第29章脂类的生物合成脂类物质的功能
合集下载
《生物化学》-脂质化学

概述
一、脂类物质概念 脂类是是生物体中的重要有机物,其共同点是
低(不)溶于水,高(易)溶于苯、乙醚、氯仿及 石油醚等有机溶剂;大多数脂质的化学本质是脂肪 酸和醇形成的酯及其衍生物。以及与这些化合物的 生物合成或生物功能紧密相关的一类物质。 二、脂类物质的分类 (一)按其化学组成分 (二)按其生物学功能分
2.命名与简写符号 系统名称按有机化合物命名原则进行。 十六碳脂肪酸(软脂酸) 十八碳脂肪酸(硬脂酸) 9-十六碳烯酸(棕榈油酸) 9-十八碳烯酸(油酸)
如18:0
18:1(9)
3.天然脂肪酸的结构特点
(1)一般为偶数碳原子,碳骨架长度4-36,常见 12-24,一般是不分支和无环、无羟基的单羧酸。
OH
(Sn-立体特异性编号体系) Sn -3-磷脂酸
常见甘油磷脂的极性头部和其净电荷(pH=7)
甘油磷脂名称
磷脂酸 磷脂酰乙醇胺 磷脂酰胆碱 磷脂酰丝氨酸
HO-X的名称
—— 胆胺 胆碱 丝氨酸
—X的结构
极性头基净电荷
磷脂酰甘油
甘油
磷脂酰肌醇
肌醇
H -1
HH
心磷脂
双磷脂酰甘油
例题:中性pH下,净电荷为零的 甘油磷脂是( )( )。
然而,催化加氢是一个可逆反应,饱和脂肪酸在 反应过程中,也会脱氢生成不饱和脂肪酸。这样,脱 氢的产物就可能有两种,顺式和反式。
反式不饱和脂肪酸比顺式不饱和脂肪酸空阻小,更 稳定,更容易生成,而且一旦生成,又不易被氢化饱 和。
所以,在顺式不饱和脂肪酸催化加氢的产物饱和脂 肪酸中,会含有一定量的反式不饱和脂肪酸。这就是 反式脂肪酸的由来。
影响油脂自动氧化的因素 (1)油脂的脂肪酸组成
不饱和脂肪酸越多,越容易发生自动氧化。 思考:为什么家用猪油比花生油更易变“哈喇”? 因为天然植物油脂中溶有维生素E,起抗氧化作用。
一、脂类物质概念 脂类是是生物体中的重要有机物,其共同点是
低(不)溶于水,高(易)溶于苯、乙醚、氯仿及 石油醚等有机溶剂;大多数脂质的化学本质是脂肪 酸和醇形成的酯及其衍生物。以及与这些化合物的 生物合成或生物功能紧密相关的一类物质。 二、脂类物质的分类 (一)按其化学组成分 (二)按其生物学功能分
2.命名与简写符号 系统名称按有机化合物命名原则进行。 十六碳脂肪酸(软脂酸) 十八碳脂肪酸(硬脂酸) 9-十六碳烯酸(棕榈油酸) 9-十八碳烯酸(油酸)
如18:0
18:1(9)
3.天然脂肪酸的结构特点
(1)一般为偶数碳原子,碳骨架长度4-36,常见 12-24,一般是不分支和无环、无羟基的单羧酸。
OH
(Sn-立体特异性编号体系) Sn -3-磷脂酸
常见甘油磷脂的极性头部和其净电荷(pH=7)
甘油磷脂名称
磷脂酸 磷脂酰乙醇胺 磷脂酰胆碱 磷脂酰丝氨酸
HO-X的名称
—— 胆胺 胆碱 丝氨酸
—X的结构
极性头基净电荷
磷脂酰甘油
甘油
磷脂酰肌醇
肌醇
H -1
HH
心磷脂
双磷脂酰甘油
例题:中性pH下,净电荷为零的 甘油磷脂是( )( )。
然而,催化加氢是一个可逆反应,饱和脂肪酸在 反应过程中,也会脱氢生成不饱和脂肪酸。这样,脱 氢的产物就可能有两种,顺式和反式。
反式不饱和脂肪酸比顺式不饱和脂肪酸空阻小,更 稳定,更容易生成,而且一旦生成,又不易被氢化饱 和。
所以,在顺式不饱和脂肪酸催化加氢的产物饱和脂 肪酸中,会含有一定量的反式不饱和脂肪酸。这就是 反式脂肪酸的由来。
影响油脂自动氧化的因素 (1)油脂的脂肪酸组成
不饱和脂肪酸越多,越容易发生自动氧化。 思考:为什么家用猪油比花生油更易变“哈喇”? 因为天然植物油脂中溶有维生素E,起抗氧化作用。
28-29-脂类代谢

Beta-hydroxyR Fatty Acyl CoA
O H O S CoA
H2O in
ETS
β
NAD
(dehydrogenase)
Beta-ketoR Fatty Acyl CoA CoA-S
ation 3
NADH + H
O O S CoA
2.5 ATP 3
H out
β
H
thiolysis
Fatty Acyl CoA
R
β
α
O S CoA
Matrix Side
FAD
oxidation 1
(dehydrogenase)
alpha-beta unsat. Fatty Acyl CoA (trans)
R
