病毒的遗传分析
合集下载
10细菌和病毒的遗传-性导、转导

如果研究三因子转导(three-factor transduction),只需分析一个实 验的结果就可以推出三个基因的次序。
普遍性转导
例如:供体基因型a+b+c+,受体的基因型为a- b- c- 。 供体用P1噬菌体感染,P1的后代再用来感染受体细胞,
然后把受体细胞接种在选择培养基上。
如果通过中断杂交已知三个基因中的一个如a不在中 间,就可对a+进行选择,即在对a+进行选择的选择培 养基上,把可以生长的a+细胞选出来。然后,再把被 选择的受体细胞重复接种在其他对b+或c+进行选择的 选择培养基上,检查a+细胞是否同时具有b+和c+。
突变子和重组子都是一个核苷酸对或者碱基对(bp)。所
以基因内每个碱基均可能发生突变,任意两个碱基间均能 发生交换重组
噬菌体突变型的互补试验
属于同一基因(功能单位)还是两个基因突变产生的呢
p59
对于两个独立起源的、表型相似的隐性突变,如何判定是 在二倍体生物中,可以建立双突变杂合体。双突变体杂合 体有两种形式:顺式(cis)和反式(trans)
普遍性转导
最少的一类转导体应当代表最难于转导的情况,
这种转导体是同时发生交换次数最多的一类。
这种转导子的基因排列应为两边是供体基因,而
中间为受体基因。
假定由实验得到的最少的转导体类别为a+b+c- ,
那么就可以确定,这三个基因的正确次序应当是 acb或bca,而不是abc。
普遍性转导
如λ的DNA,既可以以自主的状态存在,也可以整合在细菌染色 体中。这种有两种状态的遗传因子叫做附加体(episome)。
细菌和病毒的遗传学分析

gal
用不同的Hfr菌株进行中断杂交实验所作出的大肠杆菌基因连锁图,其基因向F-细胞转移的顺序大不相同。
重组作图
01
当转移时间间隔在两分钟之内, 如已知lac与ade紧密连锁,距离约为1分钟,中断杂交作图就不可靠,须用传统的重组作图(recombination mapping)
01
不用亲本类型 两对基因间的交换频率,必须在形成部分二倍体的条件下,计算重组率。 部分二倍体如果不发生重组,无法鉴别。 接合重组不产生相反的重组类型
低频重组与高频重组
高频重组(High frequence recombination, Hfr)
F因子整合到了细菌染色体上,与F-细胞接合后将供体染色体的一部分或全部传递给F-受体,当供体和受体的等位基因带有不同的遗传标记时,可观察到它们之间发生重组,频率可达到10-2以上,称为高频重组品系(菌株)
杂合DNA复制后,形成一个亲代类型的DNA和一个重组类型的DNA并导致转化细胞的形成与表达。
转化的进程
4 共转化与遗传图谱绘制
共转化:供体的一条DNA片段上的两个基因同时转换的现象。 利用共同转化绘制细菌连锁遗传图谱的基本原理: 相邻基因发生共同转化的概率与两者的距离间成正向关系,基因间距离越近,发生共同转化的频率越高,反之越低。 因此可能通过测定两基因共同转化的频率来指示基因间的相对距离。
数理与生物工程学院
单击添加副标题
遗 传 学
单击添加副标题
第七章细菌和病毒的遗传学分析
目录
1
2
二 细菌的接合与染色体作图
1.接合现象的发现
细菌的接合首先是莱德伯格( Lederberg )和塔特姆( Tatum )在1946大肠杆菌杂交试验中发现的。
用不同的Hfr菌株进行中断杂交实验所作出的大肠杆菌基因连锁图,其基因向F-细胞转移的顺序大不相同。
重组作图
01
