细胞因子的来源和功能
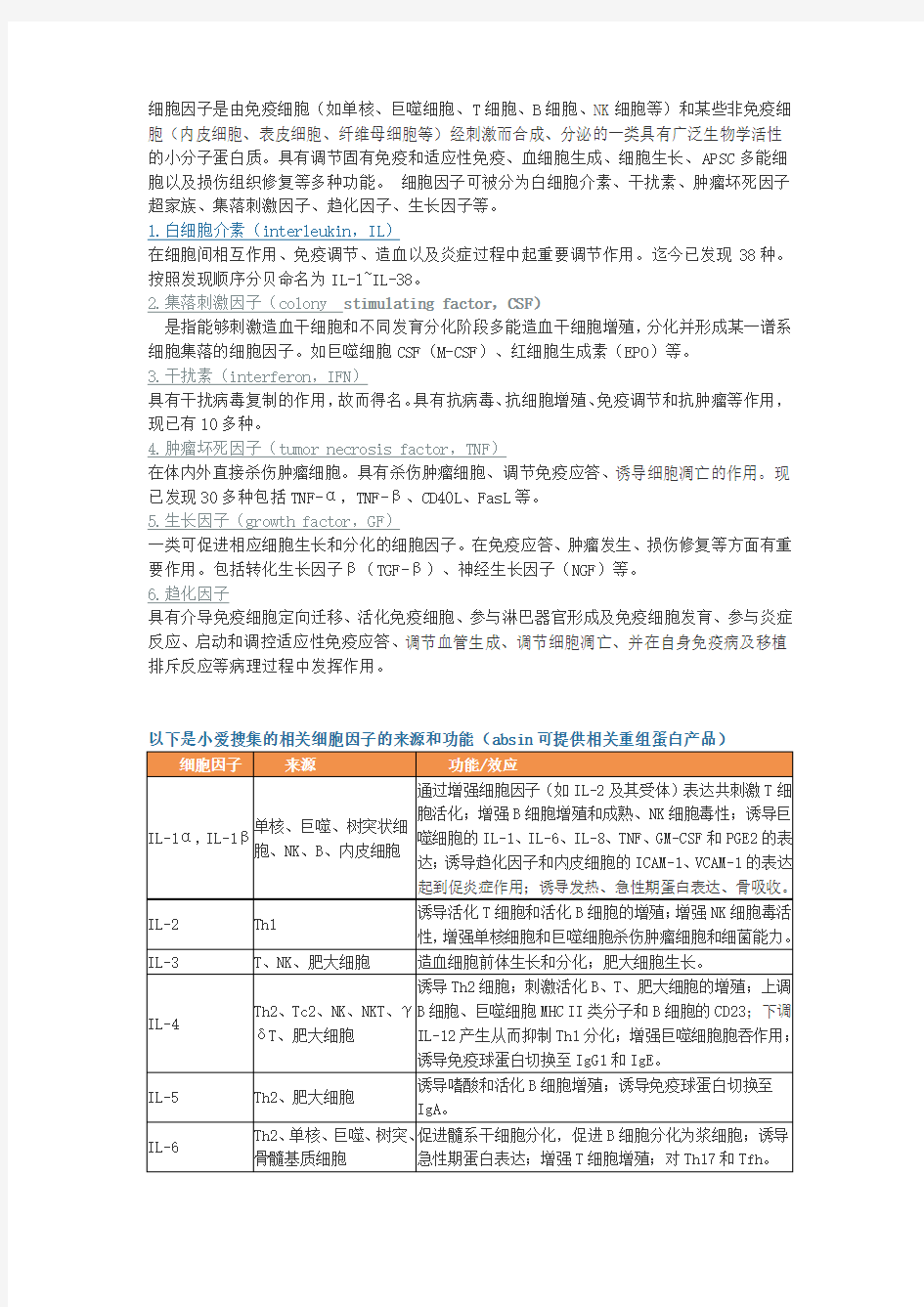
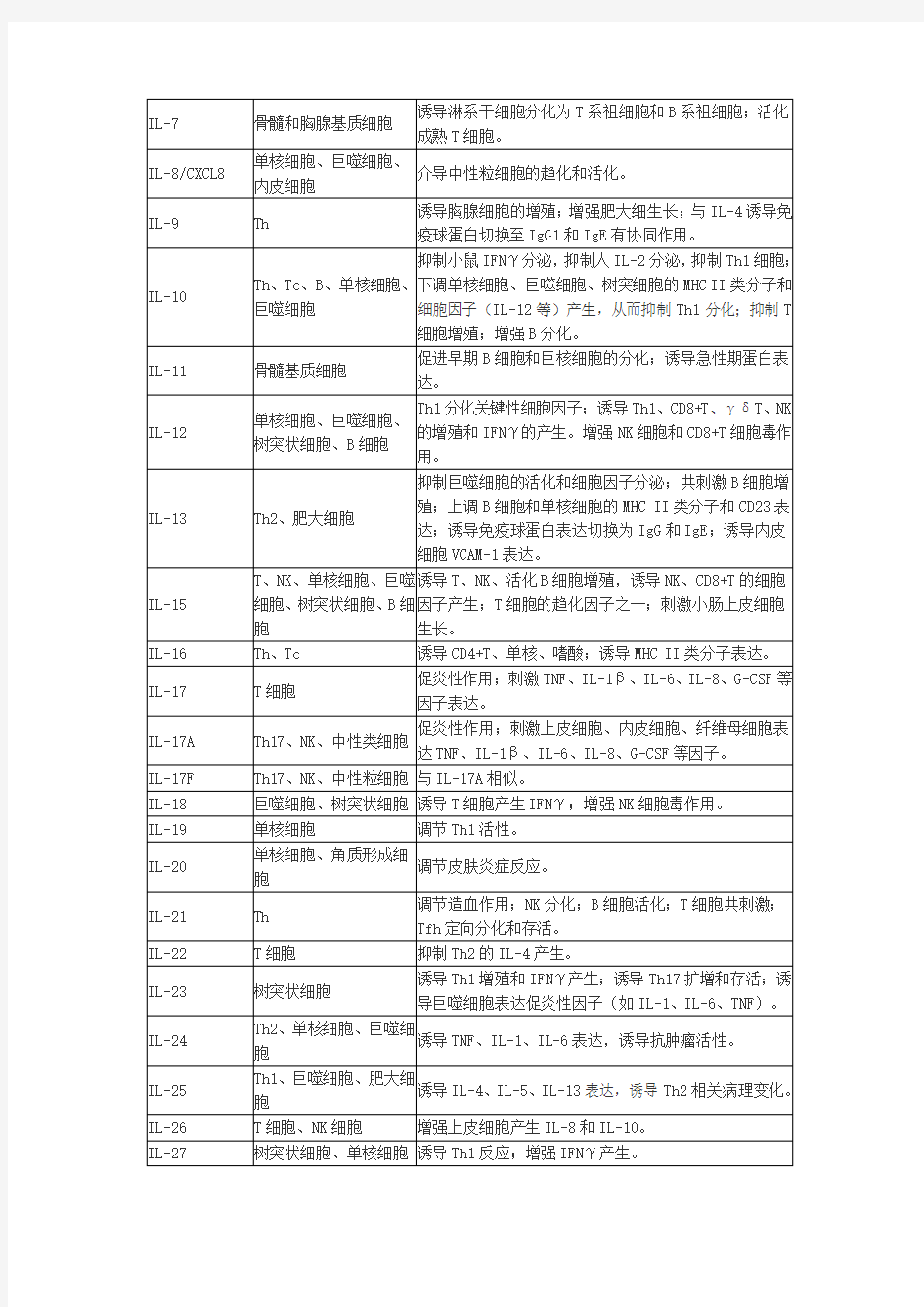
细胞因子详解
捋捋让人迷惑的细胞因子 细胞因子是一类调节蛋白或者糖蛋白,他们的分类现在还不是完全清楚。他们通过结合细胞表面的特定受体,激发细胞内信号通路起作用。 白细胞组成了免疫和炎症系统,大多数细胞因子作用于白细胞或者由白细胞表达,他们在免疫和炎症反应中起到重要的调节作用。实际上,一些免疫抑制和抗炎作用的药物就是通过调节这些细胞因子的表达起作用的。 细胞因子由特定的细胞表达并分泌到胞外,结合细胞表面的细胞因子受体后激活细胞内信号 传导通路 细胞因子分类 细胞因子最早在20世纪70年代中期被提出,它当时被认为是一种多肽因子,可以调控细胞分化和免疫系统。干扰素(IFNs)和白介素(ILs)是主要的多肽家族,在当时细胞因子主要指这两类家族。 起初细胞因子的分类主要是根据分泌该因子的细胞类型或者细胞因子初次被发现时的生物活性。然而这些分类方法现在看来都不够准确,无法满足后期的分类需求。最近,根据细胞
因子一级,二级和三级结构的分析,可以将大多数的细胞因子分为6大家族。因此,根据分类方式的不同,某些细胞因子会有多个名称。 表1:细胞因子根据结构分类结果 细胞因子家族成员 ‘β-Trefoil’ cytokines Fibroblast growth factors Interleukin-1 Chemokines Interleukin-8 Macrophage inflammatory proteins ‘Cysteine knot’ cytokines Nerve growth factor Transforming growth factors Platelet-derived growth factor EGF family Epidermal growth factor Transforming growth factor-αHaematopoietins Interleukins 2–7, -9, -13 Granulocyte colony stimulating factor Granulocyte-macrophage colony stimulating factor Leukaemia inhibitory factor Erythropoietin Ciliaryneurotrophic factor TNF family Tumour necrosis factor-α and –β
T细胞亚群的分类及功能
T细胞亚群的分类及功能 更新时间:2004-4-29 3:56:00 T细胞是不均一的群体,按其抗原识别受体,可将T细胞分为二大类。一类是TCRαβ、T细胞,另一类是TCRγδ细胞(表8-2)。 表8-2 TCRαβ+T细胞与TCRγδT细胞的特性 TCRαβT细胞TCRγδT细胞 分子结构二硫键相连的异二聚体分子二硫键相连的异二聚体分子 多样性多少 分布周围血60%~70% 周围血1%~10%表皮及肠粘膜上皮 表型CD4+CD8+(DP)60% CD4-CD8-(DN)35% CD2+100% CD5+>95% CD4-CD8-(DN) CD2+ CD5- 发育胸腺(发生晚) 胸腺(发生早)存在胸腺外途径 功能 识别与MHC分子结合的多肽复合 分子抗原 可能的作用 第一线防御细胞的原始受体 可识别MHC或MHC样分子识别由MHC 样分子呈递的抗原 TCRαβT细胞也是不均一的群体,根据其表型(phenotype)即其细胞表面的特征性分 子的不同,可将成熟T细胞分为二个亚类(subsets)即CD4+T细胞和CD8+细胞。 根据TCRαβT细胞的功能可将其分为二类。一类为调节性T细胞,可包括辅助性T细胞(helper T lymphocte,TH)和抑制性T细胞(suppressor T lymphocyte,Ts)。另一类为效应性T细胞(effector T cell),可包括杀伤性T细胞(eytolytie T cell,CTL,或TC)和迟发型超敏性T细胞(delayed type hypersensitivity T lymphoctye,TDTH)。 (一)TCRαβT细胞 二类T细胞表型分子均呈CD2+、CD3+阳性,但γδT细胞为CD4-、CD8-双阴性细胞(double negative cell,DN)或CD8+,而αβT细胞其表型为CD4+或CD8+单阳性细胞(single positive cell,SP)。
血细胞的种类和生理功能
红细胞的生理功能 红细胞的主要功能是运输O2和CO2,此外还在酸碱平衡中起一定的缓冲作用。这两项功能都是通过红细胞中的血红蛋白来实现的。如果红细胞破裂,血红蛋白释放出来,溶解于血浆中,即丧失上述功能。 白细胞的功能 白细胞是机体防御系统的一个重要组成部分。它通过吞噬和产生抗体等方式来抵御和消灭入侵的病原微生物。 1.吞噬作用吞噬作用是生物体最古老的,也是最基本的防卫机制之一。对于其要消灭的对象无特异性,在免疫学中称之为非特异性免疫作用。中性粒细胞和单核细胞的吞噬作用很强,嗜酸性粒细胞虽然游走性很强,但吞噬能力较弱。 白细胞可以通过毛细血管的内皮间隙,从血管内渗出,在组织间隙中游走。它们吞噬侵入的细菌、病毒、寄生虫等病原体和一些坏死的组织碎片。一般认为,白细胞能向异物处聚集,并将其吞噬,这是因为白细胞有趋化性。由于细菌体或死亡的细胞所产生的化学刺激,诱发白细胞向该处移动(图5-5)。组织发炎时产生一种活性多肽,也是白细胞游动的诱发物质之一。 