基因工程:第四章2
基因工程载体

二、噬菌体载体 (一)、λ噬菌体载体
1、λ噬菌体载体的特征
λ噬菌体颗粒中DNA为线状双链DNA分子,全长48502bp, 两端各有一段长度为12个核苷酸的互补单链(粘端),称 为cos位点。λ 噬菌体有61个基因,其中有1/3的区域是其 裂解性生长的非必需区,这一区段的缺失,或在此区段中 插入外源DNA,并不影响噬菌体的增殖,这就是λ 噬菌体 可作为基因载体的依据 。
第一节
微生物基因工程载体
克隆载体(cloning vecto载体(expression vector):能使目的基因在宿 主细胞中表达的一类载体。这类载体既有复制子, 更要有强启动子; 穿梭载体(shuttle vector):这类载体可以在原核细 胞中复制,也可在真核细胞中扩增和表达。
四、酵母菌载体
多数酵母中含有一种能独立复制的环状双链DNA,称为 2μ 质粒,长约6.3kb,有单一的复制起始点和一个自主 复制功能区域(ARS片段)。 根据质粒的复制方式不同将它们分为:整合型、复制型、 附加型和稳定型等类型。
共同的特点:
①含有E.coli质粒的复制起始序列,这样在外源基因转到酵 母细胞前可先在大肠杆菌中扩增。
外源基因的插入:
PstI Amp r Tet r Amp s Tet r
重组体的筛选
. .. . . . . .
涂布有Tet的培养基
.. .. . .
涂布有Amp的培养基
3、pUC系列质粒载体
(1)特点 具有更小的分子 量(2686bp)和 更高的拷贝数。 具有多克隆位点
可用组化方法鉴别:
含有一个来自于大肠杆菌的经过加工的LacZ基因 (LacZ′),它编码β -半乳糖苷酶氨基端146个氨基酸 , 可以和β -半乳糖苷酶缺陷型的大肠杆菌实现基因内互补, 即α —互补 ,恢复分解乳糖的能力。 X-gal也是β -半乳糖苷酶的一种底物,经降解后可生成溴 氯吲哚,使大肠杆菌菌落呈蓝色。当无β -半乳糖苷酶时, X-gal不被分解,菌落呈白色。 当外源基因插入到pUC质 粒的LacZ′基因内部,则LacZ′基因受到破坏,便不能 再和缺陷型受体菌中生成有活性的β -半乳糖苷酶,因此, 菌落呈白色。 反之,非重组体为蓝色菌落。 可通过插入失活法进行筛选
基因工程第四章载体
(4) 插入失活型质粒载体
载体的克隆位点位于其某一个选择性 标记基因内部。
如pDF41、pDF42、pBR329。
外源DNA
抗菌素抗性
无抗菌素抗性
(5)正选择的质粒载体 Direct selection vectors
直接选择转化后的细胞。
只有带有选择标记基因的转化菌细胞才 能在选择培养基上生长。
如pUR2、pTR262等。
目前通用的绝大部分质粒载体都是正 选择载体。
(6) 表达型质粒载体
主要用来使外源基因表达出蛋白质产物。
注意启动子的性质,终止子、起始 密码、终止密码的阅读正确。
如果在大肠杆菌里表达,必须把所克隆的 真核生物的基因置于大肠杆菌的转录—翻 译信号控制之下。
表达载体的结构
1)普通载体元件
b)细菌抗性原理 Ampr基因编码-内酰胺酶,特异地 切割氨苄青霉素的-内酰胺环。
ii)氯霉素(chloramphenicol,Cml)
a)抑菌原理 通过与50S核糖体亚基结合,干扰细胞 蛋白质的合成并阻止肽键的形成。杀死 生长的细菌。
b)细菌抗性原理
Cmlr 编码乙酰转移酶,特异地使氯霉 素乙酰化而失活。
