骨骼肌 心肌 平滑肌
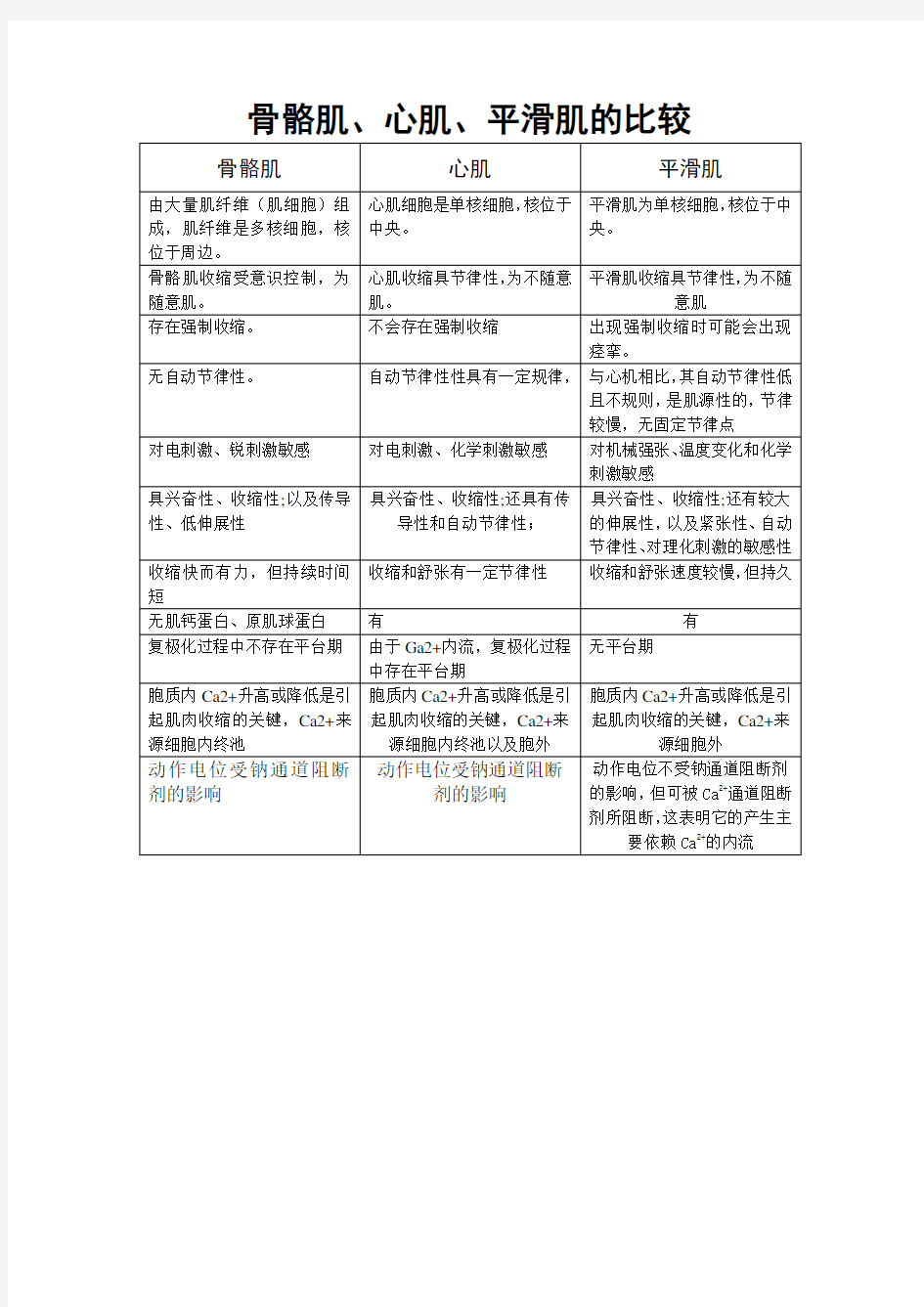
骨骼肌、心肌、平滑肌的比较
骼肌和心肌收缩的区别
骼肌和心肌收缩的区别 骨骼肌收缩可以发生强直收缩,心肌收缩不能强直收缩 造成这样的原因是肌肉的兴奋时间不同,骨骼肌动作电位快,可以在很短时间出现两次动作电位,而在肌肉收缩还未完成的时候,第二次收缩已经开始,总体效应就是强直收缩;心肌的动作电位持续时间长,并且动作电位和肌肉收缩频率几乎是同步的,所以不可能造成在肌肉收缩的时,同时发生两次动作电位来加强这种收缩。 还有一个区别就是,心肌收缩是全心同步的,骨骼肌收缩只是局部的。 骨骼肌的收缩受意识支配,收缩有力;而心肌的收缩不随意,但有节律性.目前公认的骨骼肌纤维的收缩原理是“肌丝滑动学说”.当肌纤维收缩时,细肌丝与粗肌丝的长度不变,是细肌丝在粗肌丝之间向M线方向滑动.由于是细肌丝滑入A带内,导致H带和I带变窄,甚至消失,A带宽度不变,Z线靠近,肌节缩短,既肌纤维收缩.其收缩过程是:当神经冲动传到肌纤维时,轴突末梢释放神经递质,兴奋经横小管传至三联体,引起肌浆网释放钙离子进入肌浆,肌原蛋白与钙离子结合后引起构型变化,使肌原球蛋白陷入肌动蛋白的螺旋沟内,肌动蛋白的位点暴露,粗肌丝上横桥与肌动蛋白位点接触,ATP酶被激活,分解ATP而释放能量,使横桥向M线方向转动,并将细肌丝拉向M线,肌节缩短,肌纤维收缩.收缩完成后,钙离子被肌浆网膜上的钙泵汲回肌浆网,细肌丝与粗肌丝分离,并退回原位,肌节复原,肌纤维舒张,整个收缩与舒张过程结束. 心肌收缩具有以下特点: 1.心肌的肌浆网不发达,终池贮钙的二价正离子量比骨骼肌少因而心肌细胞收缩时对细胞外液中钙的二价正离子的浓度依赖性较大. 2.心室肌的收缩期相当于有效不应期,在收缩期内心肌不能再接受刺激产生兴奋和收缩,因而心肌细胞不产生强直收缩. 3.心脏收缩具有“全或无“的特点,即心脏的收缩一旦引起,它的收缩强度就是近于相等的,而与刺激的强度无关.这是因为心肌细胞之间的闰盘区电阻很低,兴奋易于通过;另外心脏内还有特殊传导系统可加速兴奋的传导,故当某一处的细胞产生兴奋,可引起组成心房或心室的所有心肌细胞都在近于同步的情况下进行收缩.因此,可将心房和心室看成功能上的“合胞体”.
心肌与骨骼肌的区别
