数量遗传学知识点总结
遗传学-第10章-数量遗传

VF 2
玉米穗长试验结果 VF1=2.307,VF2=5.072, 在该组合中,穗长的广义遗传率为: Hb2=(5.072-2.307)/5.072×100%=54% 在该杂交组合中, F2 穗长的变异大约有 54%是由于遗传差异造成的,46%是环境 影响造成的。
三. 狭义遗传力的估算方法
例如小麦子粒颜色的遗传动态
P F1 F2 红R1R1R2R2 白r1r1r2r2 R1r1R2r2 红 1 4 6 4
1
4R
深红
3R
中深红
2R
中红
1R
淡红
0R
白色
P F1
F2
红R1R1R2R2R3R3 白r1r1r2r2r5R 4R 3R 2R 1R
VA H 100% VG VE
2 N
VA = 100% VA VD VI VE
(1) 不易受环境影响的性状的遗传率比较 高,易受环境影响的性状则较低; (2) 变异系数小的性状遗传率高,变异系 数大的则较低; (3) 质量性状一般比数量性状有较高的遗 传率; (4) 性状差距大的两个亲本的杂种后代, 一般表现较高的遗传率; (5) 遗传率并不是一个固定数值,对自花 授粉植物来说,它因杂种世代推移而 有逐渐升高的趋势。
一、遗传率(heritability)的概念
表现型是基因型和环境条件共同作用的结果。
具有相对性状的两个亲本杂交,后代的性状表 现型值的差异取决于两方面的因素,一是基因 的分离造成的,一是环境条件的影响造成的。
遗传率:在一个群体中,遗传方差在总 方差(表现型方差)中所占的比值。
广义遗传率定义为:
G A D I
加性效应,A:等位基因和非等位基因 的累加效应,可固定的分量 显性效应,D:等位基因之间的互作 效应, 属于非加性效应 上位性效应, I: 非等位基因之间的相 互作用, 属于非加性效应
动物遗传学-第九章 数量遗传学基础

根据特定资料估计的遗传力,只能作为一个估计值看待。
n
P
P1 P2 Pn
Pii 1nn Nhomakorabea同理可得:
n
Gi
G i
,
n
n
Ei
E i n
若E与G之间相互独立,在一个随机交配的大群体中
n
Ei 0 P G
i
群体均值能否代表群体基因型均值取决于群体大小,群 体越大,代表性愈强。
一、遗传力(heritability):
(一)概念: 亲代传递其遗传特性的能力。 或指性状的遗传方差在总方差(表型方差)中 所占的比率。
二、简单性状和复杂性状
简单(遗传)性状 (simply-inherited trait) 受很少数基因的控制,而且几乎不受环境变化的影响。
如孟德尔豌豆试验中所列举的性状都是简单(遗传)性 状。
复杂性状 (complex trait) 受多个基因的作用,而且易受遗传或非遗传因素的影
响。多在医学上使用。如糖尿病。
(二)对遗传力的理解注意事项:
遗传力是描述性状的一个特征量,遗传力大说 明受到遗传的影响大,遗传力低则环境的作用大。 性状遗传力的高低并不表示性状的好坏与畜群 的好坏。
0<h2 <1
(三)若干性状的遗传力
性状 初生重 断奶重 成年体重 椎骨数 乳头数 背膘厚 眼肌面积
表1 表猪1 一猪一些些性性状状的遗遗传传力 力
第九章 数量遗传学基础
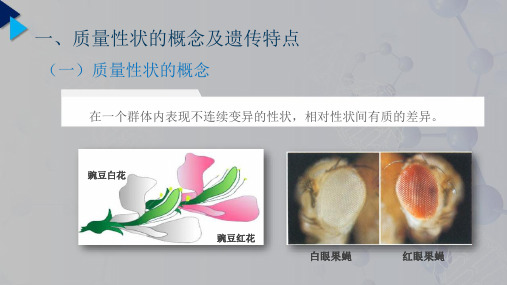
一、质量性状和数量性状
质量性状(qualitative trait) —遗传基础是单个或少数几个基因的作用,它的 表型变异是间断的。如牛的无角与有角,兔的白 化与有色。
数量遗传学知识点总结

