甲烷氧化菌_ppt课件
甲烷氧化菌

❖甲烷氧化菌中MMO 以两种形式存在, 一种是以可 溶性形式存在于细胞质中, 称为可溶性甲烷单加氧 酶( sMMO) ; 另一种是以颗粒形式存在于细胞膜 上, 称为颗粒性甲烷单加氧酶( pMMO)。
a
6
根据形态、GC%、代谢途径、膜结构、主要磷脂酸成分等系 列特征, 可将甲烷氧化菌分为二种: Ⅰ型和Ⅱ型
❖ Ⅰ型甲烷氧化菌包括Methylomonas、Methy lobacter 、Methylococcus、Methy lomicrobium、 Methylocadum、Methy losp haera 等6 属, 它们 利用5-磷酸核酮糖途径( RuMP Pathway) 同化甲 醛, 主要含16-C 脂肪酸, 胞内膜成束分布。
a
14
样品前处理
❖16组样品分8次制备样品,放置在25度黑暗 环境中,每隔3天,拿出其中2个微模型, 对其土壤进行拌匀,放液氮中保存,做好 进一步分析的准备。
a
15
T—RFLP:末端限制性片段长度多态性技术
❖是以荧光标记引物PCR为基础,根据末端限 制性片段长度区分出微生物群体组成的一 种微生物群体图谱法。
a
3
❖虽然大气中甲烷的含量仅为二氧化碳的1/ 27, 但每摩尔甲烷引起气候变化的作用是每 摩尔二氧化碳的20~30倍。因此, 减少甲烷 排放要比减少等量的二氧化碳排放, 对减少 温室效应的贡献要大的多。
a
4
❖甲烷是一种重要的温室气体, 近200 年来其 在大气中的含量以每年1%的速度急剧增加, 主要是由于甲烷排放源的增加和甲烷汇的 减少。80% ~ 90% 的甲烷来源于生物活动, 而甲烷的唯一生物汇为土壤里甲烷氧化细 菌的氧化作用, 大约占大气甲烷汇的10%。
甲烷氧化菌综述2

甲烷氧化菌国内外研究现状国外对甲烷厌氧氧化的研究开始得较早。
自1976年WiHiam Reeburgh在海洋沉积物中发现甲烷厌氧氧化作用开始,至今已有30多年。
近几年来,荷兰Nijmegen Radboud大学微生物学系对甲烷厌氧氧化的研究相对较多。
该课题组通过研究证实了多种环境中甲烷厌氧氧化的存在,尤其是在湿地幢和泥火山等1等极端环境中。
且通过分子生物学方法,从中检测出了甲烷厌氧氧化菌,并对其进行了生理生态分析,证实了甲烷厌氧氧化并非只能在硫酸盐存在时才能发生,甲烷厌氧氧化可与脱硝作用耦合,不仅消耗了甲烷,同时降低了环境中氮的含量一J。
虽然有研究者在实验室培养出具有甲烷厌氧氧化活性的混培物,并对个别甲烷厌氧氧化菌进行了分离鉴定,但是如何缩短甲烷厌氧氧化活性培养时间、探明甲烷厌氧氧化的生物学机制、运用甲烷厌氧氧化工艺还有待继续研究。
国内对甲烷厌氧氧化的研究相对较少。
2005年,浙江大学生命科学院吕镇梅、闵航等研究了在不同环境条件下水田土壤甲烷好氧氧化活性和甲烷厌氧氧化活性,并以此计算水田土壤甲烷厌氧氧化对整个甲烷氧化的贡献率旧瑚】。
此外,2006年,中国科学院广州地球化学研究所吴白军、周怀阳等以珠江河口淇澳岛一桂山岛一南海沉积物为研究对象,揭示了该区域沉积物甲烷厌氧氧化(AOM)过程的发生.并利用沉积物有机质的变化初步探讨AOM作用的影响因素汹瑚川。
所有这些研究均未涉及到甲烷厌氧氧化活性的培养、生物学机制的研究等问题。
甲烷厌氧氧化茵的分布根据微生物的形态、代谢途径、膜结构、主要磷脂酸成分等系列特征,可将甲烷氧化细菌分为两类:即I型甲烷氧化细菌和Ⅱ型甲烷氧化细菌,它们分属于变形菌纲(Proteobacteria)的1亚纲和d亚纲。
I型和Ⅱ型甲烷氧化菌在环境中的分布并不相同:I型甲烷氧化菌适合生长于低甲烷浓度的环境,在允许氧化菌陕速生长的环境中占优势;而Ⅱ型菌在高浓度甲烷、低氧气浓度条件下和贫营养环境下能存活得更好,从而有较广泛的分布【10l。
甲烷氧化菌的高密度培养

978-1-4244-9158-2/10/$26.00 ©2010 IEEE 283
CMBB2010
低,细胞碳源受限,导致甲烷氧化菌生长速度慢,细胞浓 度低,发酵周期长,培养困难,不能满足大规模生产的需 要。建立甲烷氧化菌高密度培养技术是促进甲烷氧化菌应 用的关键[3]。本文通过添加水溶性碳源甲醇和添加甲烷传 递体促进甲烷氧化菌细胞生长,分别考察了甲醇添加和甲 醇传递体添加对细胞生长延滞期、生长速度等的影响,为 进一步工业化应用打下基础。