中性粒细胞内的颗粒为溶酶体,内含多种水解酶,能消化其所摄取的病原体或其他异物。一般一个白细胞处理5~25个细菌后,本身也就死亡。死亡的白细胞集团和细菌分解产物构成脓液。 单核细胞由骨髓生成,在血液内仅生活3~4天,即进入肝、脾、肺和淋巴等组织转变为巨噬细胞。变为巨噬细胞后,体积加大,溶酶体增多,吞噬和消化能力也增强。但其吞噬对象主要为进入细胞内的致病物,如病毒、疟原虫和细菌等。巨噬细胞还参与激活淋巴细胞的特异免疫功能。此外,它还具有识别和杀伤肿瘤细胞,清除衰老与损伤细胞的作用。 2.特异性免疫功能淋巴细胞也称免疫细胞,在机体特异性免疫过程中起主要作用。所谓特异性免疫,就是淋巴细胞针对某一种特异性抗原,产生与之相对应的抗体或进行局部性细胞反应,以杀灭特异性抗原。血液中淋巴细胞按其发生和功能的差异,分为T淋巴细胞和B淋巴细胞两类。 (1)细胞免疫细胞免疫主要是由T细胞来实现的。这种细胞在血液中占淋巴细胞总数的80%~90%。T细胞受抗原刺激变成致敏细胞后,其免疫作用表现以下三个方面。直接接触并攻击具有特异抗原性的异物,如肿瘤细胞,异体移植细胞;分泌多种淋巴因子,破坏含有病原体的细胞或抑制病毒繁殖;B细胞与T 细胞起协同作用,互相加强,来杀灭病原微生物。 (2)体液免疫体疫免疫主要是通过B细胞来实现的。当此细胞受到抗原刺激变成具有免疫活性的浆细胞后,产生并分泌多种抗体,即免疫球蛋白,以针对不同的抗原。B细胞内有丰富的粗面内质网,蛋白质合成旺盛。抗体通过与相应
第三章-免疫细胞
第三章免疫细胞 Chapter 3 Immunocytes 第一部分教学内容和要求 一、目的要求 掌握:淋巴细胞的种类、T、B细胞膜表面分子及功能,T、B细胞的功能、抗原提呈细胞的种类和功能,NK细胞的功能;熟悉:淋巴细胞的分化发育,单核-巨噬细胞和NK细胞的表面受体;了解:TCR基因和重排。 二、教学内容 1.T淋巴细胞的分化发育、膜表面分子、亚群及其功能。 2.B淋巴细胞的分化发育、膜表面分子、亚群及其功能。 3.NK细胞的膜表面分子、功能;NK细胞识别和杀伤靶细胞的机制。 4.巨噬细胞、树突状细胞、B细胞和非专职性抗原提呈细胞的分布及其在免疫中所发挥的作用。 5.中性粒细胞、嗜酸粒细胞、嗜硷粒细胞、肥大细胞、红细胞和血小板等细胞在免疫应答中的作用。 第二部分测试题 一、选择题 (一)单项选择题(A型题) 1.可刺激B淋巴细胞增殖转化的刺激物 A.PWM B.PHA C.ConA D.MHC E.BCG 2.中性粒细胞在血循环中存活的时间 A.数小时 B.十几小时 C.数天 D.数周 E.数月 3.可刺激T细胞增殖的刺激物是 A.ConA B.MHC C.SPA D.AFP E.LPS 4.具有特异性杀伤功能的细胞 https://www.360docs.net/doc/ec808790.html,K 细胞 B.巨噬细胞 C.中性粒细胞 D.细胞毒性T细胞 E.NK细胞 5.淋巴细胞增殖试验可用来检测 A.细胞免疫功能 B.体液免疫功能 C.淋巴细胞数量 D.抗原提呈功能 E.补体功能 6.既具有抗原加工提呈作用又具有杀菌作用的细胞 A.树突状细胞 B.巨噬细胞 C.中性粒细胞 D.B细胞 E.T细胞 7.NK细胞表面的杀伤细胞抑制受体可识别 A.自身组织细胞表面的糖类配体复合物 B.肿瘤细胞表面的糖类配体 C.自身组织细胞表面的MHC-Ⅰ类分子 D.自身组织细胞表面的MHC-Ⅱ类分子 E.表达于感染细胞表面的病毒蛋白 8.具有SRBC受体的细胞是 A.T 细胞 B.B细胞 C.肥大细胞 D.NK 细胞 E.巨噬细胞 9.含有T细胞百分率最高的部位是 A.胸导管 B.胸腺 C.脾脏 D.外周血 E.扁桃体 10.参与非特异性免疫作用的细胞是 A.CD4+Th1细胞 B.CD4+Th2细胞 C.γδT细胞 D.αβT细胞 E.CD8+Tc细胞 11.B细胞识别抗原的表面分子是 A.C3 受体 B.mIg C.SRBC 受体 D.EB病毒受体 E.HIV受体
细胞受体类型特点及重要的细胞信号导途径
请归纳总结细胞受体类型、特点及重要的细胞信 号转导途径 受体是一类能够识别和选择性结合某种配体(信号分子)的大分子,大多数受体是蛋白质且多为糖蛋白,少数是糖脂,有的则是以上两者则是以上两者组成的复合物。受体是细胞膜上或细胞内能识别生物活性分子并与之结合的成分,它能把识别和接受的信号正确无误地放大并传递到细胞内部,进而引起生物学效应。 在细胞通讯中,由信号传导细胞送出的信号分子必须被靶细胞接收才能触发靶细胞的应答,接收信息的分子称为受体,此时的信号分子被称为配体。在细胞通讯中受体通常是指位于细胞膜表面或细胞内与信号分子结合的蛋白质。 一丶受体类型 根据靶细胞上受体存在的部位,可以将受体分为细胞内受体和细胞表面受体。细胞内受体存在于细胞质基质或核基质中,主要识别和结合小的脂溶性信号分子。细胞表面受体主要识别和结合亲水性信号分子。根据受体存在的标准,受体可大致分为三类:1.细胞膜受体:位于靶细胞膜上,如胆碱受体、肾上腺素受体、多巴胺受体、阿片受体等。 2.胞浆受体:位于靶细胞的胞浆内,如肾上腺皮质激素受体、性激素受体。 3.胞核受体:位于靶细胞的细胞核内,如甲状腺素受体。
