最新考研资料:北京大学细胞生物学第六章基质与内膜(中)
第六章细胞基质与内膜系统

4、解毒作用
• SER独特功能是对农药、污染物、毒素等有毒物 进行解毒。反应在肝细胞SER进行,故称肝细胞 的解毒作用。
• Cyt P450是肝细胞SER的膜蛋白,属单加氧酶, 或羟化酶。它催化O2中的1个氧原子加到不溶于 水的废物上使之羟化,溶于水并被转出细胞;另 一氧原子被NADH还原成水。
• 这些信号序列在蛋白质合成时将核糖体引导 到ER,进入ER后被切除。
• 1975年,Blobel正式提出信号假说
信号假说的要点:
• 1)蛋白的合成起始于胞质中游离核糖体
• 2)N端信号序列露出核糖体后,靠自由碰撞与ER 接触,N端信号序列的疏水性插入ER膜中;
• 3)蛋白质继续合成,以絆环形式穿过ER膜
3、脂的合成与转运
• SER是脂类合成主要场所。甘油三酯是由 SER合成并贮存ER腔中。
• 细胞膜所需的膜脂全都在SER合成, SER 上有合成磷脂所需的酶。
• SER合成的磷脂由胞质面转向ER腔面,转 位由ER膜中翻转酶帮助完成。
SER合成磷脂向其它膜结构转运 的2种方式:
1. 通过水溶性载体蛋白-磷脂交换蛋白(PEP), 在膜结构间转移磷脂:PEP与磷脂结合形成水 溶性复合物进入cytosol,扩散遇上其它膜后, PEP释放磷脂,将它插在膜上。
(1)起始转移信号
• 蛋白质N端的信号序列除作信号被SRP识别,还 具起始穿膜转移作用。
• 在蛋白质共翻译转移过程中,信号序列的N端 始终是朝ER外侧,插入转运通道后与通道内的 信号序列结合位点(受体)结合,其后的肽序列 是以伴环的形式通过运输通道。
• N端的起始转移序列是可切除的。
(2)内部信号序列
细胞生物学第六章总结

第六章线粒体与细胞能量转换一、基本特征1.詹纳斯绿Janus Green B一种活体染色剂,专一用于线粒体的染色。
它可以和线粒体中的细胞色素C氧化酶结合,从而出现蓝绿色。
2.结构1)外膜(outer membrane):线粒体最外层所包绕的一层单位膜,厚约5~7nm,光滑平整。
在组成上,外膜的脂质和蛋白质成分各占1/2。
2)内膜向基质折叠形成特定的内部空间内膜(inner membrane)比外膜稍薄,平均厚4.5nm,也是一层单位膜。
内膜的化学组成中20%是脂类,80%是蛋白质。
(基粒分为头部、柄部和基片三部分,是由多种蛋白质亚基组成的复合体。
基粒头部具有酶活性,能催化ADP磷酸化生成ATP,因此,基粒又称ATP合酶复合体)3)基质为物质氧化代谢提供场所线粒体中催化三羧酸循环、脂肪酸氧化、氨基酸分解、蛋白质合成等有关的酶都在基质中。
还含有线粒体独特的双链环状DNA、核糖体,这些构成了线粒体相对独立的遗传信息复制、转录和翻译系统。
4)内外膜转位接触点:核编码蛋白质进入线粒体的通道3.相对独立的遗传体系1)线粒体基因的转录i.线粒体mRNA不含内含子,也很少有非翻译区ii.每个mRNA5ˊ端的起始密码为AUG(或AUA),起始氨基酸为甲酰甲硫氨酸iii.线粒体的遗传密码也与核基因不完全相同iv.UAA的终止密码位于mRNA的3ˊ端。
某些情况下,一个碱基U就是mtDNA体系中的终止密码子v.线粒体与核密码子编码氨基酸三联体密码有差异2)线粒体DNA的复制mtDNA的复制起始点被分成两半,个是在重链上,称为重链复制起始点(O H),位于环的顶部,顺时针合成;一个是在轻链上,称为轻链复制起始点(O L),位于环L的“8点钟”位置,逆时针合成。
D型复制。
mtDNA复制不受细胞周期影响。
4.线粒体靶序列引导核编码蛋白质向线粒体转运1)核编码蛋白在进入线粒体需要分子伴侣蛋白的协助线粒体含有4个蛋白质输入的亚区域:线粒体外膜线粒体内膜膜间隙基质其中绝大多数线粒体蛋白被输入到基质,少数输入到膜间腔以及插入到内膜和外膜上。