当转移时间间隔在两分钟之内, 如已知lac与ade紧密连锁,距离约为1分钟,中断杂交作图就不可靠,须用传统的重组作图(recombination mapping)
01
不用亲本类型 两对基因间的交换频率,必须在形成部分二倍体的条件下,计算重组率。 部分二倍体如果不发生重组,无法鉴别。 接合重组不产生相反的重组类型
低频重组与高频重组
高频重组(High frequence recombination, Hfr)
F因子整合到了细菌染色体上,与F-细胞接合后将供体染色体的一部分或全部传递给F-受体,当供体和受体的等位基因带有不同的遗传标记时,可观察到它们之间发生重组,频率可达到10-2以上,称为高频重组品系(菌株)
杂合DNA复制后,形成一个亲代类型的DNA和一个重组类型的DNA并导致转化细胞的形成与表达。
转化的进程
4 共转化与遗传图谱绘制
共转化:供体的一条DNA片段上的两个基因同时转换的现象。 利用共同转化绘制细菌连锁遗传图谱的基本原理: 相邻基因发生共同转化的概率与两者的距离间成正向关系,基因间距离越近,发生共同转化的频率越高,反之越低。 因此可能通过测定两基因共同转化的频率来指示基因间的相对距离。
数理与生物工程学院
单击添加副标题
遗 传 学
单击添加副标题
第七章细菌和病毒的遗传学分析
目录
1
2
二 细菌的接合与染色体作图
1.接合现象的发现
细菌的接合首先是莱德伯格( Lederberg )和塔特姆( Tatum )在1946大肠杆菌杂交试验中发现的。
医学-第四章病毒遗传分析

终止 5`UAG 3` 3`AUC 5` 酪氨酸
(2)噬菌体的抑制因子敏感突变型类型及表现 琥珀型(amber)UAG 赭石型(ocher)UAA 乳白型(opal) UGA
表5-2携带不同专一性抑制基因宿主中sus突变噬菌体的表现
(四)无义突变与无义抑制突变
无义突变:指一个为氨基酸编码的密码变为终止密码 的突变。
• Benzer所用 T4的 r II突变就是遗传学研究 中所用的第一个条件致死突变型。
• T4噬菌体有多个迅速裂解突变型,分别称为 rl, r II,rIII等,它们位于 T4染色体 DNA的不同区 段,这 3组突变型由于在大肠杆菌不同菌株上的 反应不同可以相互区别。
• T4 r II突变使所侵染细胞迅速裂解形成大噬菌 斑,所以称为 r II突变型。
(1)抑制因子敏感突变的概念: 例如:噬菌体mRNA基因 细菌tRNA基因反密码子
正常 突变
突变
正常
基因:5`TAC 3`5`TAG 3` 3`ATC 5`3`ATG 5`
mRNA 5`UAC 3` 5`UAG 3` 3`AUC 5` 3`AUG 5`
酪氨酸 酪氨酸
表型:酪氨酸
噬菌体生长的测定——一步生长曲线
一步生长曲线:定量描述一群菌体内毒性噬菌体生 长规律的实验曲线。
感染后培养过程
被噬菌体侵染的
菌群培养过程
定时取培养 液与敏
感菌混合平 板培养
不同取样时间培养液与敏感菌混合 平板培养产生的噬菌斑数量的动态
• 四、基本术语 • 1.涂布效率(e.o.p)=噬菌斑/感染噬菌体颗
2.1 互补测验原理和方法 基础遗传学研究首先须有突变型,然后
第六章 细菌和病毒的遗传作图

15
第15页,共134页。
温和噬菌体(temperate phages)的 感染周期:
λ和P1噬菌体 溶源途径Lysogenic Cycle: 裂解途径Lytic Cycle
l
16
第16页,共134页。
Temperate Bacteriophage Lifecycle
• 能够溶源化细菌的噬菌体称为温和噬菌体
第1页,共134页。