另外也可根据受体的蛋白结构、信息转导过程、效应性质、受体位置等特点将受体分为四类: 1.离子通道偶联受体:如N-型乙酰胆碱受体含钠离子通道。 2.G蛋白偶联受体:M-乙酰胆碱受体、肾上腺素受体等。 3.酶联受体:如胰岛素受体,甾体激素受体、甲状腺激素受体等。 有些受体具有亚型,各种受体都有特定的分布部位核特定的功能,有些细胞也有多种受体。 二丶受体特点 1.受体与配体结合的特异性 特异性现为在同一细胞或不同类型的细胞中,同一配体可能有两种或两种以上的不同受体;同一配体与不同类型受体结合会产生不同的细胞反应,例如肾上腺素作用于皮肤粘膜血管上的α受体使血管平滑肌收缩,作用于支气管平滑肌上的β受体则使其舒张。 2.配体与受体结合的饱和性 受体可以被配体饱和。特别是胞浆受体,数量较少,少量激素就可以达到饱和结合。如在对甾体激素敏感的细胞中胞浆受体的数目最高每个细胞含量为10万个,雌激素受体,每个细胞中含量只有1000~50000个。故在一定浓度的激素作用下可以被饱和,而非特异性结合则不能被饱和。 3.功能上的有效性
最新T细胞亚群的分类及功能
T细胞亚群的分类及功能 1 2 更新时间:2004-4-29 3:56:00 3 T细胞是不均一的群体,按其抗原识别受体,可将T细胞分为二大类。4 一类是TCRαβ、T细胞,另一类是TCRγδ细胞(表8-2)。 5 表8-2 TCRαβ+T细胞与TCRγδT细胞的特性
6 TCRαβT细胞也是不均一的群体,根据其表型(phenotype)即其细胞 7 表面的特征性分子的不同,可将成熟T细胞分为二个亚类(subsets)即CD4+T 8 细胞和CD8+细胞。 9 根据TCRαβT细胞的功能可将其分为二类。一类为调节性T细胞,可 10 包括辅助性T细胞(helper T lymphocte,TH)和抑制性T细胞(suppressor T 11 lymphocyte,Ts)。另一类为效应性T细胞(effector T cell),可包括杀伤性 12 T细胞(eytolytie T cell,CTL,或TC)和迟发型超敏性T细胞(delayed type 13 hypersensitivity T lymphoctye,TDTH)。 14 (一)TCRαβT细胞 15 二类T细胞表型分子均呈CD2+、CD3+阳性,但γδT细胞为CD4-、CD8- 16 双阴性细胞(double negative cell,DN)或CD8+,而αβT细胞其表型为CD4+ 17 或CD8+单阳性细胞(single positive cell,SP)。 18 在末梢血主要为αβT细胞可占95%,而γδT细胞只占1%~10%。αβT 19 细胞为主要参予免疫应答的T细胞,而对γδT细胞功能不十分了解,可能是 20 具有原始受体的第一防线的防御细胞,与抗原感染有关。 21 (二)CD4+细胞 22 TCRαβTCD4+细胞(简称为CD4+细胞)的分子表型为CD2+、CD3+、CD4+、 23 CD8-。其TCR识别抗原是MHCⅡ类分子限制性。CD4+T细胞也是不均一的细胞群, 按其功能可包括二种T细胞,即辅助性T细胞(TH),和迟发型超敏性T细胞24 25 (TDTH)。前者为调节性T细胞,后者为效应性T细胞。
成纤维细胞生长因子及其与受体作用机制的研究进展
成纤维细胞生长因子及其与受体作用机制 的研究进展1 姜媛媛,任桂萍,王文飞,郝建权,李德山 东北农业大学生命科学学院生物制药教研室,哈尔滨(150030) E-mail:deshanli@https://www.360docs.net/doc/ec808790.html, 摘要:成纤维细胞生长因子(FGF)是一类多肽类物质,其中大多数成员可与肝素结合发挥作用。目前已知FGF至少包括23个因子,即FGF1~23。部分FGF家族成员N末端有大约3O个氨基酸残基组成的典型信号肽序列,可以分泌到细胞外。FGF家族成员是一类生理功能较广泛的生长因子,功能包括促进细胞有丝分裂、趋化与血管生成、促进中胚层和神经外胚层细胞的存活与生长等。本文根据最近的研究成果对 FGF因子及其受体研究进展做一综述,并主要对FGF因子特征及其研究趋势进行了探讨。 关键词:FGF,FGF受体,肝素 中图分类号:Q74 引言: 成纤维细胞生长因子最早是从脑和垂体的提取液中发现的,该物质是一种能促进成纤维细胞生长的多肽类活性物质,可以通过与细胞膜特异性受体结合对细胞生长进行调节。从70年代中期到目前已进行了大量广泛的研究,目前已知FGF至少包括23个因子,它们在一级氨基酸序列上有一定的同源性,并有类似的生物学功能,且广泛存在于体内多种组织中。FGF对中胚层和神经外胚层来源的细胞具有十分明显的促细胞分裂增殖作用,并且在机体内的胚胎发育、细胞生长分化、创伤组织愈合及肿瘤发生发展中起着十分重要的作用。 1. 成纤维细胞生长因子(FGF)家族 成纤维生长因子(Fibroblast growth factor, FGF)又被称为肝素亲和生长因子(Heparin binding growth factor, HBGF),是一类通过与细胞膜特异性受体结合发挥作用的多肽分子。现已知FGF家族至少包括23个成员,即FGF1~FGF23。FGF家族成员之间的氨基酸序列同源性约为25%~50%,其每个成员都有140个氨基酸的中轴,该中轴在不同的成员中有高度的同源性。结构分析表明,此中轴折叠成12条逆向平行的β链,它们又进一步形成圆柱状的结构。部分FGF家族成员N末端有大约3O个氨基酸残基组成的典型信号肽序列,使得它们可通过内质网一高尔基复合体的经典(即自分泌和旁分泌)途径被分泌到细胞外,但其中也有部分FGF则因本身缺乏信号肽结构,不能向外分泌,只能在细胞受损时释放[1]。多数FGF(如FGF3~8、10、15、17~19、21~23)的N末端具有典型的信号肽序列。而FGF16和FGF20虽然没有明确的信号肽序列却也能高效地分泌到细胞外[12]。FGF1 和FGF2也缺乏信号肽序列和正常的分泌途径,却也能出现在胞外基质,推测两者可能来自受伤的细胞,或者通过与内质网-高尔基体通路不同的细胞脱颗粒机制释放至胞外的。此外,FGF 11~14没有典型的信号肽序列,因此认为这些FGF是在胞内发挥作用[13]。由于FGF家族成员之间的氨基酸序列有25%~50%的同源性,分子结构有一定的共性,故FGF不同分子之间的生物学效应既有相似性,又有各自的特点[1-3]。 FGF1(aFGF)和FGF2(bFGF)是最先被发现,也是迄今为止研究最充分的两个成员,因其1本课题得到黑龙江省科技厅重点攻关项目(编号:2006G0461-00)的资助。