2.4 生物量测定
每隔 24h 取一次样,测定发酵液在 600 nm 下的吸光 值。细胞培养 7 天后取 30mL 发酵液,9000rpm 离心沉淀 80 ℃烘干,测定细胞干重。
2.5 数据分析
菌 体 细 胞 生 长 曲 线 采 用 Origin 8.0 处 理 , 应 用 Logistic 方程模拟该菌生长的变化趋势,建立 IMV 3011 的 动态变化的数学模型,公式为[5]:
2010 First International Conference on Cellular, Molecular Biology, Biophysics and Bioengineering (CMBB)
High cell density cultivation of Methylosinus trichosporium IMV 3011
Abstract—Methanotrophs are aerobic microorganisms that can only utilize C1- such as methane and methanol as primary growth substrates. Methane is the most widely used substrate employed in cultivating methanotrophs. However, a major problem encountered in high cell density cultivation of this organism, is the solubility of methane in liquid medium. This study investigates the cultivation of M. trichosporium IMV 3011 on methanol and methane. It has been found that methanol although inhibits the growth of M. trichosporium IMV 3011 above certain concentrations, is a good alternative due to its infinite solubility in water. With the addition of 0.05% (v/v) methanol, the highest density of cultivated cells was 1.03g dry wtL-1, which were 3 times of the without methanol addition. However, a longer lag time of 60.4 h prolonged the fermentation time. Paraffin as the methane vector can also shorten the lag phase and help methanotrophs to reach high cell density. With the addition of 2.5% (v/v) paraffin, the highest density of cultivated cells was 0.72 g dry wtL-1, which were 2 times of the without methanol addition. The lag time was shortened to 11.1h. With the co-addition of 0.05% methanol and 2.5% paraffin, the highest density of cultivated cells was 1.224g dry wtL-1 . the lag time wae shortened to 18.6 h and the growth rate was increased to 0.0415h-1 .