第6章 细菌及其病毒的遗传作图
第一节 噬菌体遗传分析 第二节 细菌的转化 第三节 细菌的接合
第四节 细菌的性导
第五节 转导
2
第2页,共134页。
第一节 噬菌体遗传分析
一、噬菌体的繁殖 二、噬菌体的突变型 三、噬菌体的基因重组 四、T2的环形遗传图
3
第3页,共134页。
复习1:病毒的一般特性和类型
• 最大的病毒: 牛痘苗病毒——直径超过250nm
• 最小的病毒 脊髓灰质炎病毒——28nm
• 直径 病毒:细菌:真菌 = 1:10:100
5
第5页,共134页。
病毒粒子的模式构造
图6-1
6
第6页,共134页。
复习2:噬菌体的一般特性
• 病毒可根据宿主(动物、植物、细菌)或遗传物 质(DNA或RNA)来分类。
二、转化过程
1.供体(donor)DNA与受体(receptor)细胞 结合(binding)——吸附
– 结合发生在受体细胞特定部位; – 供体DNA片段为双链; – 结合过程是一个可逆过程。
42
第42页,共134页。
感受态与感受态因子
二、转化过程
感受态指细菌能够从周围环境中吸收DNA分子进行 转化的生理状态。
31
第15页,共134页。
温和噬菌体(temperate phages)的 感染周期:
λ和P1噬菌体 溶源途径Lysogenic Cycle: 裂解途径Lytic Cycle
l
16
第16页,共134页。
Temperate Bacteriophage Lifecycle
• 能够溶源化细菌的噬菌体称为温和噬菌体
第1页,共134页。
第6章 细菌及其病毒的遗传作图
第一节 噬菌体遗传分析 第二节 细菌的转化 第三节 细菌的接合
第四节 细菌的性导
第五节 转导
2
第2页,共134页。
第一节 噬菌体遗传分析
一、噬菌体的繁殖 二、噬菌体的突变型 三、噬菌体的基因重组 四、T2的环形遗传图
3
第3页,共134页。
复习1:病毒的一般特性和类型
• 最大的病毒: 牛痘苗病毒——直径超过250nm
• 最小的病毒 脊髓灰质炎病毒——28nm
• 直径 病毒:细菌:真菌 = 1:10:100
5
第5页,共134页。
病毒粒子的模式构造
图6-1
6
第6页,共134页。
复习2:噬菌体的一般特性
• 病毒可根据宿主(动物、植物、细菌)或遗传物 质(DNA或RNA)来分类。
二、转化过程
1.供体(donor)DNA与受体(receptor)细胞 结合(binding)——吸附
– 结合发生在受体细胞特定部位; – 供体DNA片段为双链; – 结合过程是一个可逆过程。
42
第42页,共134页。
感受态与感受态因子
二、转化过程
感受态指细菌能够从周围环境中吸收DNA分子进行 转化的生理状态。
31
第六章 病毒的遗传分析

三个基因的染色体图
m 12.9cM r 20.8cM tu
一、互补测验和顺反子 二、X174条件致死突变型的互补 三、T4突变型的互补试验
一、互补测验与顺反子
❖ 互补测验:是确定突变的功能关系。(重组测验是 确定突变基因的空间关系)
❖ 根据顺式排列和反式排列的表现型来确定两个突变 体是否属于同一个基因(顺反子)。
无基盘,即尾部不完全
无头
有头部、 尾部、尾 丝,但不 装配
说明基因 13一定控 制着头部 装配过程 中的某一 步骤。
实验结果
❖ 结果发现:rⅡ突变型可分成rⅡA和rⅡB两个互补群。 ❖ 所有rⅡA突变型的突变位点都在rⅡ区的一头,所有
rⅡB突变型的突变位点都在rⅡ区的另一头。
凡是属于rⅡA互补群的突变不能互补,同理属于B互 补群的突变也不能互补;它们均是一个独立的功能单 位,在大肠杆菌K(λ)中增殖需这两种功能。