简述免疫系统的组成及主要功能
作业1 选择题 1—5EDBCC 6—10DDEDC 11—15ACCBD 16—20BCCDB 简答题 1.免疫系统具有哪些功能?这些功能正常或是失常表现出何种生物学效应。 1)免疫防御:指机体抵御外来抗原性异物入侵的一种保护功能。正常时可抵御病原微生物的感染和损害,即抗感染免疫。异常时如果防御功能过强出现超敏反应,免疫防御功能过低(免疫缺陷)会导致反复发生感染。2)免疫稳定:指维持体内环境相对稳定的生理机能。正常时可及时清除体内损伤、衰老、变性的细胞以及抗原-抗体复合物等抗原性异物,对自身成分耐受和保护。功能紊乱时会导致自身免疫疾病,失去了对自身抗原的耐受而对自身细胞发动攻击。3)免疫监视:指免疫系统及时识别、清除体内突变、畸变细胞和病毒感染细胞的一种生理保护功能。功能正常时可防止肿瘤产生,功能失调时可导致肿瘤发生,或病毒感染不能及时清除,造成病毒持续性感染。 2.简述免疫器官的组成和主要功能 主要分为中枢免疫器官,外周免疫器官。中枢免疫器官包括骨髓、胸腺。外周免疫器官包括淋巴结、脾脏、黏膜相关淋巴组织。骨髓的主要功能:1)是各类血细胞和免疫细胞发生的场所。2)骨髓是B细胞分化、成熟的场所。3)是再次免疫应答时产生抗体的主要场所。胸腺的主要功能:1)T细胞发育的主要场所。2)免疫调节作用。3)建立与维持自身免疫耐受。淋巴结的主要功能:1)T/B淋巴细胞居留的场所。2)发生免疫应答的场所。3)参与淋巴细胞再循环。脾脏的主要功能:1)T/B淋巴细胞定居的场所。2)对血液来源抗原产生免疫应答的主要场所。3)合成多种生物活性物质。4)过滤作用。黏膜相关淋巴组织主要作用:1)通过黏膜局部发生的适应性免疫应答,在消化道、呼吸道、和泌尿生殖道的免疫防御中发挥重要作用。2)黏膜局部产生的分泌型IgA,3)参与口服抗原街道的免疫耐受。 3.细胞因子的概念及分类 细胞因子是由免疫原、丝裂原或其他因子刺激多种细胞(主要是免疫细胞)合成、分泌的具有生物学活性的小分子蛋白质。根据其结构和功能可分为白细胞介素、干扰素、肿瘤坏死因子超家族、集落刺激因子、生长因子和趋化因子等多种类型。 4.简述免疫球蛋白的生物学功能 (1)与抗原发生特异性结合:主要由Ig的V区,特别是HVR的空间结构决定的。在体内表现为抗细菌、抗病毒、抗毒素等生理学效应;在体外可出现抗原抗体反应。(2)激活补体:IgG(IgG1、IgG2和IgG3)、IgM类抗体与抗原结合后,可经经典途径激活补体;聚合的IgA、IgG4可经旁路途径激活补体。(3)与细胞表面的Fc 受体结合:Ig经Fc段与各种细胞表面的Fc受体结合,发挥调理吞噬、粘附、ADCC及超敏反应作用。(4)穿过胎盘:
细胞因子及分类
细胞因子的基本概念与种类 2008-11-19 13:38 【大中小】 细胞因子(CK)是由活化免疫细胞和非免疫细胞(如某些基质细胞)合成分泌的能调节细胞生理功能、参与免疫应答和介导炎症反应等多种生物学效应的小分子多肽或糖蛋白,是不同于免疫球蛋白和补体的又一类免疫分子。 根据来源最初将活化淋巴细胞产生的细胞因子称为淋巴因子(LK),将单核-吞噬细胞产生的细胞因子称为单核因子(MK)。目前根据功能,可将细胞因子粗略分为以下6类: 1.白细胞介素 白细胞介素(IL)简称白介素,最初被定义为由白细胞产生,在白细胞间发挥作用的细胞因子。虽然后来发现它们的产生细胞和作用细胞并非局限于白细胞,但这一名称仍被沿用。 目前已报道的白细胞介素有18种,摘要列表如下: 2.干扰素(IFN) 干扰素是最先发现的细胞因子,因其具有干扰病毒感染和复制的能力故称为干扰素。根据来源和理化性质,可将干扰素分为α、β和γ三种类型。IFN-α/β主要由白细胞、成纤维细胞和病毒感染的组织细胞产生,称为Ⅰ型干扰素。IFN-γ主要由活化T细胞和NK细胞产 生,称为Ⅱ型干扰素。 丙型干扰素生物学性能比较详见下表: 3.肿瘤坏死因子(TNF) 肿瘤坏死因子是一类能引起肿瘤组织出血坏死的细胞因子。1975年Garwell等将卡介苗注射给荷瘤小鼠,两周后再注射脂多糖,结果在小鼠血清中发现一种能使肿瘤发生出血坏死的物质,称为肿瘤坏死因子。肿瘤坏死因子分为TNF-αTNF-β两种,前者主要由脂多糖/卡介苗活化的单核巨噬细胞产生,亦称恶病质素;后者主要由抗原/有丝分裂原激活的T细胞产生,又称淋巴毒素。TNF-α/β为同源三聚体分子,主要生物学作用如下: (1)对肿瘤细胞和病毒感染细胞有生长抑制和细胞毒作用; (2)激活巨噬细胞、NK细胞,增强吞噬杀伤功能,间接发挥抗感染、抗肿瘤作用; (3)增强T、B细胞对抗原和有丝分裂原的增生反应,促进MHC-Ⅰ类分子表达,增强 Tc细胞杀伤活性; (4)诱导血管内皮细胞表达粘附分子和分泌IL-1、IL-6、IL-8、CSF等细胞因子促进炎 症反应发生; (5)直接作用或刺激巨噬细胞释放IL-1间接作用下丘脑体温调节中枢引起发热; (6)引起代谢紊乱,重者出现恶病质。 4.集落刺激因子(CSF)
细胞因子的作用特点