H out
ETS
FADH2
O S CoA
1.5 ATP 2
H2O
hydration 2
(hydratase)
acetyl CoA
脂肪酸氧化分解时的能量释放: 脂肪酸氧化分解时的能量释放:
由于1分子FADH 可生成1 分子ATP ATP, 分子NADH NADH可生成 由于 1 分子 FADH2 可生成 1.5 分子 ATP ,1 分子 NADH 可生成 分子ATP ATP, 分子乙酰CoA 经彻底氧化分解可生成10 CoA经彻底氧化分解可生成 2.5 分子 ATP , 1 分子乙酰 CoA 经彻底氧化分解可生成 10 分子ATP。 分子ATP。 ATP 16C acid) 为例来计算, 以 16C 的软脂酸 ( palmitic acid ) 为例来计算 , 则生成 ATP的数目为: ATP的数目为: 的数目为 7次β-氧化分解产生4×7= 氧化分解产生4 8分子乙酰CoA可得10×8= 分子乙酰CoA可得10× CoA可得10 减去活化时消耗的 28分子ATP; 28分子ATP; 分子ATP 80分子ATP; 80分子ATP; 分子ATP 2分子ATP 分子ATP
O H O S CoA
H2O in
ETS
β
NAD
(dehydrogenase)
Beta-ketoR Fatty Acyl CoA CoA-S
ation 3
NADH + H
O O S CoA
2.5 ATP 3
H out
β
H
thiolysis
Fatty Acyl CoA
R
β
α
O S CoA
Matrix Side
FAD
oxidation 1
(dehydrogenase)
alpha-beta unsat. Fatty Acyl CoA (trans)
R
H out
ETS
FADH2
O S CoA
1.5 ATP 2
H2O
hydration 2
(hydratase)
acetyl CoA
脂肪酸氧化分解时的能量释放: 脂肪酸氧化分解时的能量释放:
由于1分子FADH 可生成1 分子ATP ATP, 分子NADH NADH可生成 由于 1 分子 FADH2 可生成 1.5 分子 ATP ,1 分子 NADH 可生成 分子ATP ATP, 分子乙酰CoA 经彻底氧化分解可生成10 CoA经彻底氧化分解可生成 2.5 分子 ATP , 1 分子乙酰 CoA 经彻底氧化分解可生成 10 分子ATP。 分子ATP。 ATP 16C acid) 为例来计算, 以 16C 的软脂酸 ( palmitic acid ) 为例来计算 , 则生成 ATP的数目为: ATP的数目为: 的数目为 7次β-氧化分解产生4×7= 氧化分解产生4 8分子乙酰CoA可得10×8= 分子乙酰CoA可得10× CoA可得10 减去活化时消耗的 28分子ATP; 28分子ATP; 分子ATP 80分子ATP; 80分子ATP; 分子ATP 2分子ATP 分子ATP
【生物化学】脂类的生物合成

多烯不饱和脂肪酸在厌氧细菌中基本不存在, 但在高等动植物体内含量丰富,他们是由单烯脂 肪酸继续去饱和而产生的。
由单烯脂肪酸(△9)去饱和产生的
C16:0
软脂酸 +C2延长
பைடு நூலகம்
去饱和 -2H
△9-C16:1
C18:0
硬脂酸
棕榈油酸
去饱和 -2H
+C2延长
神经酸 二十四碳烯-15-酸
△15-C24:1
+C2延长
O HOOC-CH2-C~SCOA +ADP+Pi
乙酰-CoA羧化酶(别构酶)
生物素羧化酶 羧基转移酶(转羧酶) 生物素羧基载体蛋白(BCCP)
原核生物乙酰-CoA羧化酶:上述三种蛋 白质组成复合体。
真核生物乙酰-CoA羧化酶:由两个相 同亚基组成,上述三种蛋白质位于同一 条多肽链上。
作用机制
生物素羧化酶
生物素-酶
Lys-e氨基
CO2-生物素-酶
3. 脂肪酸合酶系统
组成:
脂酰基载体蛋白(ACP-SH) 1) 乙酰-CoA:ACP转酰酶 2) 丙二酸单酰CoA:ACP转酰酶 3) β-酮酰-ACP合酶 4) β-酮酰-ACP还原酶 5) β-羟酰-ACP脱水酶 6) 烯脂酰-ACP还原酶 7) 软脂酰-ACP硫酯酶
4. 饱和脂肪酸的从头合成与β-氧化的比较
区别要点
从头合成
β-氧化