甲烷氧化菌

根据形态、GC%、代谢途径、膜结构、主要磷脂酸成分等 系列特征, 可将甲烷氧化菌分为二种: Ⅰ型和Ⅱ型
Ⅰ型甲烷氧化菌包括Methylomonas、Methy lobacter 、Methylococcus、Methy lomicrobium、 Methylocadum、Methy losp haera 等6 属, 它们 利用5-磷酸核酮糖途径( RuMP Pathway) 同化甲 醛, 主要含16-C 脂肪酸, 胞内膜成束分布。 而Methylosinus 和Methylocystis 则属于人们所熟 知的Ⅱ型甲烷氧化菌。 Ⅱ型菌同化甲醛的途径是丝氨酸途径( Serine pathway ) , 其占优势脂肪酸为18-C 脂肪酸, 胞内 膜分布于细胞壁的周围。
T—RFLP分析的主要步骤
1.DNA 样品的准备 2.标记性DNA序列区段的选取 3.DNA的荧光标记引物PCR扩增 4.四碱基限制性内切酶的选取 5.酶切图谱的分析 T—RFLP数据的分析 克隆的应用1.DNA 样品的准备
T—RFLP分析结果依DNA 提取质量、方法等条件不 同而产生差异,进而影响微生物群体相对含量的正 确估计。 综合各种因素考虑,建议在进行样品DNA 提取时, 要重复采样,避免取样产生的误差,并将样品均匀 混合后采用同一方法提取DNA 且设置2个或者更 多的重复,并混合DNA样品进行下一步的分析,以 确保T—RFLP 技术对样品微生物分析的准确性和真 实性。 样品DNA 由于其组分的复杂性,为保证后续分析 质量,最好用试剂盒或者凝胶电泳切胶等措施进一 步纯化,以去除多糖、有机酸等对后续PCR 分析 的影响。
5.酶切图谱的分析
选择好限制性内切酶后,根据说明对样品或者分离 菌的纯化PCR 产物进行酶切消化。值得注意的是 不同的酶所需的消化温度不同。研究表明,不完全 消化会对T—RFLP结果产生负面影响,而酶浓度过高 还会产生过多的假片段峰。 测序仪有凝胶电泳和毛细管电泳2种。 研究证明,凝胶电泳产生的T—RFLP图谱存在胶与 胶之间的差异,重复性要差于毛细管电泳。因而分 析时尽量采用毛细管电泳设备,其不仅操作方便,而 且实验结果重复性好。
甲烷氧化菌的分离鉴定及其发酵条件优化
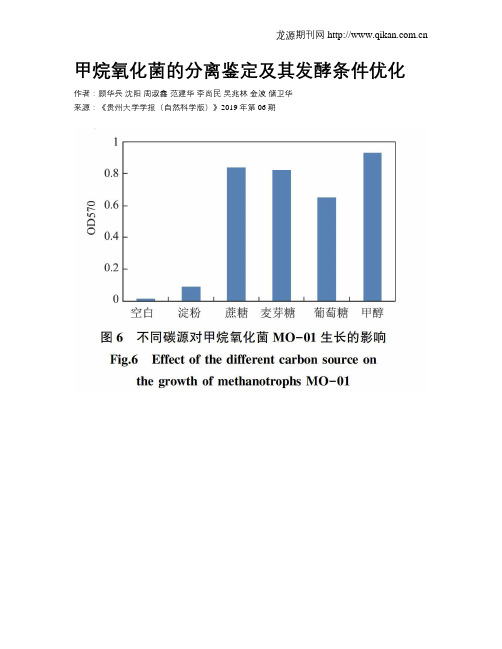
甲烷氧化菌的分离鉴定及其发酵条件优化作者:顾华兵沈阳周淑鑫范建华李尚民吴兆林金波储卫华来源:《贵州大学学报(自然科学版)》2019年第06期摘要:甲烷氧化菌是一类能以甲烷作为唯一碳源和能源进行同化和异化代谢的细菌。
本研究从污泥中分离、筛选获得一株甲烷氧化菌MO-01,根据该菌株的形态学、生理生化试验和16S rDNA序列同源性分析,证实该菌株与Methylobacterium zatmanii菌株有99%的同源性,属于Methylobacterium属。
甲烷氧化菌MO-01的实验室培养条件筛选、研究表明,该菌株以甲烷为碳源,最佳培养温度是37℃,最适PH值为7.0,铜离子浓度为30 umol/L。
本研究为今后甲烷氧化菌的放大发酵培养和动物源性单细胞蛋白的生产奠定了科学基础。
关键词:甲烷氧化菌;菌株筛选;菌种鉴定;16S rDNA中图分类号:Q-3文献标识码: A甲烷氧化菌(Methanotroph)是一种能够以甲醇、甲烷、甲酸等作为生长所需的碳源以及能源的一种甲基氧化菌[1]。