细胞因子的作用特点 目前发现并正式命名的细胞因子有数十种,每种细胞均有其独特的、起主要作用的生物学活性。尽管种类繁多、产生细胞和作用细胞多样、生物学活性广泛、发挥作用的机制不同,但众多的细胞因子具有以下共同的特性: 1.天然细胞因子是由细胞产生的正常的静息或休止(resting)状态的细胞必须经过激活后才能合成和分泌细胞因子。通常是由抗原、丝裂原或其它刺激物激活免疫细胞和相关细胞,6~8小时后细胞培养上清中即可检测出细胞因子,于24~72小时期间细胞因子水平最高。但是有些细胞株不需外源刺激就可以自发地分泌某些细胞因子。 2.细胞因子的产生和作用具有多向性(pleiotropism)即单一刺激如抗原、丝裂原、病毒感染等可使同一种细胞分泌多种细胞因子,而一种细胞因子由多种不同类型的细胞产生可作用于多种不同类型的靶细胞。 3.细胞因子的合成和分泌过程是一种自我调控的过程通常情况下,细胞因子极少储存,即不以前体形式贮存在细胞内,而是经过适当刺激后迅速合成,一旦合面后便分泌至细胞外以发挥生物学作用,刺激消失后合成亦较快地停止并被迅速降解。 4.为低分子量的分泌型蛋白质常被糖基化。分子量大小不等,大多数为15~30kD,小者仅8~10kD,一般不超过80kD。 5.细胞因子需与靶细胞上的高亲和力受体特异结合后才发挥生物学效应。 6.生物学效应极强细胞因子在pM(10-12M)水平就能发挥显著的生物学效应。这与细胞因子与靶细胞表面特异性受体之间亲和力极高有关,其解离常数在10-12~10-10M之间。 7.单一细胞因子可具有多种生物学活性,但多种细胞因子也常具有某些相同或相似的生物学活性。 8.主要参与免疫反应和炎症反应影响反应的强度和持续时间的长短。涉及到感染免疫、肿瘤免疫、自身免疫、移植免疫等诸多方面。 9.以非特异性方式发挥生物学作用且不受MHC限制。
细胞生理学题库
第二章细胞生理学 第一节细胞膜的物质转运功能 掌握内容说出跨膜物质转运的几种主要方式。复述单纯扩散、易化扩散、 主动转运的概念。列举单纯扩散的物质种类。说出易化扩散的种类及其特征。列 举离子通道的控制类型。说出钠钾泵的工作原理,列举钠钾泵的意义。复述继发 性主动转运的概念,说出继发性主动转运的原理和特点。 熟悉内容描述物质入胞和出胞转运的过程,列举入胞和出胞转运的生理 现象,说出入胞作用的几种类型。 了解内容简单复习细胞膜的成分和结构(液态镶嵌模型),解释并列举细 胞膜的主要生理功能。讨论葡萄糖的跨上皮转运机制。讨论易化扩散的生理意义。 讨论入胞和出胞转运的生理意义。 (一)名称解释 液态镶嵌模型、单纯扩散、易化扩散、通道、载体、电压门控、受体门控、机械门控、饱和现象、主动转运、继发性主动转运、入胞作用、出胞作用、受体内化。 (二)思考题与讨论 1.葡萄糖的跨上皮转运机制和临床应用。 2.钠钾泵的生理意义。 3.团块物质转运的意义。 (三)选择题 A型题 【A1型题】单项选择题,每题有A、B、C、D、E五个备选答案,请从中选出一个最佳答案。 1.下列哪种脂质成分几乎全部分布在膜的靠近胞质的内层并与第二信使DG和IP3的产生有关 A.磷脂酰肌醇B.磷脂酰胆碱C.磷脂酰乙醇胺 D.磷脂酰丝氨酸E.糖脂 2.葡萄糖或氨基酸逆浓度梯度跨膜转运的方式属于 A.单纯扩散B.经载体易化扩散C.经通道易化扩散 D.原发性主动转运E.继发性主动转运 3.在膜蛋白质帮助下,某些胞外的蛋白质分子选择性地进入胞内的跨膜转运方式属于A.原发性主动转运B.继发性主动转运C.经载体易化扩散 D.受体介导入胞E.液相入胞
完整受体与细胞功能调控重点总结推荐文档
以下为胡雅儿教授讲解部分的重点内容,请参照PPT和课本宏观把握 第一章受体与受体后信号转导系统概念 P3放射性配基结合法是迄今为止研究受体数量和亲和力最主要的手段。 P3-P4受体的现代概念(包括受体的四大特点) P4孤儿受体 第二章受体和配基结合的基本规律 P7受体和配基结合的基本规律(包括四条) 「可逆性 可饱和性,对应着图2-2分析变化趋势 特异性 .配基受体结合反应和细胞效应的一致性 第三章受体的研究方法 P19人工造成受体分子的基因突变:定点突变、缺损突变、嵌合突变转基因动物和基因敲(剔)除动物的区别 第四章受体的分类 四级分类法:类,亚类,型,亚型 P24掌握图4-1和表4-1 体结构示意图) 第五至第九章就是针对上述的五大类受体展开的详细讲解 第五章与G蛋白偶联的膜受体及其受体后信号转导 P28膜受体的概念(分为三个部分) P28三种类型G蛋白把信号传递到效应器的途径 1.2.3.(本章第六、七、八节的总结概括, 只要求宏观掌握,即能把主线理清即可) P32受体亚型的结构功能关系,三条 P33鸟苷酸结合蛋白(G蛋白)的主要分类,依据a亚单位的氨基酸序列主要分四大类Gs, Gi, Gq, G12
P34 G蛋白的结构域示意图:分a亚单位和B Y亚单位以及各自结构域可能的作用 P34重点大题G蛋白的活化和失活机制 第六章酶联受体/有酶结构的单次跨膜受体/单次跨膜有激酶活性的 受体 主要信号分子是生长因子,主要分酪氨酸激酶受体和丝氨酸苏氨酸激酶受体 第七章无酶结构的单次跨膜受体/与胞浆内可溶性酪氨酸激酶偶联 的受体(不同称谓而已) P54 JAK-STA H路,结合图7-4,宏观把握 第八章离子通道受体 (配基与受体的结合或解离控制了通道的开关,通道的开关控制了一些离子的跨膜流量,进而改变细胞内离子浓度,达到调控细胞功能的目的) 中枢神经系统兴奋受体:N-乙酰胆碱受体 中枢神经系统抑制受体:GABA A受体 脊髓和脑干抑制受体:甘氨酸受体 外周神经元兴奋:5-HT3受体 每个亚单位有四个a螺旋组成,来回穿插细胞膜,最后的羧基端在膜外,近氨基端都有一对Cys形成二硫键,几个亚单位形成一个半胱氨酸环,所以称为Cys环类的离
第九章细胞信号转导习题及答案
