具有类似胆汁的颜色而得名,其化学结构与叶绿素相似,都含有四个毗咯环,但藻胆素没有长链植醇基,也没有镁原子,而且四个毗咯环是直链的。
ii 光合单位
以往将在光合作用过程中还原一分子CO 2所需的叶绿素分子数称为光合单位
(phutosynthetic units)。后来通过分析紫色细菌载色体的结构,获得了对光合单位的进一步认识。光合色素分布于两个“系统”,分别称为“光合系统Ⅰ”和“光合系统Ⅱ”。每个系统即为一个光合单位。这两个系统中的光合色素的成分和比例不同。一个光合单位由一个光捕获复合体和一个反应中心复合体组成。光捕获复合体含有菌绿素和类胡萝卜素,它们吸收一个光子后,引起波长最长的菌绿素(P870)激活,从而传给反应中心,激发态的P870可释放出一个高能电子。
iii 光合磷酸化
光合磷酸化是指光能转变为化学能的过程。当一个叶绿素分子吸收光量子时,叶绿素性质上即被激活,导致叶绿素(或细菌叶绿素)释放——个电子而被氧化,释放出的电子在电子传递系统中的传递过程中逐步释放能量,这就是光合磷酸化的基本动力。
环式光合磷酸化
光合细菌主要通过环式光合磷酸化作用产生ATP ,这类细菌主要包括紫色硫细菌、绿色硫细菌、紫色非硫细菌和绿色非硫细菌。在光合细菌中,吸收光量子而被激活的细菌叶绿素释放出高能电子,于是这个细菌叶绿素分子即带有正电荷。所释放出来的高能电子顺序通过铁氧还蛋白、辅酶Q 、细胞色素b 和c ,再返回到带正电荷的细菌叶绿素分子。在辅酶Q 将电子传递给细胞色素c 的过程中,造成了质子的跨膜移动,为ATP 的合成提供了能量。在这个电子循环传递过程中,光能转变为化学能,故称环式光合磷酸化。环式光合磷酸化可在厌氧条件下进行,产物只有ATP ,无NADP(H),也不产生分子氧。通常以下式表示环式光合磷酸化作用:
nATP hv Bchl nPi NADP
非环式光合磷酸化
高等植物和蓝细菌与光合细菌不同,它们可以裂解水,以提供细胞合成的还原能力。它们含有两种类型的反应中心,连同天线色素、初级电子受体和供体一起构成了光合系统Ⅰ和光合系统Ⅱ,这两个系统偶联,进行非环式光合磷酸化。在光合系统Ⅰ中,叶绿素分子P7M 吸收光于后被激活,释放出一个高能电子。这个高能电子传递给铁氧还蛋白(Fd),并使之被还原。还原的扶氧还蛋白在Fd :NADP +还原酶的作用下,将NADP +还原为NADPH 。
用以还原P 700的电子来源于光合系统Ⅱ。在光合系统Ⅱ中,叶绿素分子P 680吸收光子后,释
放出一个高能电子。后者先传递给辅酶Q ,再传给光合系统Ⅰ,使P 700还原。失去电子的P 680,靠水的光解产生的电子来补充。高能电子从辅酶Q 到光合系统Ⅰ的过程巾,可推动ATP 的合成。非环式光合磷酸化的反应式为:
有的光合细菌虽然只有一个光合系统,但也以非环式光合磷酸化的方式合成ATP ,如绿硫细菌和绿色细菌。从光反应中心释放出的高能电子经铁硫蛋白、铁氧还蛋白、黄素蛋白,最后用于还原NAD +生成NADH 。反应中心的还原依靠外源电子供体,如S 2-、S 2O 32-等。外
源电子供体在氧化过程中放出电子,经电子传递系统传给失去了电子的光合色素,使其还原,同时偶联ATP 的生成。由于这个电子传递途径也没有形成环式回路,故也称为非环式光合磷酸化,反应式为:
222222222O ATP H NADPH O H Pi ADP NADP +++→+++++S ATP H NADPH hv chl Pi ADP H NAD ++++++++2
(完整版)微生物学第六章微生物的代谢
第十四授课单元 一、教学目的 使学生了解呼吸与发酵作用,重点讲解微生物代谢的特殊性,联系在食品和发酵生产上的应用,注意体现微生物不同发酵类型及代谢的特点。 二、教学内容(第六章微生物的新陈代谢 第一节微生物的产能代谢) 1. 代谢概论简单介绍新陈代谢的概念,同化作用和异化作用。 2. 微生物的产能代谢:重点介绍化能异养微生物生物氧化的三种产能方式,即发酵、有氧呼吸和无氧呼吸, 3. 介绍乙醇发酵(酵母菌的乙醇发酵途径和运动发酵单胞菌的乙醇发酵途径)、乳酸发酵(同型乳酸发酵和异型乳酸发酵)、甘油发酵、丙酮丁醇发酵、混合酸发酵及丁二醇发酵; 三、教学重点、难点及处理方法 重点:化能异养微生物生物氧化的三种产能方式,即发酵、有氧呼吸和无氧呼吸,介绍乙醇发酵(酵母菌的乙醇发酵途径和运动发酵单胞菌的乙醇发酵途径)、乳酸发酵(同型乳酸发酵和异型乳酸发酵)、甘油发酵、丙酮丁醇发酵、混合酸发酵及丁二醇发酵;由于学生在生物化学课程中已经学过各种代谢途径,因此在微生物学中不再作为重点讲解。本章内容主要使学生了解呼吸与发酵作用,重点讲解微生物代谢的特殊性,联系在食品和发酵生产上的应用,注意体现微生物不同发酵类型及代谢的特点。 难点: 化能异养微生物生物氧化的三种产能方式,即发酵、有氧呼吸和无氧呼吸的区别。尤其是发酵的概念, 学生只是在现实生活中知道这个名词, 但是不清楚其确切的生物学含义, 指在能量代谢或生物氧化中,在无氧条件下,底物(有机物)氧化释放的氢(或电子)不经呼吸链传递,而直接交给某种未完全氧化的中间产物的一类低效产能过程。实质: 底物水平磷酸化产生ATP. 另外, 联系食品和发酵生产上应用的发酵类型及代谢特点更有助于学生理解发酵的概念实质及发酵的特点. 有氧呼吸与无氧呼吸的概念, 并介绍无氧呼吸中硝酸根(反硝化作用)、硫酸根作为最终电子受体的呼吸特点,介绍不同呼吸类型的微生物。介绍化能自养微生物的生物氧化特点,光能自养微生物的光合磷酸化途径(循环光合磷酸化、非循环光合磷酸化和嗜盐菌紫膜的光合作用)。 四、板书设计 第六章微生物的新陈代谢 第一节代谢概论 能量代谢的中心任务,是生物体如何把外界环境中的多种形式的 最初能源转换成对一切生命活动都能使用的通用能源------ATP。 这就是产能代谢。 有机物化能异养微生物 最初 能源还原态无机物化能自养微生物通用能源ATP 日光: 光能营养微生物 第二节糖的代谢
常见微生物检验项目及临床意义