细胞内进行部位 酰基载体 转运机制 二碳单位参与或断裂形式 电子供体或受体 -羟酰基中间物的立体构型不同 对HCO3-和柠檬酸的需求 所需酶 能量需求或放出 消耗
细胞质 ACP-SH 三羧酸转运机制
丙二酸单酰ACP
NADPH+H+ D型 需要 7种 7ATP及14NADPH+H+
由单烯脂肪酸(△9)去饱和产生的
C16:0
软脂酸 +C2延长
பைடு நூலகம்
去饱和 -2H
△9-C16:1
C18:0
硬脂酸
棕榈油酸
去饱和 -2H
+C2延长
神经酸 二十四碳烯-15-酸
△15-C24:1
+C2延长
O HOOC-CH2-C~SCOA +ADP+Pi
乙酰-CoA羧化酶(别构酶)
生物素羧化酶 羧基转移酶(转羧酶) 生物素羧基载体蛋白(BCCP)
原核生物乙酰-CoA羧化酶:上述三种蛋 白质组成复合体。
真核生物乙酰-CoA羧化酶:由两个相 同亚基组成,上述三种蛋白质位于同一 条多肽链上。
作用机制
生物素羧化酶
生物素-酶
Lys-e氨基
CO2-生物素-酶
3. 脂肪酸合酶系统
组成:
脂酰基载体蛋白(ACP-SH) 1) 乙酰-CoA:ACP转酰酶 2) 丙二酸单酰CoA:ACP转酰酶 3) β-酮酰-ACP合酶 4) β-酮酰-ACP还原酶 5) β-羟酰-ACP脱水酶 6) 烯脂酰-ACP还原酶 7) 软脂酰-ACP硫酯酶
4. 饱和脂肪酸的从头合成与β-氧化的比较
区别要点
从头合成
β-氧化
细胞内进行部位 酰基载体 转运机制 二碳单位参与或断裂形式 电子供体或受体 -羟酰基中间物的立体构型不同 对HCO3-和柠檬酸的需求 所需酶 能量需求或放出 消耗
细胞质 ACP-SH 三羧酸转运机制
丙二酸单酰ACP
NADPH+H+ D型 需要 7种 7ATP及14NADPH+H+
生物化学脂类代谢[2]
![生物化学脂类代谢[2]](https://img.taocdn.com/s3/m/5fc4da38178884868762caaedd3383c4ba4cb44e.png)
四、酮体(ketone body)的生成及利用:
主要在肝脏的线粒体中生成,合成原料为乙酰CoA, HMG-CoA合酶是酮体生成的关键酶。
1.酮体的生成: p244
乙酰乙酰硫解酶
(1) 两分子乙酰CoA在乙酰乙酰CoA硫解酶的催化下,缩合生成一分子乙酰乙酰CoA。
乙酰CoA 转酰酶
β-酮酰 合酶
β-羟酰 脱水酶
丙二酰CoA转酰酶
β酮酰 还原酶
烯酰 还原酶
长链脂肪 酰硫酯酶
HS-ACP
(1)转酰基作用:(启动) 乙酰CoA + ACP-SH 乙酰ACP + CoASH 丙二酰CoA + ACP-SH 丙二酰ACP + CoASH
肝脂 (转变、加工)
食物
糖类
生酮氨基酸
组织脂
氧化
酮体
氧化
磷脂
CO2、 H2O、ATP
第二节 脂肪的分解代谢
1、定义:贮存于脂肪细胞中的甘油三酯在激素敏感脂肪酶(HSL)的催化下水解并释放出脂肪酸和甘油,供给全身各组织细胞摄取利用的过程。
一、脂肪动员
★
HSL主要受共价修饰调节。 促脂解激素:肾上腺素、去甲肾上腺素、胰高血糖素等 抗脂解激素:胰岛素、前列腺素E
2、过程:
甘油不被脂肪细胞利用,经血液输送到肝脏进行代谢。
二、甘油代谢
甘油
3-磷酸甘油
磷酸二羟丙酮
磷酸二羟丙酮→3-磷酸甘油醛→氧化或糖异生
三、脂肪酸的氧化 (p232)
1、部位: 肝脏、肌肉(主要),胞液(活化)+ 线粒体( -氧化) 2、过程:四个阶段
脂肪酸的活化:耗能 2ATP
脂酰CoA转运入线粒体:限速步骤
主要在肝脏的线粒体中生成,合成原料为乙酰CoA, HMG-CoA合酶是酮体生成的关键酶。
1.酮体的生成: p244
乙酰乙酰硫解酶
(1) 两分子乙酰CoA在乙酰乙酰CoA硫解酶的催化下,缩合生成一分子乙酰乙酰CoA。
乙酰CoA 转酰酶
β-酮酰 合酶
β-羟酰 脱水酶
丙二酰CoA转酰酶
β酮酰 还原酶
烯酰 还原酶
长链脂肪 酰硫酯酶
HS-ACP
(1)转酰基作用:(启动) 乙酰CoA + ACP-SH 乙酰ACP + CoASH 丙二酰CoA + ACP-SH 丙二酰ACP + CoASH
肝脂 (转变、加工)
食物
糖类
生酮氨基酸
组织脂
氧化
酮体
氧化
磷脂
CO2、 H2O、ATP
第二节 脂肪的分解代谢
1、定义:贮存于脂肪细胞中的甘油三酯在激素敏感脂肪酶(HSL)的催化下水解并释放出脂肪酸和甘油,供给全身各组织细胞摄取利用的过程。