细胞生物学章节习题-第九章 一、选择题 1、动物细胞内引起储存Ca2+释放的第二信使分子是( A )。 A. IP3 B. DAG C. cAMP D. cGMP 2、一氧化氮的受体是(B )。 A. G蛋白偶联受体 B. 鸟苷酸环化酶 C. 腺苷酸环化酶 D. 受体酪氨酸激酶 3、表皮生长因子(EGF)的穿膜信号转导是通过(A )实现的。 A. 活化酪氨酸激酶 B. 活化酪氨酸磷酸酶 C. cAMP调节途径 D. cGMP途径 4、有关cAMP信号通过,下列说法错误的是(B)。 A. 被激活的蛋白激酶A的催化亚基转为进入细胞核,使基因调控蛋白磷酸化 B. 结合GTP的α亚基具有活性,而βγ亚基复合物没有活性 C. βγ亚基复合物与游离的Gs的α亚基结合,可使Gs的α亚基失活 D. 这一通路的首要效应酶是腺苷酸环化酶,cAMP被环腺苷磷酸二酯酶消除 5、霍乱弧素引起急性腹泻是由于(A )。 A. G蛋白持续激活 B. G蛋白不能被激活 C. 受体封闭 D. 蛋白激酶PKC功能异常 E. 蛋白激酶PKA功能异常 6、G蛋白具有自我调节活性的功能,下列哪种说法可以解释G蛋白活性丧失的原因(A )。 A. α亚基的GTPase活性 B. 效应物的激活 C. 与受体结合 D. 亚基解离 7、胞内受体介导的信号转导途径对代谢调控的主要方式是下列哪种(A )? A. 特异基因的表达调节 B. 核糖体翻译速度的调节 C.蛋白降解的调节 D. 共价修饰调节 8、制备人类肝细胞匀浆液,然后通过离心技术分离细胞膜性成分和可溶性胞质。如在可溶胞质组分中加入肾上腺素,会发生下何种情况(D ) A. cAMP增加 B. 肾上腺素与其胞内受体结合 C. 腺苷环化酶的激活 D. cAMP浓度不变 9、1,4,5-三磷酸肌醇促进Ca2+从细胞那个部位释放进入细胞质(B ) A. 线粒体 B. 内质网 C. 质膜(从胞外到胞内) D. Ca2+-CaM复合体细胞 10、与视觉信号转导有关的第二信使分子是下列哪种成分(D )。 A. 花生四烯酸 B. cAMP C. Ca2+ D. cGMP 二、填空题 1、Ras蛋白在RTKs介导的信号通路中起着关键作用,具有GTPase活性,当结合GTP 时为活化状态,当结合GDP 时为失活状态。GAP增强Ras的失活。 2、介导细胞信号传递的受体分为细胞内受体、离子通道偶联受体、酶连接的受体和G蛋白偶联受体。 3、细胞分泌信号的作用方式分为:自分泌、内分泌、旁分泌;通过化学突出传递申请信号。 4、细胞表面受体丝氨酸/苏氨酸激酶是丝氨酸/苏氨酸蛋白激酶次跨膜蛋白受体,受体胞内区具有活性,它的主要配体是转化生长因子β家族成员 三、判断题 1、NO作为信号分子,它能使细胞内的cAMP水平升高。(x ) 2、Ca2+是细胞内广泛存在的信使,细胞质中游离的Ca2+浓度比胞外高。(x ) 3、细胞外信号都是通过细胞表面受体来进行跨膜信号传递的。(x ) 4、Ras蛋白被SOS激活后,可激活其下游的MEK激酶,再通过激活MEK激酶将Raf激酶
第二节 免疫细胞
第六章免疫系统 第二节免疫细胞 凡参与免疫应答或与免疫应答相关的细胞统称为免疫细胞。它们的种类繁多,功能各异,但相互作用,相互依存。根据它们在免疫应答中的功能及作用机理,可分为免疫活性细胞和免疫辅佐细胞两大类。此外还有一些其他细胞,如K 细胞、NK细胞、粒细胞、红细胞等,也参与了免疫应答中的某一特定环节。 一、免疫活性细胞 在淋巴细胞中,受抗原物质刺激后能增殖分化,并产生特异性免疫应答的细胞,称为免疫活性细胞,主要指T细胞和B细胞,在免疫应答过程中起核心作用。 (一)T、B细胞的来源与分布(图6-9)T、B细胞均来源于骨髓的多能干细胞,骨髓中的一部分多能干细胞首先分化为淋巴干细胞,并进一步分化为前T细胞和前B细胞。前T细胞进入胸腺发育为成熟的T细胞,并经血流分布到外周免疫器官的胸腺依赖区定居和增殖,并可经血液→组织→淋巴→血液再循环巡游全身各处。T细胞接受抗原刺激后活化、增殖、分化成为效应T细胞,发挥细胞免疫的功能。效应性T细胞是短寿的,一般存活4~6d,其中一小部分变为长寿的免疫记忆细胞,进入淋巴细胞再循环,它们可存活数月到数年。 前B细胞在哺乳动物的骨髓或鸟类的腔上囊分化为成熟的B细胞,成熟的B 细胞分布在外周免疫器官的非胸腺依赖区定居和增殖。B细胞接受抗原刺激后活化、增殖、分化为浆细胞,发挥体液免疫的功能。浆细胞一般只能存活2d。一部分B细胞成为免疫记忆细胞,参与淋巴细胞再循环,它们是长寿的B细胞,可存活100d以上。 (二)T、B细胞的表面标志 T细胞和B细胞在光学显微镜下均为小淋巴细胞,从形态上难于区分(图6-10)。在扫描电镜下多数T细胞表面光滑,有较小绒毛突起;而B细胞表面较为粗糙,有较多绒毛突起。但这不足以区别T细胞和B细胞。淋巴细胞表面存在着大量不同种类的蛋白质分子,这些表面分子又称为表面标志(surface marker)。T细胞和B细胞的表面标志包括表面受体和表面抗原,可用于鉴别T 细胞和B细胞及其亚群。 表面受体是淋巴细胞表面上能与相应配体(特异性抗原、绵羊红细胞、补体等)发生特异性结合的分子结构。表面抗原是指在淋巴细胞或其亚群细胞表面上能被特异性抗体(如单克隆抗体)所识别的表面分子。由于表面抗原是在淋巴细胞分化过程中产生的,故又称为分化抗原。不同的研究者和实验室已建立了多种单克隆抗体系统用以鉴定淋巴细胞表面抗原,出现了多种命名。为避免混淆,从1983年起,经国际会议商定以分化群(CD)统一命名淋巴细胞表面抗原或分子,如将单抗OKT3和单抗Leu4所识别的同一分化抗原命名为CD3等,至今已命名200余种CD抗原。 1.T细胞的表面标志 (1)T细胞抗原受体(TCR):T细胞表面具有识别和结合特异性抗原的分子
核受体概述和分类