细胞生物学:第6章细胞基质与细胞内膜系统
蛋白质在高尔基体中酶解加工类型
◆无生物活性的蛋白原(proprotein)高尔基体 切除N-端或两端的序列成熟的多肽。如胰岛素 、胰高血糖素及血清白蛋白等。
◆蛋白质前体高尔基体水解同种有活性的多肽 ,如神经肽等。
◆含有不同信号序列的蛋白质前体高尔基体加工 成不同的产物。
◆同一种蛋白质前体不同细胞、以不同的方式加工 不同的多肽。
30
电 子 显 微 镜 下 的 溶 酶 体
31
溶酶体膜稳定性
◆溶酶体的膜上嵌有质子泵 ◆溶酶体的膜上具有多种载体蛋白用 于水解产物向外转运 ◆溶酶体的膜蛋白高度糖基化 ◆溶酶体的膜含有能促进膜稳定性的 胆固醇
32
溶酶体的酶类
◆ 种类: ● A typical lysosome contains approximately 50
具消化作用
◆后溶酶体(post lysosome)
35
异 体 吞 噬
36
围歼线粒体
ER
自噬泡
37
溶酶体的功能
吞噬作用(Phagocytosis): 外来物质、衰老、死
亡的细胞(清理、防御、营养等)
自噬作用(Autophagy):细胞内受损、衰老的细
胞器、不需要的生物大分子(清理)
溶酶体病:各种储积症(如台萨氏病、糖原储积症)
沟通的网状结构。它从核膜延伸至细胞质中,靠近 细胞质内侧。
● ER的膜占细胞膜系统的一半 ●所包围的体积占细胞总体积的10%
内质网的化学组成
主要为蛋白质、脂类。 ◆内质网的标志酶是葡萄糖-6磷酸酶。
◆细胞色素P450在内质网膜中 最为丰富。
内 质 网 的 形 态 结 构
内质网种类
根据是否附有核糖体:
【生物课件】第六章 细胞基质与细胞内膜系统

③蛋白质的修饰加工
糖基化: 在内质网的腔面,寡糖链连接在插入膜内的磷酸多
萜醇上,当与糖基化有关的氨基酸残基出现后,通过 膜上的糖基转移酶,将寡糖基由磷酸多萜醇转移到相 应的天冬氨酸残基上。
N—连接的糖基化 糖:N—乙酰葡萄糖胺 氨基酸:天冬氨酸 发生部位:内质网(rER)
O—连接的糖基化 糖:N—乙酰半乳糖胺 氨基酸:丝氨酸、苏氨酸、羟赖氨酸、羟脯氨酸 发生部位:高尔基体(主要)
另外,在内质网上还发生羟基化和二硫键形成等
④新生多肽的折叠与装配
1、蛋白二硫键异构酶 切断二硫键,帮助其重新形成 二硫键,并处于正确的状态
2、结合蛋白(Bip) 能识别不正确的蛋白或未装配 好的蛋白亚单位,并促进其重 新折叠与组装
3、4肽信号 滞留在内质网中的蛋白质 的信号(-Lys-Asp-GluLeu-COO-) 如 蛋白二硫键异构酶 结合蛋白(Bip)
2.肺结核
结核杆菌不产生内、外毒素,也无荚膜和侵袭性酶。但是菌体成 分硫酸脑苷脂能抵抗胞内的溶菌杀伤作用,使结核杆菌在肺泡内大量 生长繁殖,导致巨噬细胞裂解,释放出的结核杆菌再被吞噬而重复上 述过程,最终引起肺组织钙化和纤维化。
3.各类贮积症
贮积症(storage disease):是由于遗传缺陷引起的,由于溶酶体 的酶发生变异,功能丧失,导致底物在溶酶体中大量贮积,进而影响细 胞功能,常见的贮积症主要有以下几类。
2、光滑内质网(sER)
常为分支管状,形成较为复杂的立体结构,表面没有核糖体附着
粗面内质网上有20种以上蛋白与光滑内质网不同
粗面内质网的形态
光滑内质网的形态
内质网
3、内质网与细胞内其它细胞器的关系
1、与细胞膜相连:甚至有管道相通 2、与外层核膜相接:内质网腔与核周隙相通 3、与高尔基体在结构、功能与发生上关系密切 4、rER与线粒体紧密相依:
考研资料:北京栗大学细胞生物学第六章基质与内膜(中)

more lysosomal enzymes, and resulting in accumulation of material in lysosomes as large inclusions.