一、脂肪动员
★
HSL主要受共价修饰调节。 促脂解激素:肾上腺素、去甲肾上腺素、胰高血糖素等 抗脂解激素:胰岛素、前列腺素E
2、过程:
甘油不被脂肪细胞利用,经血液输送到肝脏进行代谢。
二、甘油代谢
甘油
3-磷酸甘油
磷酸二羟丙酮
磷酸二羟丙酮→3-磷酸甘油醛→氧化或糖异生
三、脂肪酸的氧化 (p232)
1、部位: 肝脏、肌肉(主要),胞液(活化)+ 线粒体( -氧化) 2、过程:四个阶段
脂肪酸的活化:耗能 2ATP
脂酰CoA转运入线粒体:限速步骤
28 脂肪酸的分解代谢

H
Δ2 –反-烯脂酰辅酶A
烯脂酰CoA水合酶
H
RCH2C-CH2COSCoA
OH
β-羟脂酰辅酶A
β-氧化的反应过程4---脱氢
H
RCH2C-CH2COSCoA
OH
β-羟脂酰辅酶A
NAD+
羟脂酰CoA脱氢酶
O
NADH+H+
RCH2C-CH2COSCoA
β-酮脂酰辅酶A
β-氧化的反应过程5---硫解
脂酶A1、A2、C、D的作用位点如脂质一章图示,它们 广泛存在于各种类型的细胞中。
(三) 吸收
在人和动物体内,小肠可以吸收脂类的水解产物,包 括脂肪酸(70%)、甘油、β-甘油一酯以及胆碱、部分水 解的磷脂和胆固醇等。
其中甘油、单酰甘油同脂酸在小肠粘膜细胞内重新合 成三酰甘油。新合成的三酰甘油与少量磷脂和胆固醇混 合在一起,在一层脂蛋白的包裹下形成乳糜微粒,从小 肠粘膜细胞中分泌到细胞外液,进入血液,最终被组织 吸收。
ATP ADP
NAD+ NADH
甘油
α-磷酸甘油
甘油磷酸激酶
磷酸二羟丙酮
α-磷酸甘油 脱氢酶
3磷酸甘油醛
糖代谢
(一)脂肪酸的活化
合酶在催化反应中没有 ATP直接参加反应,如若ATP直 接参加反应,则是合成酶。 (这里就应当是合成酶)
RCOOH + ATP
脂肪酸
脂酰CoA合酶
RCO-AMP+PPi 脂酰AMP
Biochemistry
概述
• 脂肪酸氧化的化学反应可分为三个方面: • 一是长链脂肪酸降解为两个碳原子单元,即乙酰-CoA。 • 二是乙酰-CoA经过柠檬酸循环氧化成CO2 。 • 三是从还原的电子载体到线粒体呼吸链的电子传递。
Δ2 –反-烯脂酰辅酶A
烯脂酰CoA水合酶
H
RCH2C-CH2COSCoA
OH
β-羟脂酰辅酶A
β-氧化的反应过程4---脱氢
H
RCH2C-CH2COSCoA
OH
β-羟脂酰辅酶A
NAD+
羟脂酰CoA脱氢酶
O
NADH+H+
RCH2C-CH2COSCoA
β-酮脂酰辅酶A
β-氧化的反应过程5---硫解
脂酶A1、A2、C、D的作用位点如脂质一章图示,它们 广泛存在于各种类型的细胞中。
(三) 吸收
在人和动物体内,小肠可以吸收脂类的水解产物,包 括脂肪酸(70%)、甘油、β-甘油一酯以及胆碱、部分水 解的磷脂和胆固醇等。
其中甘油、单酰甘油同脂酸在小肠粘膜细胞内重新合 成三酰甘油。新合成的三酰甘油与少量磷脂和胆固醇混 合在一起,在一层脂蛋白的包裹下形成乳糜微粒,从小 肠粘膜细胞中分泌到细胞外液,进入血液,最终被组织 吸收。
ATP ADP
NAD+ NADH
甘油
α-磷酸甘油
甘油磷酸激酶
磷酸二羟丙酮
α-磷酸甘油 脱氢酶
3磷酸甘油醛
糖代谢
(一)脂肪酸的活化
合酶在催化反应中没有 ATP直接参加反应,如若ATP直 接参加反应,则是合成酶。 (这里就应当是合成酶)
RCOOH + ATP
脂肪酸
脂酰CoA合酶
RCO-AMP+PPi 脂酰AMP
Biochemistry
概述
• 脂肪酸氧化的化学反应可分为三个方面: • 一是长链脂肪酸降解为两个碳原子单元,即乙酰-CoA。 • 二是乙酰-CoA经过柠檬酸循环氧化成CO2 。 • 三是从还原的电子载体到线粒体呼吸链的电子传递。
脂类的生物化学

大多数的类二十烷酸是花生四烯酸的衍生物。
前列腺素类(prostaglandin),
凝血恶烷类(thromboxane) 白细胞三烯类(leucotriene)
阿司匹林(乙酰水杨酸)
天然油脂的组分
天然油脂并非一种物质组成,而是三酰甘油的混合物。
不同种类的油脂所含的脂肪酸是不相同的。
油脂的形成
• 油脂的形成:油脂由一分子甘油和三分子脂肪 酸经过逐步反应得到。反应如下:
都是由生物体产生,并能由生物体所利用(矿物油?)