(2)长度 6.3 kb。
(3)选择标记
大肠杆菌素(colicin)E1和对E1免疫 的基因(immE1)
① colicin E1基因的结构
cea 结构基因
imm
kil
免疫基因 溶菌基因
② 杀死不含有ColE1细菌的原因 cea + kil基因产物
③ 不被其他细菌的colicin E1所杀死的原因 imm基因
① 双抗菌素抗性选择标记 插入失活,分两次先后选择: 没有获得载体的寄主细胞 在Amp或Tet中都死亡。
浙江版生物必修二第四章 第二节 基因重组使子代出现变异

(5)将AAbb和aaBB杂交得到AaBb的过程实现了基因重组。杂种后代出现 性状差异的主要原因是基因突变。( × )
2.下列过程中未发生基因重组的是( ) A.表型正常的双亲生下患苯丙酮尿症的孩子 B.将人胰岛素基因导入大肠杆菌,利用大肠杆菌生产人胰岛素 C.黑色家鼠和白化家鼠杂交,F1全为黑鼠,F2中黑鼠∶淡黄鼠∶白鼠 =9∶3∶4 D.灰身长翅果蝇和黑身残翅果蝇杂交,F1全为灰身长翅,F1的测交后代出现 四种表型,比例为42∶42∶8∶8
2022 高 中 同 步 学 案 优 化 设 计 GAO ZHONG TONG BU XUE AN YOU HUA SHE JI 第四章 第二节 基因重组使子代出现变异
内
01 课前篇 自主预习
容
索
02 课堂篇 探究学习
引
课标阐释
学法指导
1.举例概述基因重组及其意义。
(生命观念)
1.回顾减数分裂中染色体的行为
2.非同源染色体间的自由组合导致基因重组 (1)生物在通过减数分裂形成配子时,非同源染色体的自由组合,导致非同 源染色体上的非等位基因也自由组合,因此产生多种类型的配子。雌、雄 配子结合后形成的新个体的基因型就有可能与亲代的基因型和表型不同。 (2)在完全显性的条件下,当控制两对相对性状的基因位于两对染色体上 时,F2的表型种类为22=4;如果涉及的各种性状分别由n对基因控制,并且各 对基因分别位于不同的染色体上,F2的表型数目是2n。
预习反馈 1.判断正误。 (1)杂交育种可将多个优良性状组合在同一个新品种中。一般可以通过杂 交、选择、纯合化等手段培养出新品种。( √ ) (2)将抗虫基因导入玉米的体细胞中,培育成抗虫玉米新品种的育种方法属 于杂交育种。( × ) (3)转基因技术人为地增加了生物变异的范围,实现种间遗传物质的交换。 (√ ) (4)杂交育种的原理是同源染色体上非等位基因自由组合。两个不抗病的 亲本杂交后代出现抗病类型是因为发生了基因重组。( × )
第四章-2基因工程

用纤维素纯化 polyRNA 没被降解 方法: mRNA的翻译 : 用体外蛋白质合成检测 mRNAs 。(麦胚无细胞转译体系或兔网织细 胞裂解物转译体系) 通过凝胶电泳分析mRNAs : 用琼脂糖或聚丙 烯酰胺凝胶电泳
DNA克隆都含有一种mRNA序列,
在选择中出现假阳性的几率比较低; 4. 克隆在细菌中表达的基因,用作基因序列的测定,
和发育过程中或组织中基因特异表达
cDNA法克隆目的基因的局限性
并非所有的mRNA分子都具有polyA结构 细菌或原核生物的mRNA半衰期很短 mRNA在细胞中含量少,对酶和碱极为敏感,
一般来说,目的基因的克隆战略分为两大类:另一类是利用PCR扩增技术甚至化学合成法体外直接合成目 的基因,然后将之克隆表达。
A 鸟枪法