② Lysosomes contain plenty acid hydrolases that can digest every kind of biological moleபைடு நூலகம்ule. ---the principal sites of intracellular digestion.
Marker enzyme: acid phosphatase
B. The Functions of Lysosomes
❖Lysosomes are involved in three major cell functions:
① phagocytosis; ② autophagy; ③ endocytosis.
Primary lys fuse with either phagocytic or autophagic vesicles, forming residual bodies that either undergo exocytosis or are retained in the cell as lipofuscin granules.
Transport proteins: transporting digested materials.
Figure 13-18 The low pH in lysosomes and endosomes. Proteins labeled with a pH-sensitive fluorescent probe (fluorescein) and then endocytosed by cells can be used to measure the pH in endosomes and lysosomes. The different colors reflect the pH that the fluorescent probe encounters in these organelles. The pH in lysosomes (red) is about 5, while the pH in various types of endosomes (blue and green) ranges from 5.5 to 6.5. (Courtesy of Fred Maxfield and Kenneth Dunn.)
细胞生物学6章 内膜

为什么游离核糖体不能与RER结合? 为什么游离核糖体不能与RER结合? RER结合 核糖体是如何附着在RER的胞质面上的? RER的胞质面上的 核糖体是如何附着在RER的胞质面上的? 何种机制引导核糖体与RER膜结合? RER膜结合 何种机制引导核糖体与RER膜结合? 蛋白质合成后如何进入RER腔中? RER腔中 蛋白质合成后如何进入RER腔中?
蛋白质折叠(folding) 2. 蛋白质折叠(folding)
驻留蛋白(retention protein) 驻留蛋白(retention 分子伴侣(molecular 分子伴侣(molecular chaperones) 能特异性识别新生肽链或部分折叠的多肽, 能特异性识别新生肽链或部分折叠的多肽, 与之结合,帮助多肽折叠、装配和转运, 与之结合,帮助多肽折叠、装配和转运,检查 折叠状态。其本身不参与终产物形成。 折叠状态。其本身不参与终产物形成。
2.GC与蛋白的糖基化修饰 GC与蛋白的糖基化修饰 GC
N-连接糖基化 N-连接糖基化
O-连接糖基化
糖基化修饰: 糖基化修饰: 1.由甘露糖苷酶Ⅰ 去掉大部分的甘露糖 甘露糖。 1.由甘露糖苷酶Ⅰ和Ⅱ去掉大部分的甘露糖。 2.乙酰氨基葡萄糖转移酶 乙酰氨基葡萄糖转移酶Ⅰ 2.乙酰氨基葡萄糖转移酶Ⅰ和Ⅱ将乙酰氨基葡萄糖 结合在存留的甘露糖基上。 结合在存留的甘露糖基上。 3.经半乳糖基转移酶和唾液酸转移酶作用 加上半 作用, 3.经半乳糖基转移酶和唾液酸转移酶作用,加上半 乳糖和唾液酸。 乳糖和唾液酸。
4.GC与蛋白质分选 GC与蛋白质分选 GC
分选信号 ( sorting signals)
救援受体 ( salvage receptor)
如:GC与溶酶体形成 GC与溶酶体形成
细胞生物学-第06章 细胞质基质与内膜系统

二、细胞质基质的功能:
第一节 细胞质溶质
(一) 中间代谢反应的进行
所有中间代谢都是在细胞质中进行的,其中大部分是 在细胞质溶质中进行的。
例如:糖酵解、核苷酸、脂肪酸和氨基酸代谢
反应必需的微环境;
细胞质基质 反应所需前体物;
合成细胞结构、完成生命活动的大分子前体物
糖 酵 解
细胞组分
数
目
体积比
细胞质基质
1
54
细胞核
1
6
内质网
1
12
高尔基体
1
3
溶酶体
3001胞内体2 Nhomakorabea01
过氧化物酶体
400
1
线粒体
1700
22
第一节 细胞质溶质