例外:卵磷脂(溶于乙醚)、鞘磷脂和脑苷脂类。
四、脂类的生物学功能
1. 结构组分 2. 储存能源 3. 溶剂 4. 保温和保护 5. 其他 ——磷脂是生物膜的主要成分 ——机体的储存燃料 ——一些活性物质的溶剂 ——防寒剂和润滑剂 ——参与机体代谢调节
• 植物固醇:
谷固醇、豆固醇,比胆固醇侧链上多一个—C2H5
• 酵母固醇:
麦角固醇,紫外线照射下,可转变为维生素D2
二、类固醇
• 胆酸和胆汁酸:胆汁的重要成分,作用于脂肪代谢
胆汁酸盐,可使脂肪乳化,促进肠壁细胞对脂肪的消化吸收。
• 固醇类激素:包括肾上腺皮质激素和性激素 • 植物类固醇:强心苷(寡糖和固醇所成的糖苷)、皂素
3、氧化作用: 油脂的不饱和脂肪酸的双键氧化分解,或油脂经微生物分解 成的脂肪酸,氧化分解形成系列产物的变质过程。 • 酸败:天然油脂长期暴露在空气中,会产生酸臭味
原因是:1、油脂受空气和光照作用,不饱和脂肪酸被氧化成过氧化 物,继续分解为低级醛、酮以及羧酸,产生酸臭味。2、霉菌或脂肪酶将 油脂水解成低级脂肪酸,再生成-酮酸,其脱羧后而成低级酮类。
油 脂 的 消 化
油脂的化学性质
生物化学第29章脂类的生物合成

50nm
脂肪酸合成Ⅰ
KSase: β-酮酰ACP合酶
ACP: 酰基载体蛋成到
脂 16 碳 脂 肪 酸 即 肪 棕 榈 酸 ( 软 脂
酸)为止。
酸 合 成
Ⅲ
脂肪酸合成碳链延伸的方向
KSase
ACP
脂肪酸合酶
脂肪酸合酶(fatty acid synthase,FAS)是 含有多种酶活性的复合体,它可以催化从丙二酰 CoA到脂肪酸的全部反应。脂肪酸合酶一般是由 酰基载体蛋白(acyl carrier protein,ACP)及6 种酶活性组成,脂肪酸合成期间就是通过其羧基 固定在酰基载体蛋白的辅基上。这个辅基是CoA 的一部分,即磷酸泛酰巯基乙胺。
α-亚麻酸合成)
多不饱和脂肪酸中 双键的排列
大多数多不饱和脂肪酸中的双键是非共轭的, 少数属于共轭系统,如乌桕酸(10:2Δ 2t,4C)和 α -桐油酸(18:3Δ 9C,11t,13t)。含有共轭双键的脂 肪酸很容易发生聚合作用。
乙酰CoA羧化酶的 共价修饰调节
乙酰CoA羧化酶是脂肪酸合成的限速酶。
1.5 19.0 18.0 8.0 1.0
0 0 17.5 10.0 25.0
人髓磷脂
0.5 10.0 20.0 8.0 1.0
0 0 8.5 26.0 26.0
牛心 线粒体
0 39.0 27.0 0.5 7.0
0 22.5
0 0 3.0
大肠杆菌 细胞膜
0 0 65.0 0 0 18.0 12.0 0 0 0
戊
酸
HMG-CoA还原酶
的
合
成
Ⅱ
甲羟戊酸
HMG-CoA的去向
HMG-CoA
HMG-CoA 裂解酶
脂肪酸合成Ⅰ
KSase: β-酮酰ACP合酶
ACP: 酰基载体蛋成到
脂 16 碳 脂 肪 酸 即 肪 棕 榈 酸 ( 软 脂
酸)为止。
酸 合 成
Ⅲ
脂肪酸合成碳链延伸的方向
KSase
ACP
脂肪酸合酶
脂肪酸合酶(fatty acid synthase,FAS)是 含有多种酶活性的复合体,它可以催化从丙二酰 CoA到脂肪酸的全部反应。脂肪酸合酶一般是由 酰基载体蛋白(acyl carrier protein,ACP)及6 种酶活性组成,脂肪酸合成期间就是通过其羧基 固定在酰基载体蛋白的辅基上。这个辅基是CoA 的一部分,即磷酸泛酰巯基乙胺。
α-亚麻酸合成)
多不饱和脂肪酸中 双键的排列
大多数多不饱和脂肪酸中的双键是非共轭的, 少数属于共轭系统,如乌桕酸(10:2Δ 2t,4C)和 α -桐油酸(18:3Δ 9C,11t,13t)。含有共轭双键的脂 肪酸很容易发生聚合作用。
乙酰CoA羧化酶的 共价修饰调节
乙酰CoA羧化酶是脂肪酸合成的限速酶。
1.5 19.0 18.0 8.0 1.0
0 0 17.5 10.0 25.0
人髓磷脂
0.5 10.0 20.0 8.0 1.0
0 0 8.5 26.0 26.0
牛心 线粒体
0 39.0 27.0 0.5 7.0
0 22.5
0 0 3.0
大肠杆菌 细胞膜
0 0 65.0 0 0 18.0 12.0 0 0 0
戊
酸
HMG-CoA还原酶
的
合
成
Ⅱ
甲羟戊酸
HMG-CoA的去向
HMG-CoA
HMG-CoA 裂解酶
脂类与生物膜
由磷脂形成的双层脂膜的示意图
在水溶液中两性的磷脂分子为避免疏水部分接触 水分子而定向排列,形成脂双层结构。脂双层中, 磷脂分子的疏水基团在内部而亲水的头部在表面。
2.1.2膜蛋白 膜蛋白是膜功能的主要承担者,依据膜蛋白与脂双层相互
作用方式不同,分为外周蛋白和内在蛋白。
分布于膜的脂双层外表面,通过极性氨基酸残基以离子 键、氢键、范德华力等次级键与膜脂极性头部或与内嵌蛋 白的亲水部分结合。
传导作用是一种化学能转化为电能的过程,肌肉收缩是化 学能转化为机械能,光合作用是光能转化为化学能,呼吸 作用是将营养物质在氧化分解过程中释放的化学能转变成 另一种高能键化学能的结果。
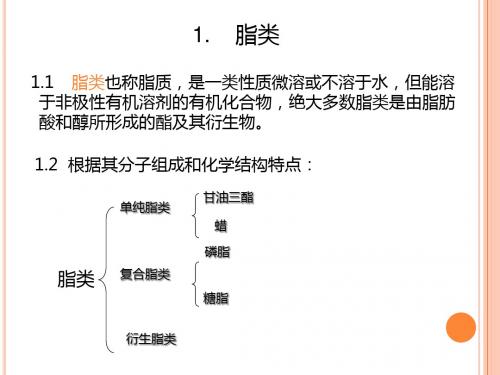
1. 脂类
1.1 脂类也称脂质,是一类性质微溶或不溶于水,但能溶 于非极性有机溶剂的有机化合物,绝大多数脂类是由脂肪 酸和醇所形成的酯及其衍生物。
1.2 根据其分子组成和化学结构特点:
单纯脂类
脂类 复合脂类
甘油三酯 蜡
磷脂
糖脂
衍生脂类
1.3脂类的生物学功能
贮存脂类——重要的贮能供能物质,每克脂肪氧化时可释放 出38.9 kJ 的能量,每克糖和蛋白质氧化时释放的能量仅分别 为17.2 kJ和23.4 kJ。蜡是海洋浮游生物体内能量物质的主要 储存形式。
1.维生素A(视黄醇)
理化性质: Vit A和胡萝卜素均耐热、酸、碱; 溶于脂肪,不溶于水,一般烹调加工不易破坏 易被氧化和被紫外线破坏; 食物中含有磷脂、Vit E、Vit C和其它抗氧化物质 时,Vit A和胡萝卜素均较稳定。
作用:
(1). 构成视觉细胞内的感光物质——视紫红质: 白天感到光刺眼,必须带墨镜;眼睛外突,
我是不是你最疼 爱的人!