细菌培养与其它检验项目的临床意义 卫生部规定接受抗菌药物治疗的住院患者微生物检验样本送检率不低于30%(卫办医政发〔2011〕56号)。特介绍微生物检验项目,方便统计与分析点评。细菌检验为感染性疾病诊断和治疗提供依据: 目标性治疗:提高微生物的送检率与检出率,有利于诊断与使用抗菌药。 经验治疗:根据本地区病原菌类型时间、区域、耐药谱等使用抗菌药。 调整治疗方案针对性用药:获得准确病原菌和细菌药敏结果。 细菌培养的临床意义 细菌培养与其它检验项目不同,由于其样本采集易受杂菌的干扰和培养条件的限制,因而造成检测结果有时与临床不完全一致,故在分析细菌培养报告时应明白:细菌培养阴性不代表无细菌感染、细菌培养阳性不代表该菌一定是病原菌,应结合患者具体情况而定。 1、血液和骨髓培养 目前血液培养仍然是菌血症和败血症的细菌学检验的基本方法,并且广泛地应用于伤寒、副伤寒及其它细菌引起的败血症的诊断。菌血症系病原菌一时性或间断地由局部进入血流,但并不在血中繁殖,无明显血液感染临床征象。常可发生在病灶感染或牙齿感染,尤以拔牙、扁桃体切除及脊髓炎手术后等多见。败血症是指病原菌侵入血流,并在其中大量生长繁殖,造成身体的严重损害,引起显著的全身症状(如不规则高热与全身中毒等症状),它多继发于组织器官感染,尤其是当机体免疫功能低下、广谱抗生素和激素的应用及烧伤等。 单次的血培养结果,对临床无鉴别指导意义,应多次多部位采集血液进行培养,才可判定检出菌究竟是病原菌还是污染菌。 抽血时应特别注意皮肤消毒和培养瓶口的灭菌。 2、脑脊液培养 正常人的脑脊液是无菌的,故在脑脊液中检出细菌(排除操作中的污染)应视为病原菌。引起脑膜炎的细菌种类不同,化脓性脑膜炎病原菌多为脑膜炎奈瑟菌,除此之外尚有肺炎链球菌、流感嗜血杆菌、金黄色葡萄球菌、大肠埃希菌、铜绿假单胞菌等。 3、尿液培养 尿液细菌培养对于膀胱和肾脏感染的及早发现和病原学诊断很有价值,对于尿道、前列腺以及内外生殖器的炎症也有一定价值。尿液中出现细菌通常称为菌尿症,如果尿液本身澄清但培养出细菌,一般为标本采集时未彻底消毒尿道口引起。如果细菌培养阳性同时伴有脓尿出现,则提示有尿路感染的可能(需指出轻度感染时可无脓尿出现)。泌尿系感染常见菌为大肠埃希菌、葡萄球菌、链球菌、变形杆菌等,除结核分枝支杆菌外,这些细菌又是尿道口常驻菌,极易引起标本留样污染,应注意鉴别。某些真菌疾病,如曲菌病在播散时也可造成肾脏感染,且尿中也能检查到。
微生物营养与培养基答案
1.培养基应具备微生物生长所需要的六大营养要素是_碳源___、__氮源__、__能源__、___无机盐___、__生长因子__和???????????____水___。 2.碳源物对微生物的功能是__提供碳素来源__和__能量来源__,微生物可用的碳源物质主要有___糖类_、___有机酸_、__脂类_、__烃__、__ CO2及碳酸盐__等。 3.微生物利用的氮源物质主要有_蛋白质_、_铵盐_、_硝酸盐__、_分子氮__、__酰胺_等,而常用的速效N源如__玉米粉__,它有利于___菌体生长___;迟效N源如__黄豆饼粉__、__花生饼粉_,它有利于___代谢产物的形成______。 4.无机盐对微生物的生理功能是__作为酶活性中心的组成部分_、__维持生物大分子和细胞结构的稳定性_____ 、_调节并维持细胞的渗透压平衡__ 和 _控制细胞的氧化还原电位和作为某些微生物生长的能源物质等_。 5.微生物的营养类型可分为__光能无机自养型__、__光能有机异养型__、_化能无机自养型和_化能有机异养型_。微生物类型的可变性有利于_提高微生物对环境条件变化的适应能力_。 6.生长因子主要包括_维生素_、__氨基酸_和__嘌呤及嘧啶_,它们对微生物所起的作用是__作为酶的辅基或辅酶参与新陈代谢_、_维持微生物正常生长_、_为合成核柑、核苷酸和核酸提供原料__。 7.在微生物研究和生长实践中,选用和设计培养基的最基本要求是__选择适宜的营养物质_、_营养物的浓度及配比合适_、_物理、化学条件适宜_、_经济节约_和__精心设计、试验比较_。 8.液体培养基中加入CaCO3的目的通常是为了__调节培养基的pH值___。 9.营养物质进入细胞的方式有__单纯扩散__、__促进扩散__、_主动运输__和___基团移位_,而金黄色葡萄球菌是通过___主动运输__方式运输乳糖,大肠杆菌又是通过_基团移位__方式运输嘌呤和嘧啶的。 10.影响营养物质进入细胞的主要因素是_营养物质本身__、__微生物所处的环境__和___微生物细胞的透过屏障___。 11.实验室常用的有机氮源有__蛋白胨__和__牛肉膏__等,无机氮源有__硫酸铵__和_硝酸钠等。为节约成本,工厂中常用___豆饼粉__等作为有机氮源。 12.培养基按用途分可分为基础培养基、增殖培养基、鉴别培养基和选择培养基四种类型。 二.是非题 1、在固体培养基中,琼脂的浓度一般为—%.(×)
代谢组学的研究方法和研究流程
代谢组学的研究方法和研究流程分子微生物学112300003林兵 随着人类基因组计划等重大科学项目的实施,基因组学、转录组学及蛋白质组学在研究人类生命科学的过程中发挥了重要的作用,与此同时, 代谢组学(metabolomics)在20世纪90年代中期产生并迅速地发展起来,与基因组学、转录组学、蛋白质组学共同组成系统生物学。基因组学、转录组学、蛋白质组学和代谢组学等各种组学0在生命科学领域中发挥了重要的作用,它们分别从调控生命过程的不同层面进行研究, 使人们能够从分子水平研究生命现象, 探讨生命的本质, 逐步系统地认识生命发展的规律.这些组学手段加上生物信息学, 成为系统生物学的重要组成部分。 代谢组学的出现和发展是必要的, 同时也是必须的。对于基因组学和蛋白质组学在生命科学研究中的缺点和不足, 代谢组学正好可以进行弥补。代谢组学研究的是生命个体对外源性物质(药物或毒物)的刺激、环境变化或遗传修饰所做出的所有代谢应答, 并且检测这种应答的全貌及其动态变化。