第一章绪论一、基本概念遗传学:生物学中研究遗传和变异,即研究亲子间异同的分支学科。
数量遗传学:采用生物统计学和数学分析方法研究数量性状遗传规律的遗传学分支学科。
二、数量遗传学的研究对象数量遗传学的研究对象是数量性状的遗传变异。
1. 性状的分类性状:生物体的形态、结构和生理生化特征与特性的统称。
如毛色、角型、产奶量、日增重等。
根据性状的表型变异、遗传机制和受环境影响的程度可将性状分为数量性状、质量性状和阈性状3 类。
数量性状:遗传上受许多微效基因控制,性状变异连续,表型易受环境因素影响的性状,如生长速度、产肉量、产奶量等。
质量性状:遗传上受一对或少数几对基因控制,性状变异不连续,表型不易受环境因素影响的性状,如毛色、角的有无、血型、某些遗传疾病等。
阈性状:遗传上受许多微效基因控制,性状变异不连续,表型易受或不易受环境因素影响的性状。
有或无性状:也称为二分类性状 ( Binary traits)。
如抗病与不抗病、生存与死亡等。
分类性状:如产羔数、产仔数、乳头数、肉质评分等。
数量性状的特点:必须进行度量,要用数值表示,而不是简单地用文字区分;要用生物统计的方法进行分析和归纳;要以群体为研究对象;组成群体某一性状的表型值呈正态分布。
3. 决定数量性状的基因不一定都是为数众多的微效基因。
有许多数量性状受主基因(major gene)或大效基因(genes with large effect)控制。
果蝇的巨型突变体基因( gt);小鼠的突变型侏儒基因( dwarf, df);鸡的矮脚基因( dw);美利奴绵羊中的Booroola 基因( FecB);牛的双肌( double muscling)基因( MSTN);猪的氟烷敏感基因( RYR1)三、数量遗传学的研究内容数量性状的数学模型和遗传参数估计;选择的理论和方法;交配系统的遗传效应分析;育种规划理论。
四、数量遗传学与其他学科间的关系理论基础奠定:孟德尔遗传学+数学+生物统计学理论体系完善:与群体遗传学关系最为密切;学科应用:与育种学最为密切,是育种学的理论基础和方法论;学科发展:与分子生物学、生物进化学、系统科学和计算机科学密切结合,并产生了新的遗传学分支学科,如分子数量遗传学等。
数量遗传学的基础 第一节 性状的分类

性状遗传变异的动态; 必须以群体为研究对象。
数量性状的特点
变异的连续性 对环境的敏感性 分布的正态性 多基因作用的微效性
数量性状的特征
对质量性状而言: • 不易受环境的影响而发生变异 • F1有显隐性之分 • F2代可明显分组,不会出现超亲遗传的现象
对数量性状而言:
玉米穗长(表)
• 易受环境的影响而发生变异 • 两个纯合的亲本杂交,F1代一般是双亲的中间类型,但有时
人的身ห้องสมุดไป่ตู้(图)
遗传特征:两个有差异的亲本杂交
1. F1为双亲中值,但有时会偏向某一亲本; 2. F2均值、F1均值、双亲中值相近 3. F2变幅增大,但是变异程度远远超过F1 4.易受环境条件影响 5.有超亲遗传的现象:当杂交的亲本不是极端类型的时候,杂 交后代中有可能会出现高于高值亲本或低于低值亲本的类型
质量性状与数量性状的关系
1 由于区分的着眼点不同 2由于杂交亲本的差别基因对数不同。数量性状一般由微效 多基因控制,但在某些情况下,也可由单一的主基因控制。 3 一因多效,一个基因可以同时控制质量性状和数量性状, 控制数量性状的基因与控制质量性状的基因可能连锁。
三、 阈性状
可以用等级或分类表示的性状,如猪肉 颜色分为5级;蛋黄颜色分为9级。