据科学测定,湿地系统内产甲烷菌产生的90%以上的甲烷气体都会被甲烷氧化菌进行氧化利用,用于合成自身细胞中的组成成分[2]。
有研究表明甲烷氧化菌能够用以生产单细胞蛋白[3]。
单细胞蛋白又称微生物蛋白,是指从纯培养的微生物细胞中提取得到的总蛋白,可作为人及动物蛋白的补充[4]。
研究已经表明,使用微生物产生的单细胞蛋白安全无毒,含有丰富的蛋白质、氨基酸和多种维生素,可以用作饲料促进畜禽生产,提高饲料利用率,代替鱼粉、大豆、骨粉、肉类和脱脂奶粉等蛋白补充饲料,具有较高的附加值[5]。
由细菌以甲烷为原料生产的新型单细胞蛋白的蛋白含量69%~80%,远远高于工业生产饲料15%~20% 的蛋白含量,具有较高的经济效益和广阔的市场空间[6-7],因此分离出能以甲烷为碳源生长的甲烷氧化细菌有着重要的意义。
本研究从污泥中分离了一株能够以甲烷为碳源和能源的菌株,并对其生物学性状进行研究,利用16S rDNA技术对筛选的菌株进行鉴定,并利用单因子控制的方法对其培养条件进行优化筛选,为后期进一步放大发酵培养和甲烷单细胞蛋白的生产提供良好科学依据。
在pH≤1条件下生存的甲烷氧化菌-氧气微电极

疣微菌门的一个新种:在pH≤1条件下生存的甲烷氧化菌泥火山、泥沸泉和火山喷气孔是不一般的地质特征,它们会有气体、水和/或半流体泥浆基质1喷出,并且伴随有相当数量的甲烷气体排放到大气中(10-1至103t y-1)2-4。
这些区域的环境条件从环境温度、中性pH到高温、低pH都有。
虽然有强烈的迹象显示在泥火山有生物学的甲烷消耗4,5,但众所周知没有甲烷氧化菌能在火山喷气孔的恶劣条件(温度高达70℃而pH值低至1.8)下繁茂生长2。
甲烷的有氧氧化第一步由可溶性或膜结合的甲烷单氧化酶进行催化。
本文报道了pmoA(编码膜结合甲烷单氧化酶的β亚基)克隆库,它们是利用从火山泥沸泉硫质喷气孔和火山喷气孔附近裸土中提取的DNA而获得的,结果显示了新的远缘pmoA基因簇。
在50℃和pH 2.0条件下对甲烷氧化菌进行富集培养后,与所有已知pmoA基因同源性都不足50%的远缘基因簇在培养菌中得到了表达。
最后,我们分离到1株嗜酸甲烷氧化菌Acidimethylosilex fumarolicum So1V,属于浮霉菌门/疣微菌门/衣原体超门6,“不属于”包括已定名甲烷氧化菌在内的α-和γ-变形菌亚门。
这种细菌生长在氧气受限条件下,以甲烷作为唯一能源,pH值可低至0.8 ——远低于曾报道过的甲烷氧化菌最佳pH值。
A. fumarolicum So1V有3种不同的pmoA基因,其中2种与泥沸泉处菌样的基因序列非常相似。
源自美国黄石公园环境高度相似的16S rRNA基因序列表明:这种新型甲烷氧化菌可能是极端环境中的常见菌种。
本文首次报道了分布广泛的疣微菌门中的代表菌与地球化学反应有关,而大部分疣微菌仍未分离鉴定6。
生产于地球地壳的大量地质甲烷一般自然地释放到大气中3,7,8。
甲烷释放的初步地球评估显示应该有足够量的甲烷资源来解释全球甲烷失踪8。
最近的Haakon Mosby和Carpatian 泥火山研究结果表明,这些系统可能也是这些地质甲烷的汇4,5,9。
甲烷厌氧氧化古菌-概述说明以及解释
甲烷厌氧氧化古菌-概述说明以及解释1.引言1.1 概述甲烷厌氧氧化古菌是一类微生物,具有重要的生态和环境意义。