代谢组学方法为生命科学的发展提供了有力的现代化实验技术手段, 同时也为新药临床前安全性评价与实践提供了新的技术支持与保障. 1 代谢组学的概念及发展 代谢组学最初是由英国帝国理工大学Jeremy N icholson教授提出的,他认为代谢组学是将人体作为一个完整的系统,机体的生理病理过程作为一个动态的系统来研究, 并且将代谢组学定义为生物体对病理生理或基因修饰等刺激产生的代谢物质动态应答的定量测定。2000年,德国马普所的Fiehn等提出了代谢组学的概念,但是与N ichols on提出的代谢组学不同, 他是将代谢组学定位为一个静态的过程,也可以称为/代谢物组学, 即对限定条件下的特定生物样品中所有代谢产物的定性定量分析。同时Fiehn还将代谢组学按照研究目的的不同分为4类: 代谢物靶标分析,代谢轮廓(谱)分析, 代谢组学,代谢指纹分析。现在代谢组学在国内外的研究都在迅速地发展, 科学家们对代谢组学这一概念也进行了完善, 作出了科学的定义: 代谢组学是对一个生物系统的细胞在给定时间和条件下所有小分子代谢物质的定性定量分析,从而定量描述生物内源性代谢物质的整体及其对内因和外因变化应答规律的科学。 与基因组学、转录组学、蛋白质组学相同, 代谢组学的主要研究思想是全局观点。与传统的代谢研究相比, 代谢组学融合了物理学、生物学及分析化学等多学科知识, 利用现代化的先进的仪器联用分析技术对机体在特定的条件下整个代谢产物谱的变化进行检测,并通过特殊的多元统计分析方法研究整体的生物学功能状况。由于代谢组学的研究对象是人体或动物体的所有代谢产物, 而这些代谢产物的产生都是由机体的内源性物质发生反应生成的,因此,代谢产物的变化也就揭示了内源性物质或是基因水平的变化,这使研究对象从微观的基因变为宏观的代谢物,宏观代谢表型的研究使得科学研究的对象范围缩小而且更加直观,易于理解, 这点也是代谢组学研究的优势之一. 代谢组学的优势主要包括:对机体损伤小,所得到的信息量大,相对于基因组学和蛋白质组学检测更加容易。由于代谢组学发展的时间较短, 并且由于代谢组学的分析对象是无偏向性的样品中所有的小分子物质,因此对分析手段的要求比较高, 在数据处理和模式识别上也不成熟,存在一些不足之处。同时生物体代谢物组变化快, 稳定性较难控制,当机体的生理和药理效应超敏时,受试物即使没有相关毒性,也可能引起明显的代谢变化,导致假阳性结果。 代谢组学应用领域大致可以分为以下7个方面:
微生物遗传学习题及答案(第二章)
遗传的物质基础 1、解词 多组分基因组(segmented genome):在一些RNA病毒中,RNA分子的容量有限,如果要增加遗传信息量,则需将病毒的基因组分段保存在2个或多个RNA片段中,以在病毒粒子中形成2个或多个RNA分子,此类病毒中的这些遗传物质称为多组分基因组。 多分体:在不同病毒粒子中含有不同的RNA片段,只有几种含有基因组中不同RNA 片段的病毒粒子同时存在时才能表现有效的侵染,在某些植物RNA病毒中存在这种多分体现象。 类病毒:一种小分子单链环状RNA分子,无蛋白质外壳保护,结构和化学组成比普通病毒简单,不需要辅助病毒便可侵入敏感的宿主细胞内进行自我复制,并使宿主致病或死亡。 朊病毒(Protein infection,Prion):一类侵染动物并在寄主细胞内复制的小分子无免疫性的疏水蛋白质,这类蛋白质能与寄主脑组织中的核酸相互作用,使脑组织海绵状损伤,引起动物的亚急性海绵样脑病。 重叠基因:具有部分公用核苷酸序列的基因,即同一段DNA携带了两种或两种以上不同蛋白质的编码信息。重叠的部分可在调控区或结构基因区,常见于病毒和噬菌体基因组中。 串珠结构:60bp的间隔线状DNA双链作为连接丝,将许多核小体串联起来并盘绕形成的染色质纤维细丝,呈念珠状,即为染色质的串珠结构。 核小体(nucleosome):由H2A、H2B、H3、H4四种组蛋白各以两个分子组成的八聚体核心和一分子组蛋白H1以及大约200bp的DNA缠绕而组成,直径一般为10nm。2、问题 Ⅰ、简述病毒、原核生物和真核生物遗传物质的特点。 病毒:核酸类型有DNA和RNA之分;核酸分子有单链和双链之分;空间结构有开放型和闭合型之分;基因组有多组份型和单组份型;有多分体现象;能够指导蛋白质合成;能够产生可遗传变异。 原核生物:原核微生物遗传物质分子量较病毒大而比真核微生物小,DNA与微量的组蛋白相结合,形成超螺旋脚手架结构;某些细菌只有一条环状双链DNA,某些拥有两个环状DNA,有些则一条环状、一条线状DNA;能够指导蛋白质合成;能够产生可遗传变异;一般情况下,一个细菌细胞只有一套基因组,其DNA含量在细胞间期十分稳定;能够自我复制,使亲子代之间保持连续性;基因组在DNA上一般是连续排列。 真核生物:真核微生物遗传物质主要存在于细胞核,细胞核有核膜包裹,核内存在多条线状dsDNA;DNA和组蛋白组成核小体,线状DNA双链缠绕在核小体上形成串珠状染色质;每一染色体只含有一条线状双链DNA;分子结构相对稳定,能够自我复制,使亲子代之间保持连续性;能够指导蛋白质合成;能够产生可遗传变异;基因组含有大量的重复序列。 Ⅱ、原核生物和真核生物染色体外遗传物质。 答:染色体外遗传物质是细胞的非固定成分,也能影响细胞的代谢活动,但它们不是细胞生存必不可少的组成部分,包括附加体和共生体。含有DNA的细胞质颗粒,即附加体,既能以完全自主的状态存在,也能组入到染色体上,成为染色体的一部分。进入细胞,与细胞建立起特殊的共生关系的一类物质即共生体。 原核生物的染色体外遗传物质:附加体如R质粒(抗性因子,使E. coli.抗一定浓度的抗菌素)、F因子(决定性别,有F因子的E. coli.为雄性------供体)等;共生体如
微生物的代谢及其调控
微生物的代谢及其调控