主要内容
• 病毒的形态结构与基因组 • 噬菌体的繁殖和突变型 • 噬菌体突变型的重组实验 • 噬菌体突变型的互补测验 • λ噬菌体基因组与位点专一性重组
❖ 病毒:专性寄生的非细胞生物,形态多种多样。 结构十分简单,只有一条染色体,由蛋白质外 壳及其包被的核酸所组成。
❖ 病毒的基因组:DNA或RNA, ss或ds,+或-, L或O型。
能互补
+ rII1 + rII2 顺式(cis)排列
能互补
2个突变点在同一基因内
反式排列,不能互补
顺式排列,能互补
Benzer就将这样一个不同突变之间没有互补的功能 区域称为顺反子(cristron) 。
2、突变子(muton):是指一个顺反子内部能发 生突变的最小单位。最小的突变子只有1对碱 基,本身无独立的功能。
细菌及病毒的遗传分析h

trp2+ his2+ tyr1+转化trp2- his2- tyr1- 实验 trp2 34 his2 13 tyr1
Hfr菌株在切除F因子时发生错误切除,分离出一个携带F因子和部分宿主染色体基因的遗传因子,这种带有宿主染色体基因的F因子称为F΄因子。
T2噬菌体的基因重组
将两种不同的T2突变体进行杂交,对其杂交子代进行重组分析 杂交方法: 将Ttor和Ttos两种大肠杆菌细胞混合 同时接种高浓度的T2噬菌体的h-r+和h+r-两种突变体,保证绝大多数细菌都被一个以上噬菌体感染 两种不同的噬菌体DNA可能在宿主细胞内进行重组,从而产生非亲本型子代h+r+和h-r-。 亲本型 重组型
F因子在杂交中的行为——接合过程
(三)中断杂交实验作图
中断杂交实验作图
1分钟≈20%的重组值
二、转化
转化(transformation):指某些细菌(或其它生物)能通过其细胞膜摄取周围介质中的DNA片段,并将此外源DNA片段整合到自己染色体组中的过程。 (一)转化的过程 非感受态细胞 外源DNA被洗掉了 转化因子 感受态细胞 外源DNA仍与细胞结合 整合 吸收 整合 供体单链DNA进入受体细胞后与受体染色体的某一部分联会,并进一步置换受体的对应染色体区段的过程。
第十章 细菌及病毒的遗传分析(2h)
1
第一节 细菌和病毒遗传研究的意义
2
第二节 噬菌体的基因重组
3
第三节 细菌基因重组
4
本章要求
5
思考题
繁殖世代所需时间短;
易于管理和进行化学分析;
便于研究基因的作用;
便于研究基因的突变;
遗传物质较简单,便于用作研究基因结构、功能及调控机制的材料。
遗传学_ 细菌和病毒的遗传分析_
1180 + 418 + 685 +107 +11940 +3660
100% = 2390 100% =13% 17990
trp2
tyr
34
his2
13 tyr1
his
40
trp
八、转导(transduction)
⚫ 普遍性转导(Generalized transduction)
转导是以噬菌 体为媒介,将 外源基因携带 入细菌,使受 体细胞发生遗 传重组的方式。
a、b间发生交换
单性状的转化子
a、b间不发生交换
双性状的转化子
七、转化作图的原理
细菌两连锁基因的交换率
=
单性状转化子的数 单性状转化子数+共转化的转化子数
100%
表7-1 枯草芽孢杆菌trp2+ his2+ tyr1+(供体)× trp2- his2- tyr1-(受体)的转化实验 座位转化子类型
噬菌体的遗传分析
一、细菌和病毒的遗传分析
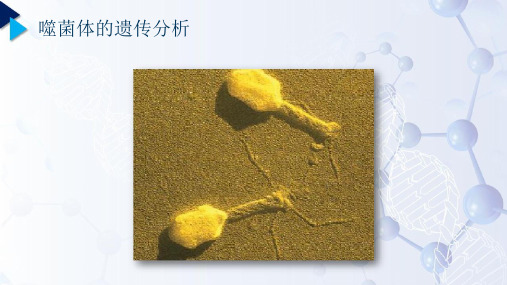