第十四章 数量遗传学基础
一、数量遗传学(quantitative genetics)
1.概念 “是遗传学原理与统计学方法相结合,研究
群体数量性状的遗传学分支科学。” 遗传学 统计学 群体 数量性状
动物生产中需要改良的性状大多是数量性状。
第一节 性状的分类
遗传学_ 数量性状遗传_
个体的基因型
✓ 个体性状的表现型数值,称为表现型值,以P表示。 ✓ 表现型值有两部分组成:
一个是基因型所决定的数值,称为基因型值,以G表示; 一个是环境条件引起的变异,用E表示。 ✓ 表现型值、基因型值,和环境变异值三者之间的数量关 系可用以下公式表示:P=G+E
环境条件的影响
✓ 表型变异用表型方差(即总方差)VP表示; ✓ 遗传变异用遗传方差(即基因型方差)VG表示; ✓ 环境变异用环境方差VE表示。 ✓ 三者的数量关系可用下式表示:Vp=VG +VE
三、纯系学说
(三)纯系学说的发展
“ 纯系的纯是相对的、暂时的,绝对的纯系
是不存在的,纯系内继续选择可能是有效的。 纯系繁育过程中,由于突变、天然杂交和机械 混杂等因素必然会导致纯系不纯,产生新的遗 传变异,可能出现更优个体。
“
遗传率及估算方法
一、数量性状变异的表示方法
生物性状 表现的 决定因素
超矮秆表型是由于D18的突变导致。 该种突变体除株高显著降低后,其他 农艺性状与野生型无显著性差异。
小麦粒色简单划分,表现质量性状,单细致 观察,籽粒颜色红到白,表现连续变异,数量性 状的特点。
二 、数量性状的概念及遗传特点
(三)数量性状和质量性状的相对性
生物还有一些性状为阈性性状: 表型呈非连续变异,而其基本物质 的数量呈潜在的连续变异的性状, 即只有超越某一遗传阈值时才出现 的性状,如动植物甚至包括人类的 抗病力、死亡率以及单胎动物的产 仔数等性状。
3 数量性状对环境条件的变化反应敏感。
4 研究方法上,依靠群体,必须用统计方法,对在杂种和后代进行分析。
二 、数量性状的概念及遗传特点
(三)数量性状和质量性状的相对性
第九章 数量遗传学

i
x)
2
n
方差和标准差是全部观察值偏离平均数的重要度量参数
S 就称为标准差(standard devi
或叫做标准误(standard error
S---标准差
均值和标准差决定了表型的分布 一般:育种要求标准差大,则差异大,利于单株的选择;
良种繁育场则标准差小,差异小,可保持品种稳定。
四、标准误
1 2 (1/4)1=1/4 (3)1=3 3
(a+b)2
2
3
4
6
(1/4)2=1/16
(1/4)3=1/64
(3)2=9
(3)3=27
5
7
(a+b)4
(a+b)6
4
n
8
2n
(1/4)4=1/256
(1/4)n
(3)4=81
(3)n
9
2n+1
(a+b)8
(a+b)2n
多基因假说要点: 1.决定数量性状的基因数目很多; 2.各基因的效应相等;
F1 F2 (A1A2B1B2C1C2)熟期介于双亲之间 27种基因型 (其中A1A1B1B1C1C1的个体将比晚熟亲本更晚,
而A2A2B2B2C2C2的个体将比早熟亲本更早)
第二节 数量性状的研究方法
需要分析杂交后代的大量个体→ 应用数理统计等方法
→ 分析平均效应(mean)、方差(variance)、协 方差(covariance )等遗传参数→ 发现数量性状遗传 规律。
数量性状(quantitative trait)。 如:人高、动物体重、植株生育期、果实大小,产量 高低等。 表现型变异分析推断群体的 遗传变异借助数量统计的分析方法 分析数量性状的遗传规律。