1微生物的代谢 微生物代谢包括微生物物质代谢和能量代谢。 1.1微生物物质代谢 微生物物质代谢是指发生在微生物活细胞中的各种分解代谢与合成代谢的总和。 1.1.1分解代谢 分解代谢是指细胞将大分子物质降解成小分子物质,并在这个过程中产生能量。—般可将分解代谢分为TP。三个阶段:第一阶段是将蛋白质、多糖及脂类等大分子营养物质降解成氨基酸、单糖及脂肪酸等小分子物质;第二阶段是将第一阶段产物进一步降解成更为简单的乙酰辅酶A、丙酮酸以及能进入三羧酸循环的某些中间产物,在这个阶段会产生一些ATP、NADH及FADH2;第三阶段是通过三羧酸循环将第二阶段产物完全降解生成CO2,并产生ATP、NADH 及FADH2。第二和第三阶段产生的ATP、NADH及FADH2通过电子传递链被氧化,可产生大量的ATP。 1.1.1.1大分子有机物的分解 (1)淀粉的分解 淀粉是许多种微生物用作碳源的原料。它是葡萄糖的多聚物,有直链淀粉和支链淀粉之分。一般天然淀粉中,直链淀粉约占20%,支链淀粉约占80%。直链淀粉为α一l、4糖苷键组成的直链分子;支链淀粉只是在支点处由α—1、6糖苷键连接而成。 微生物对淀粉的分解是由微生物分泌的淀粉酶催化进行的。淀粉酶是一类水解淀粉糖苷键酶的总称。它的种类很多,作用方式及产物也不尽相同,主要有液化型淀粉酶、糖化型淀粉酶(包括β—淀粉酶、糖化酶、异淀粉酶)。 以液化型淀粉酶为例,这种酶可以任意分解淀粉的。α-l、4糖苷键,而不能分解α-1、6糖苷键。淀粉经该酶作用以后,黏度很快下降,液化后变为糊精,最终产物为糊精、麦芽糖和少量葡萄糖。由于这种酶能使淀粉表现为液化,淀
代谢组学在医药领域的应用与进展
代谢组学在医药领域的应用与进展 一、学习指导 1.学习代谢组学的概念及内涵,掌握代谢组学的研究对象与分析方法。 2.熟悉代谢组学数据分析技术手段 3.了解代谢组学优势特点 4.了解代谢组学在医药领域的应用 5.了解代谢组学发展趋势 二、正文 基因组功能解析是后基因组时代生命科学研究的热点之一,由于基因功能的复杂性和生物系统的完整性,必然要从“整体”层面上来理解构成生物体系的各个模块功能。随着新的测量技术、高通量的分析方法、先进的信息科学和系统科学新理论的发展,加上生物学研究的深入和生物信息的大量积累,使得在系统水平上研究由分子生物学发现的组件所构成的生命体系成为可能[1]。系统生物学家们认为,将生命科学上升为“综合”科学的时机已经成熟,生命科学再次回到整合性研究的新高度,逐步由分子生物学时代进入到系统生物学时代[2]。系统生物学不同以往的实验生物学仅关注个别基因和蛋白质,它要研究所有基因、蛋白质,代谢物等组分间的所有相互关系,通过整合各组成成分的信息,以数学方法建立模型描述系统结构[3,4]。 (一)代谢组学的概念及内涵 代谢组学是继基因组学、转录组学和蛋白质组学之后,系统生物学的重要组成部分,也是目前组学领域研究的热点之一。代谢组学术语在国际上有两个英文名,即metabolomics 和metabonomics。Metabolomics是由德国的植物学家Fiehn等通过对植物代谢物研究提出来的,认为代谢组学(metabolomics)是定性和定量分析单个细胞或单一类型细胞的代谢调控和代谢流中所有低分子量代谢产物,从而监测机体或活细胞中化学变化的一门科学[5]。英国Nicholson研究小组从毒理学角度分析大鼠尿液成份时提出了代谢组学(Metabonomics)的概念,认为代谢组学是通过考察生物体系受扰动或刺激后(如某个特定基因变异或环境变化后),其代谢产物的变化或代谢产物随时间的变化来研究生物体系的代谢途径的一种技术[6]。国内的代谢组学研究小组基本用metabonomics一词来表示“代谢组学”。严格地说,代谢组学所研究的对象应该包括生物系统中所有的代谢产物。但由于实际分析手段的局限性,只对各种代谢路径底物和产物的小分子物质(MW<1Kd)进行测定和分析。 (二)代谢组学优势特点 代谢组学作为系统生物学的一个重要组成部分,代谢组可以更好地反映体系表型生物机体是一个动态的、多因素综合调控的复杂体系,在从基因到性状的生物信息传递链中,机体需通过不断调节自身复杂的代谢网络来维持系统内部以及与外界环境的正常动态平衡[7]。
我国在微生物代谢领域的研究现状及展望
我国在微生物代谢领域的研究现状及展望 发表时间:2012-06-18T14:33:59.827Z 来源:《赤子》2012年第8期供稿作者:李夏 [导读] 微生物代谢是指微生物吸收营养物质维持生命和增殖并降解基质的一系列化学反应过程,包括有机物的降解和微生物的增殖。 李夏(四川化工职业技术学院,四川泸州 646005) 摘要:微生物代谢是指微生物吸收营养物质维持生命和增殖并降解基质的一系列化学反应过程,包括有机物的降解和微生物的增殖。在分解代谢中,有机物在微生物作用下,发生氧化、放热和酶降解过程,使结构复杂的大分子降解;合成代谢中,微生物利用营养物及分解代谢中释放的能量,发生还原吸热及酶的合成过程,使微生物生长增殖。文章主要介绍我国在微生物代谢领域的研究现状及对未来的展望,为我们呈现了一个广阔的微生物代谢世界。 关键词:微生物代谢;分解代谢;合成代谢;研究现 前言 微生物在生长过程中机体内的复杂代谢过程是互相协调和高度有序的,并对外界环境的改变能够迅速做出反应。其原则是经济合理地利用和合成所需要的各种物质和能量,使细胞处于平衡生长状态。在实际生产中,往往需要高浓度的积累某一种代谢产物,而这个浓度又常常超过细胞正常生长和代谢所需的范围。因此要达到超量积累这种产物,提高生产效率,必须打破微生物原有的代谢调控系统,在适当的条件下,让微生物建立新的代谢方式,高浓度的积累人们所期望的产物[1]。 1 我国微生物代谢的研究现状 1.1 利用微生物代谢生产酶 工业上,曾由植物、动物和微生物生产酶。微生物的酶可以用发酵技术大量生产,是其最大的优点。而且与植物或动物相比,改进微生物的生产能力也方便得多。微生物的酶主要应用于食品及其有关工业中。酶的生产是受到微生物本身严格控制。为改进酶的生产能力可以改变这些控制,如在培养基中加入诱导物和采用菌株的诱变和筛选技术,以消除反馈阻遏作用。 