7-1 T4噬菌体的电镜照片
二、病毒对遗传学研究的贡献
1952年 Hershey & Chase的同位素示踪试验
证明T4病毒的遗传物质 是脱氧核糖核酸(DNA) 【1969年诺贝尔奖】
二、病毒对遗传学研究的贡献
1956年Fraemkel Conrat的烟草花叶病毒的重建试验
滑,可致病)
粗糙型R菌株 (无荚膜,菌落粗
糙,不致病)
三、转化现象的发现——Griffth的肺炎双球菌实验
IIR菌株不致病 IIIS菌株致病
灭活的IIIS菌株不致病 灭活的IIIS菌株的某种物 质使IIR菌株发生性状改 变,变成致病的IIIS菌株
红眼病病毒遗传变异分析

红眼病病毒遗传变异分析红眼病是一种常见的传染病,也称为流行性感冒性结膜炎。
该病毒属于腺病毒科,是一种DNA病毒,具有外壳和一个核心。
在不同地区和时间,红眼病病毒会发生不同的遗传变异,这对于疫苗和药物的开发具有重要的意义。
遗传结构和变异红眼病病毒的遗传结构包括两个重要的特征:一个是病毒壳体蛋白,另一个是免疫原性毒素蛋白。
这两个特征决定了病毒的侵入性和致病力。
红眼病病毒具有高度的变异能力,这是由其特殊的复制方式所决定的。
在感染人体时,病毒会在基因组复制的过程中频繁发生错误,这导致了基因序列的随机变化。
如果累积的错误达到一定数量,会引起突变。
突变后,病毒的构成和性质就会发生变化,甚至会产生新的亚型。
遗传变异的影响红眼病病毒的遗传变异对临床治疗和预防具有影响。
当病毒发生变异时,可能会导致对治疗药物和疫苗的抵抗性增强。
因此,红眼病病毒的遗传变异,使得对其进行疫苗和药物的研发变得更为困难。
研究方法和发展为了更好地研究红眼病病毒的遗传变异,科学家们使用了许多不同的技术手段。
其中,分子病毒学是一种比较先进的方法。
利用这种方法,可以对病毒基因组进行深入的分析,以确定遗传差别和变异点。
同时,该方法还可以帮助科学家确定红眼病病毒的遗传进化路径,从而为疫苗和药物的开发提供基础数据。
总结红眼病是一种非常常见的传染病,对人体健康造成较大的威胁。
由于红眼病病毒的遗传变异,使得治疗和预防的难度加大。
科学家们利用现有的技术手段,对病毒进行研究,不断探索其遗传结构和变异情况。
相信在今后的科学研究中,我们能够找到更好的治疗和预防方法,促进人类健康事业的发展。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
拟有性过程: 不经过减数分裂和授精作用而 导致遗传物质的重组的过程,是细菌或病 毒获取外源遗传物质的方式或途径。
三.病毒的形态结构与基因组 病毒的形态结构
病毒的基因组特点
①病毒的基因组大小相差较大,类型多样; ②多数RNA病毒的基因组是由连续的核糖核 酸链组成,但也有些病毒的基因组RNA由不 连续的几条核酸链组成; ③基因重叠; ④冗余序列少或没有; ⑤基因为多顺反子; ⑥绝大多数基因组都是单倍体; ⑦噬菌体(细胞病毒)的基因是连续的;而 真核细胞病毒的基因是不连续的。
噬菌体重组值的计算
重组噬菌斑数
重组值 = 总噬菌斑数
*100%
杂交组合
ra-h+×ra+hrb-h+×rb+hrc-h+×rc+h-
rxh+与r+h-杂交结果
每种基因型的%
h+r-
h-r+
h+r+
h-r-
34 42 12 12
32 56 5.9 6.4
39 59 0.7 0.9
重组值
24/100=24% 12.3/100=12.3% 1.6/99.6=1.6%
根据上表结果可以分别作出3个连锁图
有四种可能的排列顺序
????