核受体:概述和分类 摘要: 核受体超家族包括很多的转录因子,在多细胞生物体的发展和稳态方面发挥着重要的调节作用。核受体有一种特殊的功能即自身绑定到染色体上,这使得他们成为基因转录的重要起始者。此外,核受体具有在瞄准启动子和协调整个基因转录过程而依序招募各种转录因子和共调节因子的能力,证实了他们的生物学意义,并刺激了这一领域内深入的研究和高层次的科学兴趣。在这篇综述中,我们总结了当今对于作为基因表达的主要调节者核受体的结构和功能的认识。重点是介绍核受体介导的转录激活和抑制的分子机制,包括最近在这方面取得的进展。关键词:核受体、转录、配体、LBD、DBD、结构域、辅助因子、共调节因子。 核受体属于大的转录因子超家族,涉及如控制胚胎发育、器官生理、细胞分化、稳态等重要的生理功能[1,2]。除了正常的生理,核受体涉及到许多病理过程,如癌症、糖尿病、类风湿关节炎、哮喘或激素抵抗综合征[3-5]。在生物医学研究中,这些转录调节的重要性是难以低估。 核受体是可溶性蛋白,可以绑定到特定的DNA调控元件(反应元件或RES),并在转录中作为细胞类型和特异性启动子的调节器。与其他转录因子相反,核受体的活性可以通过结合到相应的配体来调节,小的亲脂性分子能轻易地穿透生物膜。最近几年中确定的一些核受体不具有任何已知的配体,这些所谓的孤儿受体自从他们可能会导致新的内分泌调节系统的发现已吸引很多人相当大的兴趣。 在一般情况下,核受体作为均聚物和异源二聚体结合到REs上,并以倒置、外翻或直接重复排列,REs包含两个PuGGTCA核心序列的拷贝。许多启动子的转录被证明是依赖核受体的,并包含核受体RE。也有大量缺乏RE的启动子和其他基因的调控元件,通过DNA独立蛋白质-蛋白质相互作用的核受体调节,这意味着核受体介导的多层次的转录调控。据认为,有一个三维的监管空间,其中的一个基因对应一种激素的响应是由指定的三个坐标的值:细胞内容物、生理方面和基因(反应元件)方面确定[5]。 核受体结构 DNA结合结构域 所有核受体组成一个超家族并根据他们的保守结构域进行了分类。DNA结合
小离子的大功能钙离子调控T细胞抗原受体活化的机制
“小离子的大功能”- 钙离子调控T细胞抗原受体活化的机制研究 施小山,许琛琦 中科院上海生科院生物化学与细胞生物学研究所分子生物学国家重点实验室/ 国家蛋白质科学中心(上海) 众所周知,脂质同蛋白质及核酸一样是生命体必须的组分之一,由其组成的细胞质膜不仅保证着细胞的完整性,同时也调控着许多生理过程1。伴随着生命科学研究技术的发展,关于脂质调控蛋白质功能的工作越来越多地被报道出来2, 3。总体来说,脂质同蛋白质的相互作用可分成特异性及非特异性相互作用两种形式,二者分别对各类信号事件进行精细的调控。作为脂质-蛋白质非特异性相互作用的主要体现形式,二者的静电相互作用(富含碱性残基的蛋白质与酸性磷脂之间的静电相互作用)被发现广泛存在于膜蛋白受体、离子通道、粘附分子及许多其他蛋白同脂质的相互作用中,它们控制着受体的活化、离子通道的开关、细胞的粘附及其他生命过程4-7。然而,关于这种静电相互作用的解除机制迄今却仍未被揭示。最近,我们针对T细胞抗原受体(TCR)的研究发现:TCR胞内区的酪氨酸磷酸化受细胞质膜中的酸性磷脂调控。在TCR未识别抗原前,其酪氨酸信号模体(ITAM)与酸性磷脂结合从而插入到膜脂双层中,处于被“屏蔽”的状态4。而在T细胞被抗原活化后,我们发现TCR周围的钙离子浓度迅速上升,这些二价阳离子可以通过静电相互作用与酸性磷脂结合,从而打破ITAM与脂质之间的静电相互作用,帮助ITAM磷酸化,并最终扩大T细胞受体的激活信号,提高T细胞对抗原的敏感性8。 T细胞抗原受体是T淋巴细胞(简称T细胞)识别抗原并最终发挥其适应性免疫功能的基础,它是由配体识别亚基TCRαβ及信号亚基CD3εγ、εδ与ζζ共同组成的一个四亚基复合物9。TCRαβ通过基因重排的方式,利用有限的DNA序列转录并翻译成庞大的TCR受体库,从而识别机体和环境中各式各样的抗原10。而另一方面,保守的信号亚基(包括一条CD3γ链、一条CD3δ链、两条CD3ε链及两条CD3ζ链)则通过特定的免疫受体酪氨酸激活模体(ITAM)来行使其传递信号的功能9。更为有趣的是,不同CD3链带有的ITAM个数并不是完全一致。CD3γ、CD3δ及CD3ε链的胞内段各带有一个ITAM,而CD3ζ链的胞内段则带有三个ITAM。一个TCR中共有十个ITAM,可以组成多种多样的磷酸化组合,进而适应不同程度的刺激信号,传导不同程度的活化信号11。 迄今关于TCR的胞内信号通路已经得到了比较广泛的研究9。当TCRαβ识别抗原递呈细胞表面的主要组织相容性抗原分子MHC所递呈的多肽抗原(peptide-MHC复合物,简称pMHC)后,信号亚基CD3上的ITAM将被激酶Lck或Fyn磷酸化,而这种ITAM磷酸化将启动一系列的信号级联放大反应,最终调控T细胞的增殖、分化及杀伤等功能。这样一系列传统的信号级联放大反应可能有助于解释T细胞可以识别甚至单个抗原而发挥功能的超敏感性12, 13。值得一提的是,在这一系列的信号级联放大反应中,第二信使浓度的上升(比如IP3、DAG及Ca2+)起着至关重要的作用9。 同胞内信号转导过程的详尽研究不同,TCR的跨膜信号转导过程由于研究难度更大一直是困扰科学家们的大问题。换言之,TCRαβ在接受pMHC的刺激后,如何引起CD3胞内段ITAM的磷酸化一直是不甚清楚的过程。同所有的磷酸化过程一样,CD3胞内段ITAM磷酸化水平会被激酶(如Lck及Fyn)及磷酸水解酶(如CD45、SHP1及SHP2)调控14, 15。目前TCR的活化机制主要有三种假说:受体聚集、结构形变以及磷酸酶排挤模型16, 17。而所有的模型最终都通过改变TCR磷酸化平衡来启动TCR活化过程。在受体聚集假说中,pMHC的刺激会诱导TCR和共受体(CD4与CD8)产生聚集现象。共受体的胞外段可以同pMHC结合,而胞内段可以与Lck相互作用。这就使得pMHC结合TCR时,它还通过共受体把Lck招募到了TCR周围,从而增加ITAM磷酸化的可能性。另一方面,pMHC与TCR 的结合会诱导TCR分子形成聚集体,这种聚集体有利于Lck分子克服空间位阻效应将TCR