1.2 利用微生物代谢产生的代谢产物生产目的物 在微生物对数生长期中,所产生的产物,主要是供给细胞生长的物质,入氨基酸、核苷酸、蛋白质、核酸、脂类和碳水化合物等。这些产物称为初级代谢产物。利用发酵生产的许多初级代谢产物,具有重大的经济意义,我国现已可以根据微生物代谢调控的理论,通过改变发酵工艺条件如pH、温度、通气量、培养基组成和微生物遗传特性等,达到改变菌体代谢平衡,过量生产所需要产物的目的。 1.3 利用微生物代谢理论发展产生了代谢工程 代谢工程是指利用基因工程技术,定向的对细胞代谢途径进行修饰、改造,以改变微生物的代谢特征,并于微生物基因调控、代谢调控及生化工程相结合,构建新的代谢途径,生产新的代谢产物的工程技术领域。 1.4 改变微生物代谢途径生产目的物 改变代谢途径是指改变分支代谢的流向,阻断其他代谢产物的合成,以达到提高目的产物的目的。改变代谢途径有各种方法,如加速限速反应,改变分支代谢途径流向、构建代谢旁路、改变能量代谢途径等不同方法[1]。 1.5 利用微生物代谢进行发酵 数千年来由于科学技术进步缓慢,各种微生物工业也未能充分发展。直到20世纪中期才建立了一系列新的微生物工业。近几年来,由于微生物代谢工程的应用,发酵工业开始进入新的发展时期。发酵产品增长快、质量明显提高,在国民经济中起重要作用。 1.6 微生物代谢在环境方面的应用 微生物降解是环境中去除污染物的主要途径。深人了解污染物在微生物内的代谢途径,将有助于人们优化生物降解的条件,从而实现快速的生物修复。这些代谢中间体大都通过萃取、分析方法进行逐个研究,并借助专家经验拟合出代谢途径,其动力学过程亦很少触及。代谢组学方法的采用有可能改变这一现状[2]。 1.7 利用微生物代谢进行赖氨酸的生产 在许多微生物中,可用天冬氨酸作原料,通过分支代谢途径合成出赖氨酸、苏氨酸和甲硫氨酸。赖氨酸在人类和动物营养上是一种十分重要的必须氨基酸,因此,在食品、医药和畜牧业上需求量很大。但在代谢过程中,一方面由于赖氨酸对天冬氨酸激酶有反馈抑制作用,另一方面,由于天冬氨酸除用于合成赖氨酸外,还要作为合成甲硫氨酸和苏氨酸的原料,因此,在正常细胞内,就难以累积较高浓度的赖氨酸。 为了解除正常的代谢调节以获得赖氨酸的高产菌株,工业上选育了谷氨酸棒杆菌的高丝氨酸缺陷型菌株作为赖氨酸的发酵菌种。由于它不能合成高丝氨酸脱氢酶,故不能合成高丝氨酸,也不能产生苏氨酸和甲硫氨酸,在补给适量高丝氨酸的条件下,可在含较高糖浓度和铵盐的培养基上,产生大量的赖氨酸[3]。 1.8 微生物代谢与分子生物学方法的结合 随着遗传学、分子生物学等方法的不断发展,人们越来越多地将这些方法运用到微生物的研究工作中。一些野生菌的合成能力或分泌能力有限,目前可通过人工诱变或构建高效的基因工程菌株等方法对其进行改造以扩大应用范围此外,现在许多细菌合成拮抗物质的基因已被克隆测序,为使植物获得微生物所具有的特殊功能,一种可能的方法是通过基因工程将目的基因导入植物体内,使植物直接表达活性物质[4]。 2 展望 2.1 微生物代谢在医药行业的展望 微生物在代谢过程中可分泌蛋白酶、纤维素酶、半纤维素酶、果胶酶、淀粉酶等几十种胞外酶进入培养基,这些酶有的可以将药物成分分解转化,形成新的化合物,有的可水解植物细胞壁的纤维素、半纤维素、果胶质等,使细胞破裂,利于有效成分溶出。特别是采用一些酶作用于药用植物材料,使细胞壁及细胞间质中的纤维素、半纤维素等物质降解,使细胞破裂,细胞间隙增加,减小细胞壁、细胞间物质传递屏障、对有效成分从胞内向胞外扩散的阻力减少,可促进有效成分的吸收提高。 2.2 微生物代谢在生理生化、微生物遗传育种方面的展望 随着分子生物学理论与技术的飞速发展,尤其是基因组和后基因组时代的到来,传统上的生理学与遗传学的交叉融合越来越多,许多
常见微生物检验项目与临床意义
微生物检验项目与临床意义 卫生部规定接受抗菌药物治疗的住院患者微生物检验样本送检率不低于30%(卫办医政发〔2011〕56号)。特介绍微生物检验项目,方便统计与分析点评。细菌检验为感染性疾病诊断和治疗提供依据:目标性治疗:提高微生物的送检率与检出率,有利于诊断与使用抗菌药。 经验治疗:根据本地区病原菌类型时间、区域、耐药谱等使用抗菌药。 调整治疗方案针对性用药:获得准确病原菌和细菌药敏结果。 细菌培养的临床意义 细菌培养与其它检验项目不同,由于其样本采集易受杂菌的干扰和培养条件的限制,因而造成检测结果有时与临床不完全一致,故在分析细菌培养报告时应明白:细菌培养阴性不代表无细菌感染、细菌培养阳性不代表该菌一定是病原菌,应结合患者具体情况而定。 1、血液和骨髓培养 目前血液培养仍然是菌血症和败血症的细菌学检验的基本方法,并且广泛地应用于伤寒、副伤寒及其它细菌引起的败血症的诊断。菌血症系病原菌一时性或间断地由局部进入血流,但并不在血中繁殖,无明显血液感染临床征象。常可发生在病灶感染或牙齿感染,尤以拔牙、扁桃体切除及脊髓炎手术后等多见。败血症是指病原菌侵入血流,并在其中大量生长繁殖,造成身体的严重损害,引起显著的全身症状(如不规则高热与全身中毒等症状),它多继发于组织器官感染,尤其是当机体免疫功能低下、广谱抗生素和激素的应用及烧伤等。 单次的血培养结果,对临床无鉴别指导意义,应多次多部位采集血液进行培养,才可判定检出菌究竟是病原菌还是污染菌。 抽血时应特别注意皮肤消毒和培养瓶口的灭菌。 2、脑脊液培养 正常人的脑脊液是无菌的,故在脑脊液中检出细菌(排除操作中的污染)应视为病原菌。引起脑膜炎的细菌种类不同,化脓性脑膜炎病原菌多为脑膜炎奈瑟菌,除此之外尚有肺炎链球菌、流感嗜血杆菌、金黄色葡萄球菌、大肠埃希菌、铜绿假单胞菌等。 3、尿液培养 尿液细菌培养对于膀胱和肾脏感染的及早发现和病原学诊断很有价值,对于尿道、前列腺以及内外生殖器的炎症也有一定价值。尿液中出现细菌通常称为菌尿症,如果尿液本身澄清但培养出细菌,一般为标本采集时未彻底消毒尿道口引起。如果细菌培养阳性同时伴有脓尿出现,则提示有尿路感染的可能(需指出轻度感染时可无脓尿出现)。泌尿系感染常见菌为大肠埃希菌、葡萄球菌、链球菌、变形杆菌等,除结核分枝支杆菌外,这些细菌又是尿道口常驻菌,极易引起标本留样污染,应注意鉴别。某些真菌疾病,如曲菌病在播散时也可造成肾脏感染,且尿中也能检查到。 尿液的细菌学培养具有重要的诊断意义,但培养阴性并不能否定泌尿道感染,一般应注意病程、采样的时机、方法以及其它原因的影响。 尿道口的消毒是中段尿细菌培养的关键步骤,是引起假阳性的最主要因素,故尿液细菌培养阳性要注意排除留样污染,如果检出菌的数量大于10万CFU/ml,则基本可认为是病原菌。 4、痰液和咽拭子标本的培养 痰液及支气管分泌物的细菌学检验对于呼吸道疾病的诊断、治疗具有一定的意义。对无法咳痰的患者,用咳嗽后的咽拭子做培养检查,仍是发现致病菌的主要依据。