(整理)数量遗传学

(整理)数量遗传学质量性状:指由一对或对基因控制,在个体间能够明显区分,呈不连续性变异的性状。
数量性状:由微效多基因控制,在群体中不能明显区分,呈连续性变异的性状。
门阈性状:由微效多基因控制的,在群体中呈不连续分布的性状,一般能够明显地区分其表现形式。
数量遗传学:指用数理统计方法和数学分析方法研究数量性状遗传和变异规律的科学。
选择:在人类和自然干预下,某一群体的基因在世代传递的过程中,某种基因型个体的比例所发生的变化现象,称作选择。
适应度:比较群体中各种基因型(以个体平均留种子女数为标准)生存适应力的相对指标。
适应度就是特定基因型的留种率和群体最佳基因型留种率之比值。
选择系数:1减去适应度就是该基因型的选择系数。
留种率+淘汰率=1遗传漂变:如果群体规模较小,下一代的实际基因频率都可能由于抽样误差而偏离理论上应有的频率。
始祖效应:当来自大群体的一个小样本在特定环境中成为一个新的封闭群体,其基因库仅包括亲本群体中遗传变异的一小部分,并在新环境中承受新进化压力的作用,因而最终可能与亲本群分体。
这种过程在体现的般规律,称为始祖效应。
瓶颈效应:当大群体经历一个规模缩小阶段之后,以及在漂变中改变了基因库(通常是变异性减少)又重新扩大时,基因频率发生的变化。
同型交配:如果把同型交配严格地定义为同基因型交配,那么近交和同质选配都只有部分的同型交配,只有极端的近交方式——自交才是完全同型交配。
群体遗传学:专门研究群体的遗传结构及其变化规律的遗传学分支学科。
群体:是指一个种、一个变种、一个品种或一个其它类群所有成员的总和。
孟德尔群体:在个体间有相系交配的可能性,并随着世代进行基因交流的有性繁殖群体。
基因库:以各种基因型携带着各种基因的许多个体所组成的群体。
亚群:由于各种原因的交配限制,可能导致基因频率分布不均匀的现象,形成若干遗传特性有一定差异的群落通常称为亚群。
随机资本:在一个有性系列的生物群体中,任何一个雌性式雄性的个体与其任何一个相反性别的个体交配的机率是相同的。
动物遗传学-数量遗传学基础

3 数量性状表型值的剖分
16
由于D和I带有一定的随机性,一般将它们与环境偏 差合并,统称为剩余效应。
17
第二节 通径理论
18
1. 概念
自然界任何两个或多个事物之间的关系不外乎两种情况:一 种是平行关系;另一种是因果关系。 所谓平行关系是指事物间不分因果,只是互为因果或两者具 有共同原因以致相互关联。 例如:猪的瘦肉率部分决定于臀肉率,部分决定于腰肉率。 臀肉率与腰肉率有一定关系,但在它们之间很难确定哪个是 “因”,哪个是“果”。
2 同一个纯系,甚至是同一个植株所结种子,重量差异更大,变 化范围20-70mg,但这种差异在相继世代是不能遗传的,纯系 内的差异是由于环境条件造成的,在纯系内选择是无效的。
例如,在他的第15个纯系中选出重量为20mg和60mg的种子种植, 其后代平均种子重量分别为46.9mg和45.6mg,均接近该纯系群体 的平均种子重量45.0mg。
44
将控制一个数量性状的所有基因座上基因的 加性效应总和称为基因的加性效应值,它可以 通过育种改良稳定改进。个体加性效应值的高 低反映了它在育种上的贡献大小,因此也将这 部分效应称为育种值。
45
一、遗传力
数量性状的表现既受到个体遗传基础的控制, 同时又受到所处环境条件的影响,而且这两种不同 的效应又可以作进一步的剖分。应该如何从量的角 度来推断出影响一个数量性状表现的遗传效应有多 大?环境效应有多大?