成分:中间代谢有关的酶类、细胞骨架结构。 特点:细胞质基质是一个高度有序的体系;通过弱键而相互
作用处于动态平衡的结构体系。
蛋白质
核糖体 RNA
50nm
在不同种类的细胞中, 分散高尔基体的大小、数 量和结构也会有所不同:
有些细胞:一个 真菌:一个潴泡 原生生物、无脊椎动物和高等植物细胞:多个 (肝细胞: 50个;有些藻类细胞: 25,000个)。
哺乳动物附睾管上皮细胞中的高尔基复合体
细胞中含有超大环形高尔基复合体
M ER
D
玉米根冠细胞中存在若干分散高尔基体
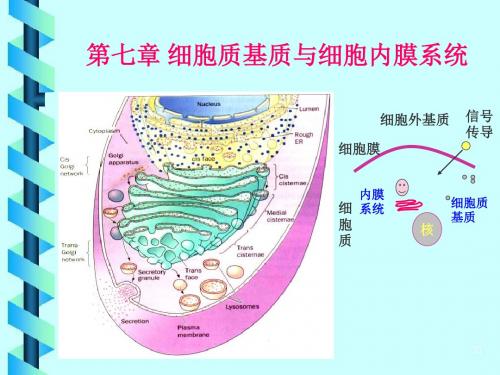
第七章 细胞质基质与内膜系统
Chapter 7 cytomatrix and endomembrane system
细胞质的组成
细胞质基质 较为均质和半透明的胶状物质
细胞质
如内质网、高尔基复合体、
有界膜的细胞器: 溶酶体、过氧化物酶体、
细胞生物学第6章内膜系统

溶酶体类型
(1)初级溶酶体(primary lysosome) (2)次级溶酶体(secondary lysosome)
是初级溶酶体和细胞内的吞噬泡、胞饮泡融合 形成的复合体,是进行消化作用的溶酶体,分为:
自噬溶酶体(autophagolysosome) 异噬溶酶体(phagolysosome)
内质网膜和细胞膜上的钙泵和钙通道对胞质中 游离Ca2+ 水平的调节非常重要。
内质网腔储存有大量的Ca2+ ,在接受上游信号 后,通过钙通道可迅速将Ca2+ 释放到胞质中,引 发细胞对信号的应答。
(6)其他功能:
❖ 具有解毒的功能,含有清除脂溶性的废物和代谢产生的 有害物质的酶。
❖ 含合成制造胆固醇并进一步产生固醇类激素的一系列酶。
(3)其它重要的生理功能
✓作为细胞内的消化“器官”为细胞提供营养 ;
➢ 分泌腺细胞中,溶酶体摄入分泌颗粒参与分 泌过程的调节;
参与清除赘生组织或退行性变化的细胞;
受精过程中的精子的顶体反应。
4.过氧化物酶体
又称微体(microbody) , 是由单层膜围绕 的内含一种或几种氧化酶类的异质性细胞 器。
2. 高尔基体
电镜下,由扁平膜囊和大小不等的囊泡构成, 不同细胞中扁囊的数目差异很大。
高尔基体是有极性的细胞器,表现于细胞中 的位置和方向、物质转运与生化极性。
冷冻蚀刻扫描电镜观察到 的高尔基体
电镜超薄切片观察到的高尔基体
高尔基体的组成
(1)高尔基体顺面网状结构(cis-Golgi network,CGN) 又称cis膜囊
蛋白质的氨基酸序列中,有决定其胞内定位和修 饰作用的有关信号,而且有决定寿命的信号。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
内膜(中)
4. The structure and functions of Lysosomes
A. Characteristics of Lysosomes
① Lysosome is a heterogenous organelle:
Primary lysosomes Second lysosomes
GlcNAc phosphotransferase
Recognition site binds to Signal patch
phosphodiesterase
Figure 6-40. The mannose 6-phosphate (M6P) pathway, the major route for targeting
脑硫脂沉积
GM1-半乳糖苷 酶 己糖胺酶A -半乳糖苷酶A 己糖胺酶A和B
葡糖脑苷酯酶
鞘磷脂水解酶 神经酰胺水解酶
半乳糖脑苷酯酶
芳基硫酸酯酶
神经节苷脂 GM1
神经节苷脂 GM2 三己糖神经酰 胺 神经节苷脂 GM2 和 红 细 胞 糖苷酯 葡糖脑苷脂
鞘磷脂
神经酰胺
半乳糖脑苷脂
脑硫脂
智力迟钝,肝脏肥大,骨 骼受累,2岁前死亡
智力迟钝,失明,3岁前死 亡
皮疹,肾功能丧失,下肢 疼痛
与泰-萨氏疾病症状相似, 但发展更快
肝脏和脾脏肿大,长骨腐 蚀,只在婴儿期发生智力 迟钝 肝脏和脾脏肿大,智力迟 钝
疼痛性与退行性的关节变 形,皮肤瘤,几年内死亡
髓磷脂缺失,智力迟钝,2 岁前死亡
智力迟钝,前十年死亡
D. Biogenesis of Lysosomes
Figure 6-23 The transport of newly synthesized lysosomal hydrolases to lysosomes. The