鸟枪法克隆目的基因的基本战略 鸟枪法操作的改进 鸟枪法克隆目的基因的局限性
引导合成法:获得的双链cDNA 能保留完整的5’端序列
G 5‘ppp’5G 3‘ HO
G 5‘ppp’5G 3‘ HOCCCCCCC
TdT NaOH
3‘ HOCCCCCCC
退火
3‘ HOCCCCCCC 5‘ pGGGGGGG
Klenow
3‘ HOCCCCCCC 5‘ pGGGGGGG
dCTP dNTPs
目的基因
5
5
‘
变性
加热
‘
5 ‘
退火
5 ‘
5 聚合
‘ 5 ‘
5
‘ 变性
5 ‘
5 ‘
引物
5 ‘
底物
5 ‘
加热
5 ‘
5
‘
3
1
5 ‘
5
‘
2
退火 引物
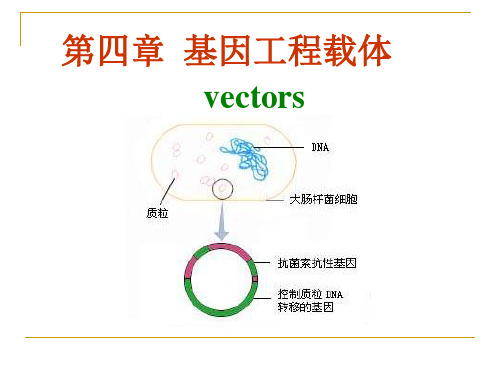
第四章 基因工程的质粒载体

SC
2 质粒DNA的转移
(1)质粒的类型:在大肠杆菌中的质粒,可 以分为:
接合型质粒:能自我转移
具有自主复制的基因,控制细菌配对和质粒接合转 移的基因。
非接合型质粒 不能自我转移
按接合转移功 能分类
非接合型质粒
主要基因
自主复制基因,产生大肠杆菌素基因
按抗性记号 分类
Col质粒
接合型质粒
自主复制基因,抗菌素抗性基 因
第二代 酵母表达 穿梭质粒 体系
第三代 哺乳类细 病毒、脂质体 胞表达体系
第四代 基因直接 DNA本身 导入
细菌 酵母 培养动物细胞 生殖细胞、 体细胞、个体
(三)基因工程载体必须具备的条件:
※(1)有复制起点 ※(2)具有若干个限制性内切酶的单一识别位点 ※(3)具备合适的筛选标记 ※(4)具备合适的拷贝数目
(c)所示,F质粒无力帮助mob-突变体进行转移,其中F性须和转移装置虽已 形成,但ColE1 DNA并没有发生缺口。
(d)表示另一种具mob+表型并带有一个顺式显性突变的ColE1突变体,它缺 失了bom位点。在这样的寄主细胞中,虽然能够合成mob蛋白质,但由于不 能发生缺口,因此仍然不能够转移。
3.若质粒DNA经过适当的核酸内切 限制酶切割之后,发生双链断裂形成 线性分子(IDNA),通称L构型
基因工程:第四章-酵母基因工程

UBC4-UBC5双突变型:
UBC4-UBC5双突变型能大幅度削弱泛
素介导的蛋白降解。
7个泛素连接酶基因的突变对衰减蛋白 降解作用同样有效。
6、内源性蛋白酶缺陷型的突变宿主菌
酿酒酵母具有20多种蛋白酶 空泡蛋白酶基因PEP4野生型和
pep4-3突变株
B-半乳糖苷酶活性明显升高
(三) 酵母菌的载体系统
酵母基因工程
酵母菌作为外源基因表达受体菌的特征 酵母菌的宿主系统 酵母菌的载体系统 酵母菌的转化系统 酵母菌的表达系统 利用重组酵母生产乙肝疫苗
1974 Clarck-Walker和Miklos发现在多数酿酒酵母 中存在质粒。
1978 Hinnen将来自一株酿酒酵母的leu2基因导入 另一株酿酒酵母,弥补了后者leu2的缺陷, 标志着酵母表达系统建立。
酵母菌有4个泛素编码基因:
UBI1 编码泛素-羧基延伸蛋白52 对数生长期表达 稳定期关闭
UBI2 编码泛素-羧基延伸蛋白52 对数生长期表达 稳定期关闭
UBI3 编码泛素-羧基延伸蛋白76 对数生长期表达 稳定期关闭
UBI4 编码泛素五聚体
对数生长期关闭 稳定期表达
酵母菌有7个泛素连接酶基因:
UBC1、UBC2、UBC3、UBC4、UBC5、UBC6、UBC7