13
3.控制数量性状的微效基因(Minor effect polygenes)与控制质 量性状的宏效基因(Major gene)都处于细胞核的染色体上, 多基因的遗传行为同样符合遗传基本规律,具有复制、分离和 重组、连锁和交换的特性。
4.由于微效基因的效应微小,多基因(Polygenes)并不能予以 个别辨认,只能按性状的表现在一起研究,对所涉及的基因对 数做粗略的估计。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第一章绪论一、基本概念遗传学:生物学中研究遗传和变异,即研究亲子间异同的分支学科。
数量遗传学:采用生物统计学和数学分析方法研究数量性状遗传规律的遗传学分支学科。
二、数量遗传学的研究对象数量遗传学的研究对象是数量性状的遗传变异。
1.性状的分类性状:生物体的形态、结构和生理生化特征与特性的统称。
如毛色、角型、产奶量、日增重等。
根据性状的表型变异、遗传机制和受环境影响的程度可将性状分为数量性状、质量性状和阈性状3类。
数量性状:遗传上受许多微效基因控制,性状变异连续,表型易受环境因素影响的性状,如生长速度、产肉量、产奶量等。
质量性状:遗传上受一对或少数几对基因控制,性状变异不连续,表型不易受环境因素影响的性状,如毛色、角的有无、血型、某些遗传疾病等。
阈性状:遗传上受许多微效基因控制,性状变异不连续,表型易受或不易受环境因素影响的性状。
有或无性状:也称为二分类性状(Binary traits)。
如抗病与不抗病、生存与死亡等。
分类性状:如产羔数、产仔数、乳头数、肉质评分等。
必须进行度量,要用数值表示,而不是简单地用文字区分;要用生物统计的方法进行分析和归纳;要以群体为研究对象;组成群体某一性状的表型值呈正态分布。
3.决定数量性状的基因不一定都是为数众多的微效基因。
有许多数量性状受主基因(major gene)或大效基因(genes with large effect)控制。
果蝇的巨型突变体基因(gt);小鼠的突变型侏儒基因(dwarf, df);鸡的矮脚基因(dw);美利奴绵羊中的Booroola基因(FecB);牛的双肌(double muscling)基因(MSTN);猪的氟烷敏感基因(RYR1)三、数量遗传学的研究内容数量性状的数学模型和遗传参数估计;选择的理论和方法;交配系统的遗传效应分析;育种规划理论。
四、数量遗传学与其他学科间的关系理论基础奠定:孟德尔遗传学+数学+生物统计学理论体系完善:与群体遗传学关系最为密切;学科应用:与育种学最为密切,是育种学的理论基础和方法论; 学科发展:与分子生物学、生物进化学、系统科学和计算机科学密切结合,并产生了新的遗传学分支学科,如分子数量遗传学等。
五、数量遗传学与群体遗传学的关系群体遗传学以孟德尔定律为依据,分析群体内控制质量性状的主基因的活动及其消涨规律,着重于基因频率变化规律的探讨。
其基本原理可用于育种学中质量性状的遗传改良。
数量遗传学着重分析群体数量性状的遗传变异规律,主要研究群体内控制数量性状的多基因的数量效应。
其重点在于通过统计分析估计各种遗传变异的数量参数,进而用于育种学中数量性状的遗传改良。
第二章 数量遗传学基础第一节 均数与方差一、数量性状表型值的剖分数量性状的表型值,即观察值,是由遗传与环境共同作用的结果,即 P = G + E + IGE 其中,P 为表型值,G 为基因型值,E 为环境偏差,IGE 为遗传与环境效应间的互作。
通常,假定遗传与环境间不存在互作,即IGE=0,则有:P = G + E 基因型值G 是由基因的加性效应(additive effect, A )、显性效应(dominant effect, D )和上位互作效应(epistatic interaction, I )共同作用的结果。