心肌(cardiac muscle)由心肌细胞构成的一种肌肉组织。广义的心肌细胞包括组成窦房结、房内束、房室交界部、房室束(即希斯束)和浦肯野纤维等的特殊分化了的心肌细胞,以及一般的心房肌和心室肌工作细胞。前5种组成了心脏起搏传导系统,它们所含肌原纤维极少,或根本没有,因此均无收缩功能;但是,它们具有自律性和传导性,是心脏自律性活动的功能基础;后两种具收缩性,是心脏舒缩活动的功能基??br /> 心肌细胞的结构特征心肌细胞与骨骼肌的结构基本相似,也有横纹,但在结构上具有以下几个特征:①心肌细胞为短柱状,一般只有一个细胞核,而骨骼肌纤维是多核细胞。心肌细胞之间有闰盘结构。该处细胞膜凹凸相嵌,并特殊分化形成桥粒,彼此紧密连接,但心肌细胞之间并无原生质的连续。心肌组织过去曾被误认为是合胞体,电子显微镜的研究发现心肌细胞间有明显的隔膜,从而得到纠正(参见彩图插页第37、40页)。心肌的闰盘有利于细胞间的兴奋传递。这一方面由于该处结构对电流的阻抗较低,兴奋波易于通过;另方面又因该处呈间隙连接,内有15~20埃的嗜水小管,可允许钙离子等离子通透转运。因此,正常的心房肌或心室肌细胞虽然彼此分开,但几乎同时兴奋而作同步收缩,大大提高了心肌收缩的效能,功能上体现了合胞体的特性,故常有“功能合胞体”之称。②心肌细胞的细胞核多位于细胞中部,形状似椭圆或似长方形,其长轴与肌原纤维的方向一致。肌原纤维绕核而行,核的两端富有肌浆,其中含有丰富的糖原颗粒和线粒体,以适应心肌持续性节律收缩活动的需要。从横断面来看,心肌细胞的直径比骨骼肌小,前者约为15微米,而后者则为100微米左右。从纵断面来看,心肌细胞的肌节长度也比骨骼肌的肌节为短。③在电子显微镜下观察,也可看到心肌细胞的肌原纤维、横小管、肌质网、线粒体、糖原、脂肪等超微结构。但是心肌细胞与骨骼肌有所不同;心肌细胞的肌原纤维粗细差别很大,介于0.2~2.3微米之间;同时,粗的肌原纤维与细的肌原纤维可相互移行,相邻者又彼此接近以致分界不清。心肌细胞的横小管位于Z线水平,多种哺乳动物均有纵轴向伸出,管径约0.2微米。而骨骼肌的横小管位于A-I带交界处,无纵轴向伸出,管径较大,约0.4微米。心肌细胞的肌质网丛状居中间,侧终池不多,与横小管不广泛相贴。总之,心肌细胞与骨骼肌细胞在形态和功能上均各有其特点。 心肌的生理特性心肌细胞的结构特征决定了心肌的生理特性。 自律性动物的心脏在适宜的离子浓度、渗透压、酸碱度、温湿度以及充分的氧气和能源供应等条件下,即使除去所有的神经,甚至在离体条件下,它仍然能够保持其固有的节律性收缩活动。