微生物生理学实验指导圈起来的不做
微生物生理学实验 ―――铜绿假单胞菌fabI基因的克隆及功能鉴定 目录 实验〇铜绿假单胞菌fabI基因鉴定的原理 实验一细菌总DNA的提取与检测 实验二铜绿假单胞菌fabI基因PCR扩增 实验三铜绿假单胞菌pafabI基因表达载体构建 实验四质粒的提取与检测 实验五大肠杆菌感受态细胞的制备及DNA转化 实验六遗传互补大肠杆菌fabI温度敏感突变菌株 实验七铜绿假单胞菌烯酯酰ACP还原酶的原核表达 实验八铜绿假单胞菌烯酯酰ACP还原酶纯化 实验九体外鉴定烯酯酰ACP还原酶的酶活性 实验十构建铜绿假单胞菌fabI突变菌株 实验十一菌落PCR筛选重组子 实验十二铜绿假单胞菌脂肪酸合成分析 实验报告
实验〇铜绿假单胞菌fabI基因鉴定的原理脂肪酸的生物合成是一种细胞生物体重要的基础代谢。生物体脂肪酸合成系统(FAS)为磷脂和类脂等生物质膜组分提供重要的前体物质,同时生物体内一些重要的化合物如硫辛酸、生物素以及细菌群体感应的信号分子等也都需要以脂肪酸合成代谢中的中间产物为前体物质合成。 在不同生物体内,脂肪酸的生物合成过程的基本原理是相似的,但是催化反应的蛋白序列和结构则有着很大的差异。根据合成酶系统的这种差异,人们将脂肪酸生物合成系统分为I型脂肪酸合成酶系(FASI)和II型脂肪酸合成酶系(FAS II)(Cronan, 2006)。I型脂肪酸合成酶系主要分布在哺乳动物和真菌等生物中,该种脂肪酸合成由一个(或两个)分子量较大的多功能酶蛋白催化。这种多功能酶蛋白有多个不同功能的结构域,脂肪酸延伸循环中的各个反应分别在酶蛋白的不同结构域进行(Heath et al, 2002a; White et al, 2005)。II型脂肪酸合成酶系主要分布在细菌,植物和原生动物中,该类生物的脂肪酸合成由一组独立存在的酶催化完成,该系统中各个独立的酶蛋白分别行使一种功能(Cronan, 2006; White et al, 2005)。细菌脂肪酸的合成过程中,酰基链通过硫脂键连接在酰基载体蛋白(ACP)上。ACP是一种小分子量蛋白,一般由70-80个氨基酸残基组成,是FAS II的核心成员之一。 以大肠杆菌为例,说明脂肪酸合成的路径。 1. 脂肪酸合成的起始:脂肪酸合成的起始反应是以细菌体内的乙酰辅酶A 合成起初的四碳的短链脂肪酸的过程,该四碳的短链脂肪酸(acyl-CoA)及CO 2 产物是酰基链延长反应的基础(Revill et al, 2001; White et al, 2005)。此过程通过羧化、转移、缩合三步来完成:第一步,羧化:乙酰CoA在乙酰辅酶A 羧化酶(ACC)作用下转化为丙二酸单酰辅酶A(Mal-CoA)。ACC有4个独立的蛋白构成(AccA,AccB,AccC,AccD),其中AccB的行使功能需要生物素作为共价结合的辅因子(Cronan et al, 2002)。第二步,转移:Mal-CoA在丙二酰转酰基酶(FabD)的作用下将丙二酸单酰基团转移至ACP上,形成Mal –ACP(Jackowski et al, 1987)。第三步,缩合:在β酮脂酰ACP合成酶III(KAS III,FabH)的作用下将乙酰CoA所携带的酰基链合缩合到Mal-ACP上,生成β酮丁酰ACP,起始新的酰基链的生成(Revill et al, 2001)。
微生物代谢产物生产食品综述
微生物代谢产物生产食品综述 摘要:微生物代谢产物的种类很多,已知的有37个大类,其中16类属于药物,如氨基酸、核并酸、蛋白质、核酸、糖类、抗生素、生物碱、细菌毒素等。利用微生物的代谢产物可以生产十分丰富的食品。本文介绍了发酵生产食品的一般过程,并以谷氨酸为例详细说明。 关键词:微生物代谢产物食品谷氨酸发酵 正文: 微生物代谢产物的种类很多,已知的有37个大类,其中16类属于药物。在菌体对数生长期所产生的产物,如氨基酸、核并酸、蛋白质、核酸、糖类等,是菌体生长繁殖所必需的。这些产物叫做初级代谢产物,许多初级代谢产物在经济上具有相当的重要性,分别形成了各种不同的发酵工业。在菌体生长静止期,某些菌体能合成一些具有特定功能的产物,如抗生素、生物碱、细菌毒素等。这些产物与菌体生长繁殖无明显关系,叫做次级代谢产物。次级代谢产物多为低分子量化合物,但其化学结构类型多种多样,据不完全统计多达47类,其中抗生素的结构类型,按相似性来分,也有14类。由于抗生素不仅具有广泛的抗菌作用,而且还有抗病毒、抗癌和其他生理活性,因而得到了大力发展,已成为发酵工业的重要支柱。 利用微生物的代谢产物可以生产十分丰富的食品。以下举例说明: 1、食醋:食醋是人们日常生活所必需的调味品,也是最古老的利用微生物生产的食品之一。食醋生产是利用醋酸菌在充分供氧的条件下将乙醇氧化为醋酸。能用于食醋生产的醋酸菌有纹膜醋酸菌、许氏醋酸菌、恶臭醋酸菌和巴氏醋酸菌等。不同原料还需加入不同的微生物。以淀粉为原料时加入霉菌和酵母菌,糖类为原料时加入酵母菌。获得风味迥异的食醋品种。 2 .酒类:酒类的发酵生产主要是利用酵母菌在厌氧条件下将葡萄糖发酵为酒精的过程。不同酒类的发酵工艺不同:不同的酒类酿造所选用的酵母菌不同。所选用的原料、水质、甚至环境都会影响酒类的品质和风味。