核受体NR4A1功能及调控的研究进展_谷亚龙
·综述·核受体NR4A1功能及调控的研究进展 谷亚龙 张新东 金保方 【摘要】 核受体NR4A1是核受体NR4A家族中的重要一员,可通过对靶细胞基因转录的调 节,参与细胞的增殖、凋亡调控,在肿瘤发生、血管重塑以及类固醇合成等重要生命活动过程中发 挥重要作用,其功能受到磷酸化、蛋白质相互作用等多种途径的调控。本文就有关NR4A1的生物学 功能及其调控机理的研究进展作一综述。 【关键词】细胞增殖;细胞凋亡; NR4A1; 类固醇合成 The research progress in function and regulation of nuclear receptor NR4A1Gu Yalong, Zhang Xindong, Jin Baofang. Institute of Andrology, Nanjing University of Chinese Medicine, Nanjing 210023, China Corresponding author: Jin Baofang, Email: hexiking@https://www.360docs.net/doc/ec808790.html, 【Abstract】 NR4A1 is an important transcription factor that belongs to NR4A family. NR4A1 plays important roles in numerous cellular responses by regulating the transcription of genes from target cells, including the cell proliferation and apoptosis, tumorigenesis, vascular remodeling and steroidogenic. The function of NR4A1 is regulated by phosphorylation and other interactions among proteins. In this paper, the research progress of NR4A1 in biological function and regulation was reviewed. 【Key words】 Cell proliferation; Apoptosis;NR4A1;Steroidogenic synthesis 细胞分化是细胞类型多样性的生理基础,此过程对于每一个有机体正常功能的维持必不可少。对于所有细胞来讲,其分化和功能受多种激素、信号分子和旁分泌因子的严密调控。这些信号经过传递与转化,引起某个基因组的应答,都需要一系列转录因子的参与。明确这些因子并阐明它们的作用机制,对于我们理解所有组织细胞的分化和功能至关重要。NR4A1是NR4A家族中的一员,该家族被归类为及早反应基因家族,其可以迅速但短暂地被一些刺激诱导表达,这些刺激包括化学因素:环磷酸腺苷、佛波醇酯、生长因子、肽类激素和神经递质等;物理因素:膜去极化、机械搅拌和磁场等。研究表明,NR4A1作为一种响应蛋白在信号传导的早期发挥作用[1]。接受信号刺激后的NR4A1分子构象发生改变,通过对靶基因调控区的特异性识别和结合发挥转录调控作用,参与细胞增殖、凋亡,肿瘤发生以及机体的免疫应激、能量代谢、类固醇合 DOI:10.3877/cma.j.issn.1674-0785.2014.08.025 基金项目:国家自然科学基金(81273760);国家自然科学基金青年基金(81302969);科技部国家基础规划“973”项目(2010CB530401)作者单位:210023南京中医药大学男科学研究所 通讯作者:金保方,Email: hexiking@https://www.360docs.net/doc/ec808790.html, 成等生命活动,并受蛋白酶降解、磷酸化、蛋白质相互作用等多种途径的调控。 NR4A1广泛表达于各种组织,包括:胸腺、肌肉、肺、肝、睾丸、卵巢、前列腺、肾上腺、甲状腺、垂体[2-4]。在这些组织中,NR4A1以多种形式参与细胞功能的调控。 一、NR4A1的结构特征 NR4A1由六个功能域组成保守的模体结构:一个氨基端长度可改变的转录激活域(AF-1),一个包含两个锌指结构的DNA结合域(DBD),一个构象可以改变的铰链区,一个包含二聚化位点的配体结合域(LBD),另一个配体依赖的转录激活域(AF-2),一个功能未知的可变羧基端结构域。不同核受体的AF-1在长度上和氨基酸的组成上呈现不同,此区域负责与其他转录因子相互作用,调节转录活性。DBD包含有两个高度保守的锌指结构,由此来介导核受体与DNA特异性的相互作用。同很多其他的转录因子一样,NR4A1的DBD结构域以α螺旋的方式同位于目标DNA大沟中的识别元件结合,调控基因表达。在激素识别的特异性和生理反应的选择性方面,LBD的功能必不可少,它被认为是一个分子开关,其与特异性的配体结合改变了核受体的分子构象,使核受体具备转录活性。