2.维生素D(阳光维生素) 理化性质:
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
(二)脂肪肝 脂肪肝形成的生化机理:胰岛素欠缺;化学药品影
响
体内脂类从何而来?
为什么不进食脂肪也能长胖?其生化机 理何在?
动物体内脂类的来源
1、摄入:如脂肪进入机体后,不需要参与供能 而被贮存;
2、合成: (1)糖类代谢中间产物可转变生成脂类; (2)蛋白质代谢中间产物可转变生成; (3)核酸代谢中间产物可转变生成脂类; (4)脂类代谢中间产物亦可重新被合成脂类。
缩合
-酮脂酰-ACP合成酶
还原
-酮酯酰-ACP还原酶
四步反应延伸生长脂肪链的两 个碳(反应3、4)
-羟脂酰-ACP 脱水酶
脱水
烯酰ACP-还原酶
还原
肪
肪
肪
肪
酸
酸
氧
合
化
成
的
的
四
四
步
步
重
重
复
复
反
反
应
应
举例:软脂酸合成的全程
软脂酰-ACP硫脂酶
软脂酸合成总反应
1. 7分子丙二酸单酰CoA形成 7乙酰 CoA+7CO2+7ATP
7丙二酸单酰CoA +7ADP+7Pi 2.7次循环的缩合和还原 乙酰 CoA+7丙二酸单酰CoA+14NADPH+14H+ 软脂酸+8HS-CoA+6H2O+7CO2+14NADP+ 总反应:
8乙酰CoA+7ATP+14NADPH+14H+ 软脂酸+8HSCoA+6H2O
+7ADP+7Pi+14NADP+
7、脂肪酸合成的调节
自学: 要点: (1)乙酰-CoA羧化酶是合成过程中限速
步骤,是合成调控关键所在。 (2)柠檬酸可激活该酶,而脂酰-CoA抑
制该酶。
思考
• 为什么仅摄入糖也能长胖?阐述其生化 机理。
脂类代谢与糖类代谢的协调构通
1、脂肪可氧化提供能量; 2、糖可转变为脂肪。
(二)其它脂类的生物合成
脂酰-CoA
酰基转移酶 磷脂酸磷酸酶
1,2-二酰
三
甘油-3-磷酸
脂
酰
甘
油
的
合
成
二羟丙酮磷酸:真核
甘油:原核
2
3-磷酸甘油
、
磷
酰基转移酶 酰基转移酶
脂
的
酰基转移酶
合 成
磷脂酸
CDP-二脂酰甘油合成
磷脂酸
CDP-二脂酰甘油
CDP-二脂酰甘油可分头代谢合成其它脂类物 质
以CDP-二脂酰甘油为起点合成磷脂类
5、软脂酸合成与分解代谢的区别
?
6、脂肪酸碳链的加长和去饱和
自学: 要点: (1)胞质中脂肪酸合成停止于16碳脂肪酸即软
脂酸(其它脂肪酸的前体)。 (2)碳链延长发生于线粒体和内质网。 (3)动物体不饱和脂肪酸的形成发生于光面内
质网。 *哺乳动物缺少能够在C-9位以上引进双键的酶
→必需脂肪酸:
亚油酸(18, 9,12 ),亚麻酸(18, 9,12, 15 -型),花生四烯酸( 18, 5,8,11,14)。
草酰乙酸
CoA
苹果酸
的
苹果酸
转
丙酮酸
丙酮酸
运
P259页图29-3
2 、 丙 二 酸 单 酰
乙酰CoA羧化酶
来 源
CoA
3、脂肪酸合酶
是一个多酶复合体,包含7种酶活性和一个酰基载体蛋 白(ACP),该蛋白的辅基是磷酸泛酰巯基乙胺(含 泛酸)
7种酶活性: (1)乙酰-CoA:ACP酰基转移酶; (2)丙二酸单酰CoA:ACP酰基转移酶; (3)-酮酯酰-ACP合成酶;(缩合) (4) -酮酯酰-ACP还原酶;(还原) (5) -羟脂酰-ACP脱水酶;(脱水) (6)烯酰ACP-还原酶; (还原) (7)软脂酰-ACP硫脂酶
控作用); (3)氨基酸碳架的分解而来(补) ; (4)核苷酸的核糖分解代谢而来(补) 。
乙酰-CoA基本产生于线粒体内,线粒体内膜 对乙酰-CoA不透过,需要特殊的运输体,乙酰 -CoA通过柠檬酸合成酶与草酰乙酸生成柠檬酸 被运送到胞质。(三羧酸转运体系)
线粒体基质
细胞质
细胞质
柠檬酸
柠檬酸
乙
酰
草酰乙酸
决定反应速度的作用
习题
• 酰基载体蛋白
• 在磷脂生物合成中,起重要作用的高能化合物是:
• A ATP B GTP C CTP
• D UTP E TTP F CDP
• 乙酰基从线粒体内转运到胞液中的化合物是:
• A 乙酰辅酶A B 脂酰辅酶A C 肉碱
• D 胆碱
E 柠檬酸 F 异柠檬酸
• 脂肪酸合成的限速酶是