你能否确定这4个基因座的关系?还 缺什么条件?怎么解决?为什么?
三.λ噬菌体的基因重组与作图
Kaiser(1955),λ噬菌体的重组作图。 用UV照射获得了5个λ噬菌体的突变型:
s型,噬菌斑较小, mi型,噬菌斑特别小, c型,噬菌斑完全清晰, co1型,噬菌斑中间模糊,四周清晰, co2型,噬菌斑的中部较co1型更为浓密。
Benzer重组实验:r47 +和+ r104
• 利用这种方法测得最小重组率的意义 最小测得重组率=0.02% =0.02 cM T4噬菌体图距=1500 cM (遗传重组测得) T4噬菌体1.8 105 bp(生物化学测得) 回头看作图的精度:
0.02cM=2-3bp = [1.8 105bp /1500cM] 0.02cM
wa w × wa Y(杏色)
F2 wa wa wa w wa Y wY
杏色
白色
在大量的F2群体中却出现了1/1000野生型 红眼出现
P wa+/ wa+ × +w/ Y
F1
wa+/ +w × wa+/ Y
(雌配子) wa+ +w waw ++
(雄配子) wa+ Y
wa+ +w waw ++ wa+ +w waw ++ wa+ wa+ wa+ wa+ Y Y Y Y
wa+/ +w两个突变分别在两条染色体上,称 为反式(trans), wa w /++两个突变同时排 在一条染色体上,而另一条染色体上两个位 点均正常,称为顺式(cis)。反式表现为突 变型,顺式排列为野生型,这种由于排列方 式不同而表型不同的现象称为顺反位置效应 (cis-trans position effect) 。
二.T2突变型的两点测交与作图
T2突变型及特性
快速溶菌突变型: 野生型: T2宿主范围野生型: T2宿主范围突变型:
细菌
B r 大斑 r+ 小斑 h+ +
h+
B2 大斑 小斑
-
+
B+B2 大斑 小斑 半透明 透明
表现型 透明,小 半透明,大 透明,大 半透明,小
基因型 hr+ 亲组合 h+r 亲组合 hr 重组合 h+r+ 重组合
利用互补测验确定顺式排列或反式时能否发生 功能互补,顺式排列能发生功能互补且反式排 列也能互补,属两个顺反子(两个基因)。顺 式排列能发生功能互补且反式排列不能互补, 是属一个顺反子(一个基因)。
斑 点 测 试 法 ( )
spot test
顺反子:Benzer把在反式构型中不能互补的各 个突变位点在染色体上所占的一个区域称为一 个顺反子。一个顺反子就是一个基因,就是一 个基因座位,包含多个基因位点,是遗传上的 一个功能单位,但不是一个突变单位或重组单 位。
四.T4突变型的三点测交与作图
T4的 m r tu + + + 三点试验结果
亲本类型 单交换型 单交换型 双交换型
合计
类 型 噬菌斑数 %
重组频率%
m-r r-tu m-tu
m r tu +++
3467 3279
69.6%
m++ + r tu
520 474
9.6% ∨
∨
mr+ + + tu
853 965
17.5%
∨∨
m + tu 162 + r +
10342
3.3% ∨ ∨ 12.9 20.8 27.1
作图: m 12.9 r 20.8 tu
第三节 噬菌体突变型的互补测验
一.互补测验与顺反子
1. 拟等位现 P
象和顺反位置
效应
F1
wa wa × wY 杏色 白色
第一节 噬菌体的遗传基础
一.病毒遗传研究的意义和优越性
① 世代周期短,繁殖块,繁殖系数高。 ② 易于管理和进行生物化学分析。 ③ 遗传物质比较简单,用于研究基因结构 、功能及表达调控机制比较方便。 ④ 便于研究基因的突变。 ⑤ 便于研究基因的作用。 ⑥ 可用作研究高等生物的简单模型。