假定3种遗传效应间的互作为0,则G = A + D + I 式中的D 和I ,由于世代传递中的分离和重组,不能真实遗传,因而在育种中不能被固定;而加性效应值A 则能稳定地遗传给后代,因此,育种中又称之为育种值。
二、表型值:一个多基因系统控制的数量性状能够直接度量或观察的数值。
基因型值:表型中由基因型决定的那部分数值。
环境偏差: 表型值与基因型值的离差。
加性效应:等位基因间和非等位基因间的累加作用引起的遗传效应。
显性效应:同一基因座上等位基因间的互作所产生的遗传效应。
上位效应:不同基因座间非等位基因相互作用所产生的遗传效应。
环境偏差又可剖分为一般环境偏差Eg 和特殊环境偏差Es ,即E = Eg + Es ,综上所述,有:P = G + E = A + D + I + Eg + Es ,从育种学角度来看,上式中,只有A 可以真实遗传,通常将A 和D 合并到环境偏差中,称为剩余值(residual value, R ),即:P=A+R大群体中,D 、I 和E 的值有正有负,则: P G E P G N N N ==+=∑∑∑即: 而: A D I A G A N N N N =++==∑∑∑∑故: P G A ==0D I E ===∑∑∑三、一般环境:是指影响个体全身的、时间上是持久的、空间上是非局部的环境。
例如奶牛在生长发育早期营养不良,生长发育受阻,成年后无法补尝,影响是永久的。
特殊环境:是指暂时的或局部的环境。
例如,成年奶牛因一时营养条件差而泌乳量减少,但如果环境有了改善,其产量仍可恢复正常。
永久性环境:对某一特定个体的性能产生持久影响,而且是以相似的方式影响一个个体的每个记录的环境。
暂时性环境:只对某一特定性能产生影响的环境。
永久性环境和暂时性环境的剖分,是针对重复测定性状而言的。
群体的平均表型值就等于平均基因型值,也等于平均育种值。
四、群体平均值显性水平与显性度设一对等位基因A 1、A 2的频率分别为p 和q ,三种基因型A 1A 1、 A 1A 2 、A 2A 2的基因型值分别为+a 、d 、和-a 。
其中d 决定于基因的显性程度大小,即显性水平。
基因型值的标准尺度群体平均值的计算 ①用上式计算出的群体平均基因型值也等于群体的平均表型值(各基因型值是以与两纯合子平均值的离差度量的);②涉及多个基因座时,根据加性原理,由多个基因座产生的群体平均值是各基因座各自贡献之和,即: MP =∑a (p - q) +2∑pqd五、基因的平均效应概念:在一个群体内,携带某一基因的配子,随机和群内的配子结合,所形成的全部基因型的均值与群体平均基因型值的离差。
计算:设A 1、A 2基因的平均效应值分别为α1、α2,A 1可以与A 1、A 2 形成两种基因型A 1A 1、A 1A 2,其均值为pa+qd ;同样A 2可以与A 1、A 2形成两种基因型A 1A 2、A 2A 2,其均值为pd –qa 。
配 子 产生基因型的频率 基因型平均值A 1A 1(a) A1A 2(d) A 2A 2(-a) A1 p q - pa+qd显性水平 显性基因 d负向超显性 负向完全显性 负向部分显性 无显性 正向部分显性 正向完全显性 正向超显性 A2 A2 A2 无 A1 A1 A1 d < -a d = -a 0>d>-a d = 0 0<d<ad = a d > a基因型 基因型频率(f ) 基因型值 (x ) 频率×基因型值(fx ) A 1A 1 p 2 +a p 2a A 1A 2 2pq d 2pqd A 2A 2 q 2 -a -q 2a 群体平均值= ∑基因型频率×基因型值 M = ∑fx =[ p 2a+2pqd+q 2 (-a) ]=a (p - q) + 2pqdA 2 - p q pd-qaα1 =[ pa + qd ]-[ a (p - q)+2pqd ] =q [a+d (q -p) ] α2 =[ pd - qa ]-[ a (p - q) +2pqd ]=-p [a+d (q -p) ]基因替代的平均效应(两个平均效应之差)设α1与α2之差为,即:α=α1-α2=a +d (q - p),于是: α1=α+α2=q α;α2=α1-α=-p α;α被称为基因替代的平均效应六、育种值(BV )概念:育种值即加性遗传效应值,为组成某一基因型的两个等位基因平均效应之和。