酵母菌表达外源基因的优势: 全基因组测序,基因表达调控机理清楚,遗传 操作简便。 具有真核生物蛋白翻译后加工修饰系统。 能将外源基因表达产物分泌至培养基中。 大规模发酵工艺简单、成本低廉。
不含特异性病毒、不产毒素,被美国FDA认定为 安全的基因工程受体系统。
酵母菌表达外源基因的缺点:
表达产物的糖基化位点和结构特点 与高等真核生物有差距。
特点:
基因工程 电子版
作者:吴乃虎出版社:高等教育出版社第一章基因工程概述第一节基因操作与基因工程一、基因操作与基因工程的关系二、基因工程的诞生与发展第二节基因工程是生物科学发展的必然产物一、基因是基因重组的物质基础二、DNA的结构和功能三、基因操作技术的发展促进基因工程的诞生和发展四、基因工程的内容第三节基因的结构——基因操作的理论基础一、基因的结构组成对基因操作的影响二、基因克隆的通用策略第一篇基因操作原理第二章分子克隆工具酶第一节限制性内切酶一、限制与修饰二、限制酶识别的序列三、限制酶产生的末端四、DNA末端长度对限制酶切割的影响五、位点偏爱六、酶切反应条件七、星星活性八、单链DNA的切割九、酶切位点的引入十、影响酶活性的因素十一、酶切位点在基因组中分布的不均一性第二节甲基化酶一、甲基化酶的种类二、依赖于甲基化的限制系统三、甲基化对限制酶切的影响第三节DNA聚合酶一、大肠杆菌DNA聚合酶二、KIenow DNA聚合酶三、T4噬菌体DNA聚合酶四、T7噬菌体DNA聚合酶五、耐热DNA聚合酶六、反转录酶七、末端转移酶第四节其他分子克隆工具酶一、依赖于DNA的RNA聚合酶二、连接酶三、T4多核苷酸激酶四、碱性磷酸酶五、核酸酶六、核酸酶抑制剂七、琼脂糖酶八、DNA结合蛋白九、其他酶第三章分子克隆载体第一节质粒载体一、质粒的基本特性二、标记基因三、质粒载体的种类第二节λ噬菌体载体一、λ噬菌体的分子生物学二、λ噬菌体载体的选择标记……第四章人工染色体载体第五章表达载体第六章基因操作中大分子的分离和分析第七章基因芯片技术第八章PCR技术及其应用第九章DNA序列分析第十章DNA诱变第十一章DNA文库的构建和目的基因的筛选第十二章基因组研究技术第二篇基因工程应用第十三章植物基因工程第十四章动物基因工程第十五章酵母基因工程第十六章细菌基因工程第十七章病毒基因工程第十八章医药基因工程第十九章基因工程产品的安全及其管理第一章基因工程概述第一节基因操作与基因工程一、基因操作与基因工程的关系基因操作(gene manipulation):指对基因进行分离、分析、改造、检测、表达、重组和转移等操作的总称。
基因工程-第四章
(4) 插入失活型质粒载体 载体的克隆位点位于其某一个选择性标记基因内部。
抗菌素抗性
外源DNA 无抗菌素抗性
(5)正选择的质粒载体(Direct selection vectors)
直接选择转化后的细胞。只有带有选择标记基因的转化 菌细胞才能在选择培养基上生长。
目前通用的绝大部分质粒载体都是正选择载体。
各类载体
pBR322 外源基因Pst I
Tet中存活 但在Amp中死亡
外源基因BamH I Amp中存活 但在Tet中死亡
pBR322 外源基因Pst I
Tet中存活 但在Amp中死亡
外源基因BamH I Amp中存活 但在Tet中死亡
3. pUC系列