二.病毒的拟有性过程
四.病毒的增殖与突变型 增 殖
烈 性 噬 菌 体 生 活 周 期
温和噬菌体生活周期
突变型
条件致死突变型:在某些条件下,这些突变型
是致死的,而在另一些条件下仍可进E. 行Co繁li B殖+B,2
从而得以扩增进行研究。常用的有两大类条件
致死突变:“抑制因子敏感”突变和“温度敏
感”突变。 噬菌斑形态突变型:
E.Coli B
宿主范 围突变 型:
T4 野生型和突变型的区别
类型 不同大肠杆菌菌斑平板上表型
野生型 rI r II
r III
B
K(λ)
S
小噬菌斑 小噬菌斑 小噬菌斑
大噬菌斑 大噬菌斑 大噬菌斑
大噬菌斑 无噬菌斑 小噬菌斑 (致死)
小噬菌斑 大噬菌斑 大噬菌斑
第二节 噬菌体突变型的重组测验
一.Benzer的重组测验与基因的精细结构分 析
拟等位基因(pseudoalleles):表型效应相似, 功能密切相关,在染色体上的位置又紧密连 锁的基因。这些基因产生的表型相似,但位 置上并不等位。
2. 互补测验原理与方法
互补测验(complementation test):也称顺 反测验(cis-trans test),指将两个突变分别 处于顺式和反式, 根据其表型确定两个突变 是否是同一基因的试验(看顺式反式排列是 否有功能上的互补)。
三.病毒的形态结构与基因组 病毒的形态结构
病毒的基因组特点
①病毒的基因组大小相差较大,类型多样; ②多数RNA病毒的基因组是由连续的核糖核 酸链组成,但也有些病毒的基因组RNA由不 连续的几条核酸链组成; ③基因重叠; ④冗余序列少或没有; ⑤基因为多顺反子; ⑥绝大多数基因组都是单倍体; ⑦噬菌体(细胞病毒)的基因是连续的;而 真核细胞病毒的基因是不连续的。
噬菌体重组值的计算
重组噬菌斑数
重组值 = 总噬菌斑数
*100%
杂交组合
ra-h+×ra+hrb-h+×rb+hrc-h+×rc+h-
rxh+与r+h-杂交结果
每种基因型的%
h+r-
h-r+
h+r+
h-r-
34 42 12 12
32 56 5.9 6.4
39 59 0.7 0.9
重组值
24/100=24% 12.3/100=12.3% 1.6/99.6=1.6%
根据上表结果可以分别作出3个连锁图
有四种可能的排列顺序
????
你能否确定这4个基因座的关系?还 缺什么条件?怎么解决?为什么?
三.λ噬菌体的基因重组与作图
Kaiser(1955),λ噬菌体的重组作图。 用UV照射获得了5个λ噬菌体的突变型:
s型,噬菌斑较小, mi型,噬菌斑特别小, c型,噬菌斑完全清晰, co1型,噬菌斑中间模糊,四周清晰, co2型,噬菌斑的中部较co1型更为浓密。
Benzer重组实验:r47 +和+ r104
• 利用这种方法测得最小重组率的意义 最小测得重组率=0.02% =0.02 cM T4噬菌体图距=1500 cM (遗传重组测得) T4噬菌体1.8 105 bp(生物化学测得) 回头看作图的精度:
0.02cM=2-3bp = [1.8 105bp /1500cM] 0.02cM
wa w × wa Y(杏色)
F2 wa wa wa w wa Y wY
杏色
白色
在大量的F2群体中却出现了1/1000野生型 红眼出现
P wa+/ wa+ × +w/ Y
F1
wa+/ +w × wa+/ Y
(雌配子) wa+ +w waw ++
(雄配子) wa+ Y
wa+ +w waw ++ wa+ +w waw ++ wa+ wa+ wa+ wa+ Y Y Y Y