它们能够利用甲烷这一常见的温室气体作为能量和碳源,通过厌氧氧化过程将其转化为二氧化碳。
这个过程减少了甲烷的释放和积累,从而有助于减缓全球变暖和气候变化的发展。
甲烷厌氧氧化古菌的发现引起了科学界的广泛关注。
在过去的几十年中,研究人员通过从不同的环境样品中分离和鉴定微生物,发现了许多具有甲烷厌氧氧化能力的古菌。
这些古菌广泛存在于湿地、海洋、农田和其他自然环境中,其数量和多样性进一步证明了其在全球碳循环中的重要性。
甲烷厌氧氧化古菌的特征也引起了研究人员的兴趣。
这些微生物具有特殊的代谢途径和功能基因,使其能够在缺氧环境中生存并利用甲烷。
甲烷厌氧氧化古菌通常以共生形式存在于特定的细菌或古菌体内,通过协同作用实现甲烷的氧化过程。
此外,研究人员还发现了一些新的甲烷厌氧氧化古菌类群,这为进一步研究甲烷循环和生态系统功能提供了新的线索。
综上所述,甲烷厌氧氧化古菌具有重要的生态和环境意义,它们在全球碳循环中发挥着重要作用。
深入了解甲烷厌氧氧化古菌的发现和特征,对于理解地球生物地球化学循环以及应对气候变化具有重要意义。
未来的研究将进一步探索甲烷厌氧氧化古菌的多样性、代谢途径和与其他生物的相互作用等方面,以期为环境保护和生态系统管理提供科学依据。
1.2文章结构本文将分为引言、正文和结论三个部分来展开讨论甲烷厌氧氧化古菌的相关内容。
在引言部分,我们将首先对甲烷厌氧氧化古菌的概述进行介绍,包括其基本特征和研究背景。
随后,我们将介绍本文的结构安排,明确各个部分的内容和组织方式。
最后,我们将明确本文的目的,即阐述甲烷厌氧氧化古菌的发现、特征,以及其在生态环境中的重要性和未来研究的方向。
接下来的正文部分,将详细介绍甲烷厌氧氧化古菌的发现和其特征。
在2.1节中,我们将回顾甲烷厌氧氧化古菌的发现历程,包括相关研究的里程碑和重要进展。
甲烷氧化细菌生物催化二氧化碳制甲醇
辛嘉英崔俊儒李树本夏春谷 中国科学院兰确化学物理磅究瞬羰基合或与选择氧化国家重点实验室.兰///,
730000.Tel:093i一8277i47,Fsx:0931-8277088,E-mail:xinjyl966氇h0}mail.COIB
二氧化碳是温室气体的主要成分,可谓取之不尽,用之不竭。通过化学或生 物途径,二氧化碳可被转化成为多种无机或有机化合物,如低碳醇、有机酸等。 其中将二氧化碳转化成为甲醇具有重大的研究意义和应用价值。它不但可以解决 温室气体二氧化碳造成气候变暖的问题,还可以为人们提供重要的化工原料和洁
同家自然科学基金重点项目(29933040),国家重点基础研究规划(G1999022406
235
MDH:Methanol Dehylrogenase MMO:Methane monooxygenase
Figure 1.Time course of methanol production by cell suspensions of methylosinus
净的燃料能源——甲醇,解决人们能源危机的问题。
由二氧化碳向甲醇的转化,有非均相催化法、电催化法、光催化法等几种具 有代表性的化学方法。但这些方法要求的条件往往都很苛刻,如非均相催化法需 要高温高压,而后两种方法则需要外加电能或光能,且转化率不高。温和条件下, 实现C02转化为甲醇是催化领域一个具有挑战性的课题。