•University of California 的 J. Messing 和 J. Vieria 于1978年,在pBR322 的基础上改造而成,如 pUC7、pUC8、pUC9、pUC10、pUC11、pUC18、 pUC19。 • 元件来源
• 质粒空间构型与电泳速率
分子量相同的,scDNA最快、l DNA次之、ocDNA最 慢。
OC L
SC
质粒的生物学基本特性
1.自主复制性
• 质粒复制子是质粒 DNA 中能自主复制并维持正常拷贝数的 一段最小的核酸序列单位。
• 两部分组成:复制起始区(ori)及其相关的调控元件。 • 质粒能利用寄主细胞的 DNA 复制系统进行自主复制。 • 质粒 DNA 上的复制子结构决定了质粒与寄主的对应关系
必要的条件能力。
理想载体至少必备的条件
① 能在宿主细胞中自主复制 ②容易进入宿主细胞 ③ 容易插入外来核酸片段 ④ 容易从宿主细胞中分离纯化,便于重组操作 ⑤ 具有合适的筛选标记 ⑥ 具有针对受体细胞的亲缘性或亲和性
分子生物学第四章--基因工程常用工具酶
同裂酶:识别位点相同,酶的来源不同。
同尾酶:识别位点不同,切出片段有相同末端序列。
B.以切出片段末端性质不同可分,粘性末端和平末端。
粘性末端:(Cohesive Ends)两个突出末端可退火互补— — DNA是分子重组的基础
15
同裂酶
又称异源同工酶。指来源不同,但具有相同的识别 序列。 在切割DNA时,其切割点可以是相同的,产生平 头末端,称为同识同切; 切割点也可以是不同的,产生3ˊ或5ˊ粘性末端, 称为同识异切。
第四章 基因工程常用工具酶
1
Manipulating Genes
- Transferring Genes
Restriction Ligation Extract DNA
Transformation
Selection
Culturing
2
重组DNA实验中常见的主要工具酶
3
我们的基本目的是:把外源基因与载体 连接在一起形成重组DNA分子,最少需要以 下两类工具酶:
23
如果用一种限制酶,切割两种不同的DNA时,
产生相同的末端,混合后“退火”,这两种不同的
DNA分子彼此可以连接,形成重组DNA分子。
24
限制性内切酶的剪切方式
25
Yu Zheng, et al. Using shotgun sequence data to find active restriction enzyme genes. Nucleic Acids Res., 2009, 37: e1. Whole genome shotgun sequence analysis has become the standard method for beginning to determine a genome sequence. The preparation of the shotgun sequence clones is, in fact, a biological experiment. It determines which segments of the genome can be cloned into Escherichia coli and which cannot. By analyzing the complete set of sequences from such an experiment, it is possible to identify genes lethal to E. coli.