precursors of lysosomal hydrolases are covalently modified by the addition of mannose 6-phosphate in the CGN. They then become segregated from all other types of proteins in the TGN because a specific class of transport vesicles budding from the TGN concentrates mannose 6-phosphate-specific receptors, which bind the modified lysosomal hydrolases. These vesicles subsequently fuse with late endosomes. At the low pH of the late endosome the hydrolases dissociate from the receptors, which are recycled to the Golgi apparatus for further rounds of transport. In late endosomes the phosphate is removed from the mannose on the hydrolases, further ensuring that the hydrolases do not return to the Golgi apparatus with the receptor.
Mannose 6-phosphate residues target proteins to lysosomes
Targeting of soluble lysosomal enzymes to endosomes and lysosomes by M-6-P tag
Phosphorylation of mannose residues on lysosomal enzymes catalyzed by two enzymes
heterophagic autophagic Residual body
Primary Lys.
ห้องสมุดไป่ตู้
Second Lys
表1. 神经鞘脂贮积病
疾病
缺失酶类
主要贮积底物
后果
GM1 神 经 节 苷 脂 贮 积症 泰-萨二氏病 法布莱氏病 山霍夫氏病
高歇氏病
尼-皮二氏病 Farber’s 脂肪肉芽肿 病 Krabbe’s 病
lysosomal enzymes to lysosomes. Precursors of lysosomal enzymes migrate from the rER to the cis-Golgi where
mannose residues are phosphorylated. In the TGN, the phosphorylated enzymes bind to M6P receptors, which direct the enzymes into vesicles coated with the clathrin. The clathrin lattice surrounding these vesicles is rapidly depolymerized to its subunits, and the uncoated transport vesicles fuse with late endosomes. Within this low-pH compartment, the phosphorylated enzymes dissociate from the M6P receptors and then are dephosphorylated. The receptors recycle back to the Golgi, and the enzymes are incorporated into a different transport vesicle that buds from the late endosome and soon fuses with a lysosome. The sorting of lysosomal enzymes from secretory proteins thus occurs in the TGN, and these two classes of proteins are incorporated into different vesicles, which take different routes after they bud from the Golgi.[G. Griffiths et al., Cell 52:329; S. Kornfeld, Annu. Rev. Biochem. 61:307; and G. Griffiths and J. Gruenberg, Trends Cell Biol. 1:5]