应用生物酶催化法转化二氧化碳,条件温和,近年来备受关注。在二氧化碳 的固定和还原反应中已有应用。Robyn Obert…等采用甲酸脱氢酶(Formate Dehydrogenase).甲醛脱氢酶(Formaldehyde Dehydrogenase)和乙醇脱氢酶
在pH≤1条件下生存的甲烷氧化菌-氧气微电极
疣微菌门的一个新种:在pH≤1条件下生存的甲烷氧化菌泥火山、泥沸泉和火山喷气孔是不一般的地质特征,它们会有气体、水和/或半流体泥浆基质1喷出,并且伴随有相当数量的甲烷气体排放到大气中(10-1至103t y-1)2-4。
这些区域的环境条件从环境温度、中性pH到高温、低pH都有。
虽然有强烈的迹象显示在泥火山有生物学的甲烷消耗4,5,但众所周知没有甲烷氧化菌能在火山喷气孔的恶劣条件(温度高达70℃而pH值低至1.8)下繁茂生长2。
甲烷的有氧氧化第一步由可溶性或膜结合的甲烷单氧化酶进行催化。
本文报道了pmoA(编码膜结合甲烷单氧化酶的β亚基)克隆库,它们是利用从火山泥沸泉硫质喷气孔和火山喷气孔附近裸土中提取的DNA而获得的,结果显示了新的远缘pmoA基因簇。
在50℃和pH 2.0条件下对甲烷氧化菌进行富集培养后,与所有已知pmoA基因同源性都不足50%的远缘基因簇在培养菌中得到了表达。
最后,我们分离到1株嗜酸甲烷氧化菌Acidimethylosilex fumarolicum So1V,属于浮霉菌门/疣微菌门/衣原体超门6,“不属于”包括已定名甲烷氧化菌在内的α-和γ-变形菌亚门。
这种细菌生长在氧气受限条件下,以甲烷作为唯一能源,pH值可低至0.8 ——远低于曾报道过的甲烷氧化菌最佳pH值。
A. fumarolicum So1V有3种不同的pmoA基因,其中2种与泥沸泉处菌样的基因序列非常相似。
源自美国黄石公园环境高度相似的16S rRNA基因序列表明:这种新型甲烷氧化菌可能是极端环境中的常见菌种。
本文首次报道了分布广泛的疣微菌门中的代表菌与地球化学反应有关,而大部分疣微菌仍未分离鉴定6。
生产于地球地壳的大量地质甲烷一般自然地释放到大气中3,7,8。
甲烷释放的初步地球评估显示应该有足够量的甲烷资源来解释全球甲烷失踪8。
最近的Haakon Mosby和Carpatian 泥火山研究结果表明,这些系统可能也是这些地质甲烷的汇4,5,9。
whittenbury 甲烷氧化菌分类
whittenbury 甲烷氧化菌分类1. 介绍甲烷氧化菌甲烷氧化菌是一类能够利用甲烷作为唯一碳源和能源的微生物。
这类细菌在甲烷氧化反应中起到了关键作用,可以将甲烷氧化为甲醛和甲酸,是甲烷生物地球化学循环中不可或缺的一部分。
2. whittenbury 甲烷氧化菌的发现whittenbury 甲烷氧化菌是由 Michael Whittenbury 和他的团队在20世纪60年代初发现的。
他们首先从土壤中分离出了这种细菌,并确定了其在甲烷氧化过程中的作用。
3. whittenbury 甲烷氧化菌的分类whittenbury 甲烷氧化菌是属于一类特定的细菌裙,其分类与其他细菌略有不同。
该类细菌在分类系统中通常被归类为甲烷氧化菌属(Methylococcus) 或甲烷氧化细菌属 (Methylobacter)。
4. whittenbury 甲烷氧化菌的特征whittenbury 甲烷氧化菌具有如下特征:(1) 单细胞微生物:这类细菌为单细胞微生物,通常为球形或短杆状。
(2) 甲烷氧化能力:这类细菌具有利用甲烷氧化为甲醛和甲酸的能力,是甲烷生物地球化学循环中的关键参与者。