计算:A (A 1A 2) =2α1= 2q α; A (A 1A 2) =α1+α2= (q - p)α; A (A 1A 2) =2α2 = -2p α说明:育种值是用群体平均值的离差表示的;一个HW 平衡的大群体,平均育种值等于0,即:Ā=ΣfA=2p 2q α+2pq (q - p)α-2q 2p α=2pq α(p+q-p-q)=0;如用绝对值表示,则平均育种值等于平均基因型值,也等于平均表型值。
七、显性离差(显性遗传效应)概念:考虑一个基因座时, 特定基因型值G 与育种值A 之差, 称为显性离差,常用D 表示。
计算:将各基因型值表示为与群体平均值的离差:G d (A 1A 1) = a-M =2q(α-qd);G d (A 1A 2) = d- M =(q-p)α+2pqd ; G d (A 2A 2) =-a- M =-2p(α+pd)D = Gd-A ,有D (A 1A 1) = Gd (A 1A 1)–A (A 1A 1) = -2q 2d ;D (A 1A 2) = Gd (A 1A 2)–A () = 2pqd ;D () = Gd ()–A () = -2p 2d所有基因型的显性离差都是平均显性离差值为0,即:D =ΣfD= -2p 2q 2d + 4p 2q 2d - 2p 2q 2d= 0八、上位互作离差如果考虑两个以上的基因座, 基因型值可能包含基因座间非加性组合产生的互作离差。
令G A 和G A 分别为A 、B 二基因座的基因型值,则I AB 为两个基因座基因的互作离差,即:G = G A + G B + I AB由于数量性状涉及的基因座很多,互作的情况相当复杂,难以将各单一基因间的作用都区分开来。
就一群体而言,∑I = 0。
九、数量性状表型方差的剖分假定,遗传效应间、环境效应间及遗传及环境效应间无互作,即不考虑协方差的情况,则:V P = V G + V E =V A + V D + V I + V Eg + V Es 式中,V G 称为基因型方差,V A 称为加性遗传方差,V D 称为显性方基因型 A 1A 1 A 1A 2 A 2A 2频率(f ) p 2 2pq q 2基因型值(G) +a d -a离差基因型值 (Gd) 或 2q (a-pd) a(q-p)+d(1-2pq) -2p (a+qd) 2q (α-qd) (q-p)α+2pqd -2p (α+pd)育种值(A) 2q α (q - p)α -2p α显性离差(D) -2q 2d 2pqd -2p 2d差,V I称为互作方差,V D+V I=V NA称为非加性遗传方差,VE称为环境方差。
V Eg和VEs分别为一般和特殊环境方差。
育种值方差:V A=ΣfA2=p2 (2qα)2 + 2pq [(q-p)α]2 + q2 (-2pα)2=2pqα2 =2pq[a+d(q-p)]2显性遗传方差:V D=∑fD2= p2 (-2q2d)2 + 2pq (2pqd)2 + q2 (-2p2d)2 = (2pqd)2基因型值方差:若d = 0,即无显性时,VG = V A = 2pqα2若d = a,即完全显性时,V G = V A + V D= 8pq3a2+ 4p2q2a2= 4p2q2a2(1+q)若0<d<a,即不完全显性时,V G = V A + V D= 2pq[a+d(q-p)]2 + [2pqd]2均数、方差与协方差第二节数量性状的遗传机制微效多基因假说一、多基因:数量性状是由许多基因的联合效应控制的。