1、原核 CDP-二脂酰甘油 2、真核 CDP-二脂酰甘油
磷脂酰乙醇胺 磷脂酰甘油 二磷脂酰甘油
磷脂酰乙醇胺 等
磷脂酰胆碱
鞘磷脂和鞘糖脂及胆固醇的合成
自学。
* 胆固醇的合成:所有碳原子来自乙酰CoA 乙酸甲羟戊酸异戊二烯衍生物(角)鲨烯
羊毛固醇胆固醇 * HMG-CoA还原酶在胆固醇生物合成途径中起
绝大多数脂肪酸以三酰甘油或磷酸甘油 脂形式存在着。
*1、三脂酰甘油 *2、磷脂类 3、鞘磷脂和鞘糖脂 4、类十二烷酸 5、胆固醇
1、脂肪(脂酰甘油)合成
脂肪合成的两种主要前体:甘268)。
真核
原核
甘油-3-P脱氢酶
甘油激酶
甘油-3-磷酸
ACP作用:将底物在酶复合体上从一处的催化中心转移到另一处。
4、由脂肪酸合酶催化的各步反应
• 启动
与脂酸-氧化比较:
• 装载
活化
• 缩合
脱氢
• 还原 • 脱水
四个重复步骤
水化 脱氢
• 还原
硫解
• 释放
见书P236
见书P262--263
见书P262--263
合酶
脂
合酶
肪
酸
合
成
四步反应延伸生长脂肪链的两 个碳(反应1、2)
*通过怎样的途径合成?
二、脂类的生物合成
*脂肪酸是几乎所有脂质的重要组成成分。
(一)脂肪酸的生物合成 (1)场所:大多发生于细胞溶胶(植物:
叶绿体); (2)重要的原料物质:
乙酰-CoA 丙二酸单酰-CoA
1、乙酰-CoA来源
(1)葡萄糖→丙酮酸→乙酰-CoA; (2)脂肪酸-氧化生成的乙酰-CoA(取决于调
• A、柠檬酸合成酶
B、脂酰基转移酶
• C、乙酰CoA羧化酶
第29章 脂类的生物合成
脂类物质的功能: (1)贮存能量:脂肪 (2)细胞膜成分:磷脂,胆固醇 (3)特殊脂类具有的活性:维D,
维K,胆汁盐,胞内胞外信使(如 前列腺素)等
一、贮存脂肪(的动员)
(一)贮存脂肪 来自膳食的脂肪必须得先转化为贮存脂肪。需要脂 肪分解代谢提供能量ATP或用于合成其他物质, 再将其分解动员。 动员:脂肪仓库中贮存的脂肪释放出游离脂肪酸, 并转移至肝脏的过程。 需磷脂酶,脂酶的参与,见P257页。 分解产物:甘油 :在细胞溶胶中代谢 脂肪酸:线粒体中代谢
响
体内脂类从何而来?
为什么不进食脂肪也能长胖?其生化机 理何在?
动物体内脂类的来源
1、摄入:如脂肪进入机体后,不需要参与供能 而被贮存;
2、合成: (1)糖类代谢中间产物可转变生成脂类; (2)蛋白质代谢中间产物可转变生成; (3)核酸代谢中间产物可转变生成脂类; (4)脂类代谢中间产物亦可重新被合成脂类。
缩合
-酮脂酰-ACP合成酶
还原
-酮酯酰-ACP还原酶
四步反应延伸生长脂肪链的两 个碳(反应3、4)
-羟脂酰-ACP 脱水酶
脱水
烯酰ACP-还原酶
还原
肪
肪
肪
肪
酸
酸
氧
合
化
成
的
的
四
四
步
步
重
重
复
复
反
反
应
应
举例:软脂酸合成的全程
软脂酰-ACP硫脂酶
软脂酸合成总反应
1. 7分子丙二酸单酰CoA形成 7乙酰 CoA+7CO2+7ATP
7丙二酸单酰CoA +7ADP+7Pi 2.7次循环的缩合和还原 乙酰 CoA+7丙二酸单酰CoA+14NADPH+14H+ 软脂酸+8HS-CoA+6H2O+7CO2+14NADP+ 总反应:
8乙酰CoA+7ATP+14NADPH+14H+ 软脂酸+8HSCoA+6H2O
+7ADP+7Pi+14NADP+
7、脂肪酸合成的调节
自学: 要点: (1)乙酰-CoA羧化酶是合成过程中限速
步骤,是合成调控关键所在。 (2)柠檬酸可激活该酶,而脂酰-CoA抑
制该酶。
思考
• 为什么仅摄入糖也能长胖?阐述其生化 机理。
脂类代谢与糖类代谢的协调构通
1、脂肪可氧化提供能量; 2、糖可转变为脂肪。
(二)其它脂类的生物合成
脂酰-CoA
酰基转移酶 磷脂酸磷酸酶
1,2-二酰
三
甘油-3-磷酸
脂
酰
甘
油
的
合
成
二羟丙酮磷酸:真核
甘油:原核
2
3-磷酸甘油
、
磷
酰基转移酶 酰基转移酶
脂
的
酰基转移酶
合 成
磷脂酸
CDP-二脂酰甘油合成
磷脂酸
CDP-二脂酰甘油
CDP-二脂酰甘油可分头代谢合成其它脂类物 质
以CDP-二脂酰甘油为起点合成磷脂类
5、软脂酸合成与分解代谢的区别
?