(3) 厌氧生长:一些 whittenbury 甲烷氧化菌在厌氧条件下能够生长和繁殖,但也有部分菌株可以在氧气充足的条件下生长。
5. whittenbury 甲烷氧化菌的应用whittenbury 甲烷氧化菌在环境保护和能源开发领域具有重要应用价值。
由于其具有利用甲烷产生有机化合物的能力,可以用于甲烷氧化反应的生物处理技术,减少甲烷的排放。
一些 whittenbury 甲烷氧化菌还被用于甲烷生物转化为生物燃料的研究中,为可再生能源开发提供了新的途径。
6. 总结whittenbury 甲烷氧化菌是一类能够利用甲烷氧化为甲醛和甲酸的微生物,其在甲烷生物地球化学循环和环境保护中具有重要作用。
通过对其分类和特征的研究,可以更好地理解这类微生物的生物学特性和应用潜力,为其在环境和能源领域的进一步利用提供理论基础。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
甲烷氧化途径的第二个酶是甲醇脱氢酶 ( methanol dehy drog enase, MDH) 。所有 革兰氏阴性甲基氧化菌都有编码甲醇脱氢 酶大亚基的mxaF基因, 其序列高度保守, 因 此MDH 可以作为这些生物在环境中存在的 很好指示剂。 Ⅰ型和Ⅱ型甲烷氧化菌在环境中的分布并 不相同:Ⅰ型甲烷氧化菌在允许氧化菌快速 生长的环境中占优势, 而Ⅱ型菌在贫营养环 境下能存活得更好,从而有较广泛的分布。
甲烷氧化细菌
根据形态差异、休眠阶段类型、胞质内膜 精细结构和一些生理特征的不同, 甲烷氧化 菌分为: 甲基单胞菌属( Methylomonas ) 甲基细菌属( Methylobacter) 甲基球菌属( Methylococcus ) 甲基孢囊菌属( Methylocyt is) 甲基弯曲菌属( Methylosinus ) 甲基微菌属( Methy lomicrobium )
几乎所有的甲烷氧化菌都能合成pMMO, pMMO 只有在铜离子浓度超0.85~1umol· g- 1细胞( 干重) 时 才表现活性。 铜离子的含量对pMMO/ sMMO 的平衡具有重要 意义。 无论培养基是否含有铜离子, 增加铜离子浓度可导 致pMMO 活性增加; 增加铜离子浓度还可以导致 合成更多的胞内膜、与pMMO 相关的膜蛋白的出 现、生长量的增加和sMMO 活性的减少。 拥有pMMO 的甲烷氧化菌比含有sMMO 的甲烷氧 化菌具有更高的生长速率和更大的甲烷亲和力, 因 此有人认为某些甲烷氧化菌合成sMMO 只是作为 在许多环境条件下铜离子限制pMMO 的活性而由 细菌产生的一种生存机制。
LOGO
甲烷氧化菌
工业常常被责为使地球变暖的“温室”气体 之源,但是农业也向大气中排放“温室”气体。
中文名称:
甲烷
英文名称:
methane;CH4
定义:
甲烷分子的结构图
一种主要由稻田和湿地释放出来的温室气体。 甲烷在自然界分布很广,是天然气、沼气、油田 气及煤矿坑道气的主要成分。它可用作燃料及制 造氢气、碳黑、一氧化碳、乙炔、氢氰酸及甲醛 等物质的原料。
根据形态、GC%、代谢途径、膜结构、主要磷脂酸成分等 系列特征, 可将甲烷氧化菌分为二种: Ⅰ型和Ⅱ型