即心肌本身具有自动节律性,简称自律性。绝大多数脊椎动物心肌的自律性是肌源性的,而不是神经源性的。鸡胚在孵化后的第2天,尚无神经纤维长入,就已经出现自律性舒缩活动。心肌细胞经过组织培养过程而新生一代的心肌细胞也有自律性。这些都是有力的证据。但在无脊椎动物,如有些节肢动物,其心肌的自律性是神经源性的,如鲎就是一例。但鲎在胚胎发育阶段,心搏自律性也是肌源性的,直到第28天神经发育完善以后,它的管状心脏的自律性搏动才变成神经源性的;切断神经后会使心搏停止。乙酰胆碱可使成年鲎心的搏动加速,而在胚胎期的鲎心则对乙酰胆碱无反应。脊椎动物和无脊椎动物中的软体动物、被囊动物的心搏自律性属肌源性;环形动物、昆虫纲动物的心搏多属神经源性。蜜蜂、蝗虫、蟋蟀、蟑螂的心搏都受外部神经和激素的调节,有些昆虫如蚕的心似有几个起搏点,因此常发生逆行性搏动。在生理情况下,哺乳动物心脏的起搏传导系统中,自律性最高的是窦房结起搏细胞,其起搏节律在整体情况下,因受神经的调节而保持于每分钟70次左右(在成年
正常人体运动学 第三章 骨骼肌
第三章骨骼肌运动学 本章内容 骨骼肌的生物学基础 骨骼肌的运动适应机制 骨骼肌的运动控制与协调 骨骼肌的功能与运动障碍 第一节肌的生物学基础 骨骼肌——动力器官(人体运动) 心肌——心脏的跳动 平滑肌——胃肠道的运动等 第一节肌的生物学基础 一、骨骼肌的功能解剖 肌细胞:肌膜、肌浆(细胞质)、细胞核。肌浆中有平行排列的肌原纤维和复杂的肌管系统。 (一)肌原纤维 纵贯肌纤维全长,直径约1-2 m。平行排列的粗、细肌丝组成。 肌小节:两条Z线之间的结构。 (二)肌丝的分子组成 粗肌丝:头部有一膨大部——横桥①能与细肌丝上的结合位点发生可逆性结合②具有ATP酶的作用。 细肌丝:肌动蛋白原肌球蛋白肌钙蛋白 ●肌钙蛋白是含有三个亚单位的复合体。亚单位I、亚单位T和亚单位C分别对肌动蛋白、原肌球蛋白和Ca2+具 有高亲和力。 ●把原肌球蛋白附着肌动蛋白上。 ●Ca2+通过和肌钙蛋白结合,诱发横桥和肌动蛋白之间的相互作用。 (三)肌管系统 包绕在每一条肌原纤维周围的膜性囊管状结构,是骨骼肌兴奋收缩耦联过程的形态学基础。 ●横小管系统:肌细胞膜从表面横向伸入肌纤维内部的膜小管系统。 ●纵小管系统:肌质网系统。 ●终池:肌质网在接近横小管处形成特殊的膨大。 ●三联管结构:每一个横小管和来自两侧的终末池构成复合体。 (四)兴奋在神经肌肉接头的传递 1.神经——肌肉接头的结构 神经——肌肉接头的结构又称为运动终板。 ①接头前膜(终板前膜) ②接头后膜(终板后膜) ③接头间隙(终板间隙) 运动神经末梢去极化 Ca2+进入神经膜内 (兴奋- 分泌耦联) ACh的释放 (神经分泌) R-ACh (化学接受) 终板电位 肌膜锋电位 肌肉收缩 (五)肌肉的兴奋—收缩耦联 终池膜上的钙通道开放==终池内的Ca2+进入肌浆==Ca2+与肌钙蛋白结合==肌钙蛋白的构型改变==原肌球蛋白位移,
骨骼肌与心肌的生理特性比较研究
骨骼肌与心肌的生理特性比较研究 实验目的:了解骨骼肌收缩总和现象; 通过对期前收缩和代偿间歇的观察,探讨心肌每发生一次 兴奋后,其兴奋性发生什么变化。 研究内容:骨骼肌与心肌在收缩特性,对胞外Ca2+、K-及Na+离子浓度改变的反应不同以及应对收缩负荷(前负荷、后负荷) 的反应。 拟采取的研究方法和实验方案:取蟾蜍一只,用刺蛙针刺毁其脑脊髓并制备蟾蜍(或蛙)的坐骨神经-腓肠肌标本。收缩总和: 启动波形显示图标,调节扫描速度为5~10nm/s,调节单收 缩幅度为1.5cm左右。调节刺激设置为双刺激方式,并使 两个阈上刺激强度相等。先调节刺激间隔大于单收缩的时 程,然后逐渐缩短刺激间隔,分别观察并记录肌肉收缩形 式的变化。强直收缩:减慢扫描速度(5mm/s)并衰减 振幅增益,使单收缩的幅度减小至3~5mm。调节刺激设 置为串刺激方式,分别以1.5次/s(串长为5)、6次/s(串长 为10)、10次/s(串长为20)、30次/s(串长为35)和45次/s(串 长为50)的频率刺激标本,观察肌肉收缩曲线的变化。 暴露青蛙心脏,心肌在经历一次兴奋后,其兴奋性会发生 一系列周期性的变化。心肌兴奋后的兴奋性变化特点是其 有效不应期特别长,约相当于心动周期的整个收缩期甚至 包括舒张早期。在此期中,任何强大的刺激均不能使之产
生动作电位。此后为相对不应期,可对强刺激产生动作电位。最后为超常期。后两期均发生在心肌的舒张期内。因此,在舒张期内(有的不包括舒张早期),给予心室一次阈上刺激,便可以在正常节律性兴奋到达心室之前,引起一次扩布性的兴奋和收缩。而正常的节律性兴奋到达时,心肌正好处于期前收缩的有效不应期,因而不能引起心室的兴奋和收缩,此时心室停留舒张状态。直至下一次正常节律性兴奋到达时,才恢复正常的节律性收缩。