纯净的矿泉水往往较河水和自来水好。有人发现,贵州茅台酒之所以具有其独特的芬芳风味,与其酿酒厂环境中存在的微生物区系有关。 3 、发酵生产乳制品:利用乳酸细菌进行发酵,使成为具有独特风味的食品很多。如酸制奶油、干酪、酸牛乳、嗜酸菌乳(活性乳)、马奶酒、面包格瓦斯以及酸泡菜、乳黄瓜等等。这些乳制品不仅具有良好而独特的风味,而且由于易于吸收而提高了其营养价值。发酵乳制品的主要乳酸菌有干酪乳杆菌、保加利亚乳杆菌、嗜酸乳杆菌、植物乳杆菌、瑞士乳杆菌、乳酸乳杆菌、乳链球菌、乳脂链球菌、嗜热链球菌、噬柠檬酸链球菌、副柠檬酸链球菌等许多种。嗜柠檬酸链球菌还可以把柠檬酸代谢为具有香味的丁二酮等,使乳制品具有芳香味。 4 、发酵生产酱油:酱油是包括霉菌、酵母菌和细菌等多种微生物参与原料物质转化的混合作用的结果。对发酵速度、成品色泽、味道鲜美程度影响最大的是米曲霉和酱油曲霉,而影响其风味的是酵母菌和乳酸菌。米曲霉含有丰富的蛋白酶、淀粉酶、谷氨酸胺酶和果胶酶、半纤维素酶、酯酶等。涉及酱油发酵的酵母菌有 7 个属的 23 个种,其中影响最大的是鲁氏酵母,易变圆酵母等。 5 、腐乳的发酵生产:腐乳是大豆制品经多种微生物及其产生的酶,将蛋白质分解为胨、多肽和氨基酸类物质以及一些有机酸,有机醇和酯类而制成的具有特殊色香味的豆制品。涉及的微生物主要是毛霉中的腐乳毛霉、鲁氏毛霉、五通
赵立平和他的肠道微生物
赵立平:我和我的肠道菌群 1987 年,赵立平跟刘英结婚。两年不到,他们有了一个女儿,赵立平还完成了他的博士学位。新的压力加上美食——藉刘英做得一手好菜——我们的微生物学家长胖了。到 1990 年,体重从60 公斤增至 80 公斤。后来,在美国康乃尔大学读博士后期间,他又长胖了 10 公斤。1995 年腰围量110 厘米,整个人的健康状况也极为糟糕。赵立平认为,调节肠道菌群是他减肥成功的关键。 进入平台期的科研 转机2004 年,他读到一篇论文。论文的主要作者戈登,是美国华盛顿大学医学院的微生物学家。用实验表明,肥胖症和小鼠肠道中的菌群存有关联。 2006 年,他开始了一种饮食疗法,食谱中包括山药和苦瓜,同时对自己的体重以及肠道中的菌群进行监测。山药和苦瓜里面含有益生元,可以被肠道细菌发酵利用,据说有调节人体肠道菌群生长的功效。再加上以粗粮为主的饮食,在两年内减掉了 20 公斤。他的血压、心率和胆固醇水平也都降了下来。 抗炎细菌Faecalibacterium prausnitzii的数量大幅增加,从最开始根本检测不到,到后来增加为他肠道细菌总量的 14.5%。这些变化使赵立平决定,集中研究微生物在他身体状况转变中所发挥的影响。从小鼠身上做起的实验,赵立平把它扩展到了人的身上。 寻找新的启发 有一天,一位兽医学的同事问他要一些芽孢杆菌的菌株,说是这种细菌能缓解猪和鸡的腹泻。赵立平意识到,自己研究的菌株里面,很可能就有能抗植物感染,甚至抗人类感染功效的。 20 世纪 90 年代,赵立平涉足猪的微生物研究,试图探究用菌株来控制猪身上的感染这一设想,但无法获得资金。在此期间,他家人的健康状况每况愈下。本来就偏胖的父亲胆固醇水平激增,还得遭遇了两次中风。赵立平的两个弟弟也都成了大胖子。几年后,赵立平看到了戈登的论文,对他来说,这是“肠道菌群可以调节宿主的基因的首个证据”。于是,赵立平拿自己当小白鼠,试图找出体重增加可能跟哪些微生物有关。要从生活在人体肠道内上百种不同的微生物中,找出让体重增加的那一种,确实是个棘手的大难。 低热量饮食结合剧烈运动的减肥方法,在他看来完全说不通。“从营养上讲,你的身体是在压力状态下的,”赵立平说,“然后你再加上生理上的压力。也许这样你是能减肥,但同时也减掉了你的健康。” 腰围变小之后,他开始在动物身上进行实验,试图筛选出与肥胖有关的细菌。今年 4 月,赵立平《国际微生物生态学会会刊》上发表了一项研究,他们将小鼠从正常饮食转换到高脂肪饮食,然后再换回到正常饮食上来,期间每两周监测一
常见微生物的代谢方式
常见微生物的代谢方式 马丽甘肃省临夏回民中学(731100) 微生物种类繁多,代谢方式多样,本文将一些常见微生物的代谢方式归纳如下。所涉及生物中,除特别标注外,其它均为原核生物。 1、光能自养需氧型 这类微生物以光为能源,以CO2为主要碳源,适合生存于有氧环境,如:蓝藻、衣藻(原生生物)。 2、化能自养需氧型 这类微生物以无机化学能为能源,以CO2为主要碳源,适合生存于有氧环境,如:铁细菌、无色硫细菌、硝化细菌。 3、光能自养厌氧型 这类微生物如:绿硫菌,以光为能源,以CO2为主要主要碳源;有光合色素,进行光合作用获取生长所需要的能量;以无机物如H2、H2S、S等作为供氢体或电子供体,使CO2还原为细胞物质。适合生存于无氧环境。 4、化能异养需氧型 这类微生物的能源和碳源均来自于有机物,适合生存于有氧环境,真菌和绝大多数的细菌都是这一类型,常见的有:霉菌(真核生物)、草履虫及变形虫(原生生物)、放线菌、根瘤菌、圆褐固氮菌、肺炎双球菌、结核杆菌、霍乱弧菌、炭疽杆菌、麻风杆菌、黄色短杆菌、土壤农杆菌、枯草芽孢杆菌、苏云金芽孢杆菌、谷氨酸棒状杆菌等。 5、化能异养厌氧型 这类微生物的能源和碳源也是均来自于有机物,但是只有在缺氧的条件下才能很好的生长,如:乳酸菌、甲烷杆菌、反硝化细菌、破伤风杆菌、幽门螺旋杆菌。 6、化能异养兼性厌氧型 这类微生物的能源和碳源也是均来自于有机物,在有氧和无氧的条件下均能生长,如:大肠杆菌、酵母菌(真核生物)、金黄色葡萄糖球菌、支原体、酿脓链球菌。 7、兼性营养需氧型 这类微生物比较少见,如:裸藻,又叫眼虫(原生生物),适合生存于有氧环境,它在含有有机物的水中,能够靠细胞膜吸取水里的有机物“食物”,过着动物式的化能异养生活。但是同时,眼虫的细胞中具有含叶绿素的叶绿体,在无有机物的情况下,能够自己制造营养物质进行光合作用。因此兼有光能自养和化能异养的代谢方式。 8、兼性营养兼性厌氧型 这类微生物也是比较少见,如:红螺菌,它的同化方式是兼性营养型,以光为能源,以二氧化碳为主要碳源,以水或其他无机物作为供氢体,进行光合作用,还原CO2合成有机物。属于光能自养;或者以光为能源,以有机物为主要碳源,并且以有机物作为供氢体进行光合作用,同化有机物形成自身物质,属于光能异养。而它的异化方式也是兼性的,在湖泊、池塘的淤泥中进行厌氧呼吸;而在废水处理体系中却是需氧的。