Ⅰ型甲烷氧化菌包括Methylomonas、Methy lobacter 、Methylococcus、Methy lomicrobium、 Methylocadum、Methy losp haera 等6 属, 它们 利用5-磷酸核酮糖途径( RuMP Pathway) 同化甲 醛, 主要含16-C 脂肪酸, 胞内膜成束分布。 而Methylosinus 和Methylocystis 则属于人们所熟 知的Ⅱ型甲烷氧化菌。 Ⅱ型菌同化甲醛的途径是丝氨酸途径( Serine pathway ) , 其占优势脂肪酸为18-C 脂肪酸, 胞内 膜分布于细胞壁的周围。
微生物利用甲烷的关键酶是甲烷单加氧酶 ( MethaneMonooxygenase,MMO) 。 第一步由MMO将甲烷活化生成甲醇, 甲醇进一步 氧化为甲醛, 甲醛再同化为细胞生物量或通过甲酸 氧化为CO2, 然后经过一系列的脱氢反应生成 CO2 重新回到大气的碳库中, 即甲醇→甲醛→甲 酸盐→CO2。 甲烷氧化菌中MMO 以两种形式存在, 一种是以可 溶性形式存在于细胞质中, 称为可溶性甲烷单加氧 酶( sMMO) ; 另一种是以颗粒形式存在于细胞膜 上, 称为颗粒性甲烷单加氧酶( pMMO)。
基于sMMO 基因建立的PCR 技术在对铜离 子缺乏环境中的甲烷氧化菌研究中很有用。 在低铜离子浓度( 低于1 umol· L- 1) 的条件 下,sMMO 基因表达并产生活性。 然而更好的功能基因探针应基于pMMO 基 因, 设计出针对pmo基因的PCR 引物, 成功 用于对从许多环境样品中分离的甲烷氧化 菌DNA 的扩增。
样品前处理
16组样品分8次制备样品,放置在25度黑暗 环境中,每隔3天,拿出其中2个微模型, 对其土壤进行拌匀,放液氮中保存,做好 进一步分析的准备。
T—RFLP:末端限制性片段长度多态性技术
是以荧光标记引物PCR为基础,根据末端限 制性片段长度区分出微生物群体组成的一 种微生物群体图谱法。 末端限制性片段长度多态性技术T—RFLP具 有快速、重复性高、灵敏、高通量等优点。 虽然该技术已经应用了十几年,但是DNA 提 取、PCR 扩增、酶切以及数据分析等诸多 过程中的技术细节如果不注意,有可能引起 T—RFLP分析误差。
甲烷氧化细菌
甲烷同化细菌( methane-assimilat ing bacteria MAB)
甲烷共氧化细菌( autot rophic ammonia-oxi菌是甲基氧化菌的一个分支, 其独特之 处在于其能利用甲烷作为唯一的碳源和能源。几 乎所有的甲烷氧化菌都是专性甲烷氧化菌。
www,
实验结论
设计微模型
甲烷菌种 群假说
样品前处理
实验分析
微模型甲烷 菌种群分析 样品操作
引物分析
图形对应分析
设计微模型
模拟自然状态下水稻田里甲烷菌种群的生 活状态,这个实验选用养分充足的湿润土 壤,人工创造氧隔分界 设置16组相同的微模型组,分阶段,分析 影响种群结构的环境土壤参数
虽然大气中甲烷的含量仅为二氧化碳的1/ 27, 但每摩尔甲烷引起气候变化的作用是每 摩尔二氧化碳的20~30倍。因此, 减少甲烷 排放要比减少等量的二氧化碳排放, 对减少 温室效应的贡献要大的多。
甲烷是一种重要的温室气体, 近200 年来其 在大气中的含量以每年1%的速度急剧增加, 主要是由于甲烷排放源的增加和甲烷汇的 减少。80% ~ 90% 的甲烷来源于生物活动, 而甲烷的唯一生物汇为土壤里甲烷氧化细 菌的氧化作用, 大约占大气甲烷汇的10%。