基因工程复习资料
基因工程复习资料第一章核酸的制备1.主要步骤:分、切、接、转、筛、表2.基因工程的概念:基因工程又称基因堆叠技术和dna重组技术,就是以分子遗传学为理论基为础,以分子生物学和微生物学的现代方法为手段,将不同来源的基因按预先设计的蓝图,在体外构建杂种dna分子,然后导入活细胞,以改变生物原有的遗传特性、获得新品种、生产新产品。
第二章基因工程工具酶1.生物催化剂:核酶、抗体酶、模拟酶。
2.限制性内切核酸酶:定义:限制性内乌核酸酶就是一类能够辨识双链dna中特定核苷酸序列(辨识序列),并在识别序列上使每条链的一个磷酸二酯键断开的内脱氧核糖核酸酶。
命名:限制性内乌核酸酶通常就是以第一次抽取至这类酶的生物的种名的第一个字母和种名的第一、第二个字母命名的,有的在后面还加菌株(型)代号中的一个字母。
如果从同一种生物中先后提取到多种限制性内切核酸酶,则依次用罗马数字ⅰ、ⅱ、ⅲ表示。
并且名称的前三个字母须用斜体,第一个字母用大写。
3.dna连接酶:定义:dna连接酶也称dna黏合酶,在分子生物学中扮演一个既特殊又关键的角色,那就是连接dna链3‘-oh末端和,另一dna链的5’-p末端,使二者生成磷酸二酯键,从而把两段相邻的dna链连成完整的链的一种酶。
种类:大肠杆菌dna连接酶、t4dna连接酶、tscdna连接酶、真核生物细胞辨认出的连接酶,例如酶ⅰ、酶ⅱ、酶ⅲ等多种类型。
4.dna片段的相连接方法:①具互补黏性末端dna片段之间的连接:可用e?colidna连接酶,也可用t4dna连接酶。
②尼奥罗末端dna片段之间的相连接:就可以用t4dna连接酶,并且必须减少酶的用量。
③dna片段末端修饰后进行连接:dna片段末端同聚物加尾后进行连接,可按互补粘性末端片段之间的连接方法进行连接;粘性末端修饰成平末端后进行连接;dna片段5′端脱磷酸化后进行连接;dna片段加连杆或衔接头后连接。
5.dna聚合酶:①定义:dna聚合酶就是指用dna单链为模板,以4种脱氧核苷酸为底物,催化剂制备一条与模板链序列优势互补的dna新链的酶。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
4.2.5 常用λ噬菌体载体
1.Charon载体
* Charon 16A:是一种插入型载体,它的β-半乳糖苷酶基因(lacZ)上有一个EcoR I切点,插入外源DNA片段后,lacZ基因失活,在补加有IPTG和X-gal的培养平板上形成无色噬菌斑,很易检出。
该载体一般可容纳10 kb的外源DNA片段。
* Charon 4:是一种替换型载体,它的可被外源DNA取代的EcoR I片段中包含大肠杆菌的lac5区段,其上有lacZ基因,以及它的操纵基因和启动基因。
该EcoR I片段被取代后的载体感染lac-指示菌,只能产生无色的噬菌斑。
该载体可容纳23 kb的外源DNA片段。
* Charon 28:是一种替换型载体,它的可取代区段两端各有一个BamH I切点,内部有red和gam基因,并且自身不含chi位点。
当外源DNA取代该片段后,就成为red-gam-,这样的噬菌体突变体获得了Spi-表型,能够在P2噬菌体溶源性的大肠杆菌细胞中生长,形成噬菌斑。
该载体可用于克隆Sau 3A部分消化的DNA 片段,容量可达20 kb。
2.EMBL系列载体
是一类替换型载体,适用于克隆长度为8~23 kb(几乎接近理论值)的外源DNA片段,常用作基因组文库的构建。
在λEMBL3和λEMBL4的可取代区段的两侧,各有一段切点相同、方向相反的多克隆位点,可被EcoR I、BamH I和Sal I所识别,因而可以用来克隆由这三类限制酶所产生的任何一种限制片段。
其中BamH I切点还可以连接经Sau 3A或Mbo I部分消化的DNA片段(因为它们产生的粘性末端完全一样,都是GA TC),虽然在克隆过程中载体上的BamH I位点被破坏了,但可以选用EcoR I或Sal I作消化作用,便能回收到克隆的外源DNA片段。