wa+/ +w两个突变分别在两条染色体上,称 为反式(trans), wa w /++两个突变同时排 在一条染色体上,而另一条染色体上两个位 点均正常,称为顺式(cis)。反式表现为突 变型,顺式排列为野生型,这种由于排列方 式不同而表型不同的现象称为顺反位置效应 (cis-trans position effect) 。
二.T2突变型的两点测交与作图
T2突变型及特性
快速溶菌突变型: 野生型: T2宿主范围野生型: T2宿主范围突变型:
细菌
B r 大斑 r+ 小斑 h+ +
h+
B2 大斑 小斑
-
+
B+B2 大斑 小斑 半透明 透明
表现型 透明,小 半透明,大 透明,大 半透明,小
基因型 hr+ 亲组合 h+r 亲组合 hr 重组合 h+r+ 重组合
利用互补测验确定顺式排列或反式时能否发生 功能互补,顺式排列能发生功能互补且反式排 列也能互补,属两个顺反子(两个基因)。顺 式排列能发生功能互补且反式排列不能互补, 是属一个顺反子(一个基因)。
斑 点 测 试 法 ( )
spot test
顺反子:Benzer把在反式构型中不能互补的各 个突变位点在染色体上所占的一个区域称为一 个顺反子。一个顺反子就是一个基因,就是一 个基因座位,包含多个基因位点,是遗传上的 一个功能单位,但不是一个突变单位或重组单 位。
四.T4突变型的三点测交与作图
T4的 m r tu + + + 三点试验结果
亲本类型 单交换型 单交换型 双交换型
合计
类 型 噬菌斑数 %
重组频率%
m-r r-tu m-tu
m r tu +++
3467 3279
69.6%
m++ + r tu
520 474
9.6% ∨
∨
mr+ + + tu
853 965
17.5%
∨∨
m + tu 162 + r +
10342
3.3% ∨ ∨ 12.9 20.8 27.1
作图: m 12.9 r 20.8 tu
第三节 噬菌体突变型的互补测验
一.互补测验与顺反子
1. 拟等位现 P
象和顺反位置
效应
F1
wa wa × wY 杏色 白色
第一节 噬菌体的遗传基础
一.病毒遗传研究的意义和优越性
① 世代周期短,繁殖块,繁殖系数高。 ② 易于管理和进行生物化学分析。 ③ 遗传物质比较简单,用于研究基因结构 、功能及表达调控机制比较方便。 ④ 便于研究基因的突变。 ⑤ 便于研究基因的作用。 ⑥ 可用作研究高等生物的简单模型。
二.病毒的拟有性过程
四.病毒的增殖与突变型 增 殖
烈 性 噬 菌 体 生 活 周 期
温和噬菌体生活周期
突变型
条件致死突变型:在某些条件下,这些突变型
是致死的,而在另一些条件下仍可进E. 行Co繁li B殖+B,2
从而得以扩增进行研究。常用的有两大类条件
致死突变:“抑制因子敏感”突变和“温度敏
感”突变。 噬菌斑形态突变型:
E.Coli B
宿主范 围突变 型:
T4 野生型和突变型的区别
类型 不同大肠杆菌菌斑平板上表型
野生型 rI r II
r III
B
K(λ)
S
小噬菌斑 小噬菌斑 小噬菌斑
大噬菌斑 大噬菌斑 大噬菌斑
大噬菌斑 无噬菌斑 小噬菌斑 (致死)
小噬菌斑 大噬菌斑 大噬菌斑
第二节 噬菌体突变型的重组测验
一.Benzer的重组测验与基因的精细结构分 析
拟等位基因(pseudoalleles):表型效应相似, 功能密切相关,在染色体上的位置又紧密连 锁的基因。这些基因产生的表型相似,但位 置上并不等位。
2. 互补测验原理与方法
互补测验(complementation test):也称顺 反测验(cis-trans test),指将两个突变分别 处于顺式和反式, 根据其表型确定两个突变 是否是同一基因的试验(看顺式反式排列是 否有功能上的互补)。