6、脂肪酸碳链的加长和去饱和
自学: 要点: (1)胞质中脂肪酸合成停止于16碳脂肪酸即软
脂酸(其它脂肪酸的前体)。 (2)碳链延长发生于线粒体和内质网。 (3)动物体不饱和脂肪酸的形成发生于光面内
质网。 *哺乳动物缺少能够在C-9位以上引进双键的酶
→必需脂肪酸:
亚油酸(18, 9,12 ),亚麻酸(18, 9,12, 15 -型),花生四烯酸( 18, 5,8,11,14)。
草酰乙酸
CoA
苹果酸
的
苹果酸
转
丙酮酸
丙酮酸
运
P259页图29-3
2 、 丙 二 酸 单 酰
乙酰CoA羧化酶
来 源
CoA
3、脂肪酸合酶
是一个多酶复合体,包含7种酶活性和一个酰基载体蛋 白(ACP),该蛋白的辅基是磷酸泛酰巯基乙胺(含 泛酸)
7种酶活性: (1)乙酰-CoA:ACP酰基转移酶; (2)丙二酸单酰CoA:ACP酰基转移酶; (3)-酮酯酰-ACP合成酶;(缩合) (4) -酮酯酰-ACP还原酶;(还原) (5) -羟脂酰-ACP脱水酶;(脱水) (6)烯酰ACP-还原酶; (还原) (7)软脂酰-ACP硫脂酶
控作用); (3)氨基酸碳架的分解而来(补) ; (4)核苷酸的核糖分解代谢而来(补) 。
乙酰-CoA基本产生于线粒体内,线粒体内膜 对乙酰-CoA不透过,需要特殊的运输体,乙酰 -CoA通过柠檬酸合成酶与草酰乙酸生成柠檬酸 被运送到胞质。(三羧酸转运体系)
线粒体基质
细胞质
细胞质
柠檬酸
柠檬酸
乙
酰
草酰乙酸
决定反应速度的作用
习题
• 酰基载体蛋白
• 在磷脂生物合成中,起重要作用的高能化合物是:
• A ATP B GTP C CTP
• D UTP E TTP F CDP
• 乙酰基从线粒体内转运到胞液中的化合物是:
• A 乙酰辅酶A B 脂酰辅酶A C 肉碱
• D 胆碱
E 柠檬酸 F 异柠檬酸
• 脂肪酸合成的限速酶是
1、原核 CDP-二脂酰甘油 2、真核 CDP-二脂酰甘油
磷脂酰乙醇胺 磷脂酰甘油 二磷脂酰甘油
磷脂酰乙醇胺 等
磷脂酰胆碱
鞘磷脂和鞘糖脂及胆固醇的合成
自学。
* 胆固醇的合成:所有碳原子来自乙酰CoA 乙酸甲羟戊酸异戊二烯衍生物(角)鲨烯
羊毛固醇胆固醇 * HMG-CoA还原酶在胆固醇生物合成途径中起
绝大多数脂肪酸以三酰甘油或磷酸甘油 脂形式存在着。
*1、三脂酰甘油 *2、磷脂类 3、鞘磷脂和鞘糖脂 4、类十二烷酸 5、胆固醇
1、脂肪(脂酰甘油)合成
脂肪合成的两种主要前体:甘268)。
真核
原核
甘油-3-P脱氢酶
甘油激酶
甘油-3-磷酸
ACP作用:将底物在酶复合体上从一处的催化中心转移到另一处。
4、由脂肪酸合酶催化的各步反应
• 启动
与脂酸-氧化比较:
• 装载
活化
• 缩合
脱氢
• 还原 • 脱水
四个重复步骤
水化 脱氢
• 还原
硫解
• 释放
见书P236
见书P262--263
见书P262--263
合酶
脂
合酶
肪
酸
合
成
四步反应延伸生长脂肪链的两 个碳(反应1、2)
*通过怎样的途径合成?
二、脂类的生物合成
*脂肪酸是几乎所有脂质的重要组成成分。
(一)脂肪酸的生物合成 (1)场所:大多发生于细胞溶胶(植物:
叶绿体); (2)重要的原料物质:
乙酰-CoA 丙二酸单酰-CoA
1、乙酰-CoA来源
(1)葡萄糖→丙酮酸→乙酰-CoA; (2)脂肪酸-氧化生成的乙酰-CoA(取决于调
• A、柠檬酸合成酶
B、脂酰基转移酶
• C、乙酰CoA羧化酶
第29章 脂类的生物合成
脂类物质的功能: (1)贮存能量:脂肪 (2)细胞膜成分:磷脂,胆固醇 (3)特殊脂类具有的活性:维D,
维K,胆汁盐,胞内胞外信使(如 前列腺素)等
一、贮存脂肪(的动员)
(一)贮存脂肪 来自膳食的脂肪必须得先转化为贮存脂肪。需要脂 肪分解代谢提供能量ATP或用于合成其他物质, 再将其分解动员。 动员:脂肪仓库中贮存的脂肪释放出游离脂肪酸, 并转移至肝脏的过程。 需磷脂酶,脂酶的参与,见P257页。 分解产物:甘油 :在细胞溶胶中代谢 脂肪酸:线粒体中代谢