3.λgt系列载体
是一类插入型载体,常用于构建cDNA文库。
* λgt10:长度为43 kb,用于克隆小于6 kb的EcoR I切割片段。
EcoR I的单一切点位于cI基因内,该位点插入外源DNA片段后使cI基因失活,产生的重组噬菌体形成清晰的噬菌斑,有利于重组体的筛选。
* λgt11:长度为43.7 kb,用于克隆小于7.2 kb的EcoR I切割片段。
EcoR I的单一切点位于lacZ基因的近羧基端,该位点插入失活的重组噬菌体在IPTG和X-gal 平板上形成无色噬菌斑,很容易筛选。
λgt11的S基因含有琥珀突变(Sam100),在SupF(琥珀突变抑制基因)宿主菌内能正常增殖,在非抑制性宿主菌内,Sam100突变的噬菌体不能裂解细菌,从而使表达的融合蛋白在细胞内积累。
λgt11的cI857基因产生的λ阻遏蛋白是热不稳定的,在32℃时,有功能的λ阻遏蛋白使噬菌体倾向于溶源生长,宿主菌不发生裂解;42℃时,λ阻遏蛋白失活,噬菌体使宿主菌裂解。
* λgt22和λgt23:基本保留了λgt11的特点,不同的是λgt22和λgt23位于lacZ基因上的多克隆位点是由5个单一切点组成的,即Not I、Xba I、Sac I、Sal I和EcoR I。
由于真核DNA中Not I和Sal I位点很少,所以使用这两种酶进行克隆能显著增加获得全长cDNA的机率。
Not I:GC↓GGCCGC
Sal I:G↓TCGAC
λgt22和λgt23之间的差别是多克隆位点的方向正好相反。
4.3 柯斯质粒载体(cosmid vector)
是一类由人工构建的含有λDNA的cos序列和质粒复制子的特殊类型的质粒载体,也称粘粒载体。
4.3.1 柯斯质粒载体的组成与特点
1. 组成:
(1)一个质粒的复制起始部位和一个抗药性标记
(2)一个或多个限制酶的单一位点
(3)一个带有λ噬菌体cos位点的片段
(4)分子量小(4~6 kb),可容纳的真核DNA 达45 kb
柯斯质粒pHC 79兼备了λ噬菌体载体和pBR 322质粒载体两方面的优点:
来自pBR 322质粒部分的是一个完整的复制子,编码着一个复制起点和两个抗菌素抗性基因Amp R和Tet R。
来自λ DNA部分的片段,除了提供了cos位点外,在cos位点两侧还具有与噬菌体包装有关的DNA短序列,这样它能够包装成具有感染性的噬菌体颗粒。
2. 特点:
(1)具有λ噬菌体的特性:柯斯质粒载体连接适宜长度的外源DNA片段后,可在
体外包装成噬菌体颗粒,并能高效地转导对λ噬菌体敏感的大肠杆菌寄主细胞。
但由于其不含有λ噬菌体的全部基因,故不能通过溶菌周期,也不能形成子代噬菌体颗粒。
(2)具有质粒载体的特性:能象质粒DNA一样在寄主细胞内复制,也能在氯霉
素作用下进行扩增。
通常都具有抗菌素抗性基因,有些还带有基因插入失活的克隆位点,为重组体的筛选提供了方便的标记。
(3)具有高容量的克隆能力:λ噬菌体载体的克隆容量理论极限值是23 kb,一般
为15 kb左右,而cosmid载体的克隆极限可达45 kb左右。
这是因为cosmid 载体本身比较小,一般低于5 kb,这样插入一个45 kb的外源DNA片段,不
影响λ噬菌体地包装。
另一方面,由于包装限制的缘故,cosmid的克隆能力还有一个最低极限值,对于5 kb的cosmid载体,插入的外源DNA片段至少达到30 kb,才能包装成为具有感染性的λ噬菌体颗粒。
因此cosmid载体克隆大片段的DNA分子特别有效。
4.3.2 柯斯质粒载体中的克隆
λ噬菌体具有一种位点特异的切割体系,叫做末端酶或Ter体系,它能识别两个相距适宜的cos位点,把多连体分子切割成λ单位长度的片段,并把它们包装到λ噬菌体头部中去。
可以看出,Ter体系要求被包装的DNA片段具有两个cos位点,在这两个cos位点之间要保持36~52 kb的距离,这对于柯斯质粒克隆外源基因,进行体外包装是非常重要的。
1. 一般程序:
2. 在经两种限制酶消化的粘粒中进行克隆
3. 使用双cos位点载体进行粘粒克隆。