第五章微生物代谢 答案
第五章微生物能量代谢 一、选择题(只选一项,将选项的的字母填在括号内) 1.下列哪种微生物能分解纤维素?( B ) A金黄色葡萄球菌B青霉C大肠杆菌D枯草杆菌 2.下列哪种产能方式其氧化基质、最终电子受体及最终产物都是有机物?( A ) A发酵B有氧呼吸C无氧呼吸D光合磷酸化 3.硝化细菌的产能方式是( D ) A发酵B有氧呼吸C无氧呼吸D无机物氧化 4.微生物在发酵过程中电子的最终受体是(A) A有机物B有机氧化物C无机氧化物D.分子氧 5.乳酸发酵过程中电子最终受体是( B ) A乙醛B丙酮 C O2 D NO3ˉ 6.硝酸盐还原菌在厌氧条件下同时又有硝酸盐存在时,其产能的主要方式是( C ) A发酵B有氧呼吸C无氧呼吸D无机物氧化 7.下列哪些不是培养固氮菌所需要的条件?( A ) A培养基中含有丰富的氮源B厌氧条件C提供A TP D提供[H] 8.目前认为具有固氮作用的微生物都是( D ) A真菌B蓝细菌C厌氧菌D原核生物 9.代谢中如发生还原反应时,( C )。 A从底物分子丢失电子B通常获得大量的能量 C 电子加到底物分子上D底物分子被氧化 10.当进行糖酵解化学反应时,( D )。 (a)糖类转变为蛋白质 (b)酶不起作用 (c)从二氧化碳分子产生糖类分子 (d)从一个单个葡萄糖分子产生两个丙酮酸分子 11.微生物中从糖酵解途径获得( A )ATP分子。 (a)2个 (b)4个 (c)36个 (d)38个 12.下面的叙述( A )可应用于发酵。 (a)在无氧条件下发生发酵 (b)发酵过程发生时需要DNA (c)发酵的一个产物是淀粉分子 (d)发酵可在大多数微生物细胞中发生 13.进入三羧酸循环进一步代谢的化学底物是( C )。 (a)乙醇 (b)丙酮酸 (c)乙酰CoA (d)三磷酸腺苷 14.下面所有特征适合于三羧酸循环,除了( D )之外。 分子以废物释放 (b)循环时形成柠檬酸 (a)C0 2 (c)所有的反应都要酶催化 (d)反应导致葡苟糖合成 15.电子传递链中( A )。 (a)氧用作末端受体 (b)细胞色素分子不参加电子转移 (c)转移的一个可能结果是发酵 (d)电子转移的电子来源是NADH 16.化学渗透假说解释( C )。 (a)氨基酸转变为糖类分子 (b)糖酵解过程淀粉分子分解为葡萄糖分子 (c)捕获的能量在ATP分子中 (d)用光作为能源合成葡萄糖分子 17.当一个NADH分子被代谢和它的电子通过电子传递链传递时,( C )。 (a)形成六个氨基酸分子 (b)产生一个单个葡萄糖分子 (c)合成三个ATP分子 (d)形成一个甘油三酯和两个甘油二酯 18.己糖单磷酸支路和ED途径是进行( C )替换的一个机制。
(完整版)微生物学第七章生长与控制
第十六授课单元 一、教学目的 此章为本课程的重点内容之一,使学生掌握微生物生长发育的规律及生长条件的控制,掌握生长的测定方法,学会同步培养和连续培养的方法,了解物理因素、化学因素对微生物生长发育的影响及实际应用,掌握消毒和灭菌的原理和方法等 本教学单元注重使学生了解微生物生长的测定:重点介绍单细胞微生物的典型生长曲线,并了解丝状真菌的生长曲线;介绍同步培养的方法(机械筛选法和环境条件控制法);恒浊连续培养和恒化连续培养的原理、控制方法和应用。 二、教学内容 第七章微生物的生长及其控制 第一节个体细胞生长概述 第二节微生物的群体生长 一、单细胞微生物的生长曲线 二、丝状真菌的生长曲线 三、同步培养 四、连续培养 第三节微生物生长的测定 一、计数法 二、质量法 三、生理指标法 三、教学重点、难点及处理方法 重点: 1. 单细胞微生物的典型生长曲线, 在介绍单细胞微生物的生长曲线之前让学生了解微生物生长测定的方法. 根据对于微生物生长的测定, 重点介绍单细胞微生物的典型生长曲线, 说明各个时期微生物生长的特点, 并结合实践说明微生物生长曲线对于生产有何指导意义. 2. 同步培养的方法(机械筛选法和环境条件控制法);恒浊连续培养和恒化连续培养的原理、控制方法和应用. 连续培养的原理来自于典型生长曲线, 使微生物保持一定比生长速率进行生长. 在一个恒定体积的培养物中, 通过不断地移出营养物质和以同样速率移走培养物的方法来得以实现. 难点: 1. 单细胞微生物的典型生长曲线对于单细胞微生物生长曲线中的指数期的三个重要参数例如: 繁殖代数, 代时和生长速率常数的意义及其相互关系及计算方法应说明清楚. 并将生长曲线各个时期对于实践的指导意义举例加以说明. 以加深对于生长曲线的理解. 分析微生物的生长曲线, 有重要的实际意义. 首先在扩大培养各级种子时就必须选择适宜的菌龄和接种量.其次为了获得大量菌体或代谢产物, 需经常设法延长细胞的对数生长阶段. 这就是连续培养的根据. 2. 恒浊连续培养和恒化连续培养的原理、控制方法和应用. 恒浊连续培养和恒化连续培养的原理比较复杂, 应用画图的方法加以说明, 主要通过多媒体, 为学生展示恒浊连续培养主要是通过不断调节流速使培养液浊度保持不变, 从而使微生物保持一定比生长速率进行生长. 而此生长速率一般是微生物生长曲线中的最高生长速率. 但是恒化连续培养中, 细菌的生长速率取决于限制性因子的浓度, 并低于最高生长速率. 营养物质浓度对微生物有影响, 一般认为营养物质适当时, 并不影响微生物的生长速率, 而低浓度时, 则会影响. 而且在一定范围内生长速率与营养浓度成正比关系. 恒化培养所用的培养基成分中, 要将一种必须营养物质控制在较低浓度, 以作为限制生长因子, 其它营养均可过量, 这样细胞的生长速率将