微生物学课件第二节 细菌基因转移的方式
合集下载
微生物学课件第二节 细菌基因转移的方式

二十世纪生物科学具有划时代意义的巨大事件,推动了 生物科学的迅猛发展,并带动了生物技术产业的兴起。
微生物在基因工程的兴起和发展过程中起着 不可替代的作用!
“微生物与基因工程”
一、基因工程的基本过程
1. 基因分离: a)分别提取供体DNA和载体DNA b)用专一性很强的限制性核酸内切酶分别切割供体和载体DNA
Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子, 特称为F′因子。
F′×F-与F+×F-的不同:给体的
部分染色体基因随F′一起转入受体细胞
a)与染色体发生重组; b)继续存在于F′因子上,
形成一种部分二倍体;
二 细菌的转导(transduction)
由噬菌体介导的细菌细胞间进行遗传交换的一种方式: 一个细胞的DNA通过病毒载体的感染转移到另一个细胞中
能将一个细菌宿主的部分染色体或质粒DNA 带到另一个细菌的噬菌体称为转导噬菌体
细菌转导的二种类型:
普遍性转导 局限性转导
1 普遍性转导(generalized transduction)
噬菌体可以转导给体细菌染色体的任何部分到
受体细胞中的转导过程
1951年,Joshua Lederberg和Norton Zinder为了证实大肠杆菌以外 的其它菌种是否也存在接合作用,用二株具不同的多重营养缺陷型 的鼠伤寒沙门氏菌进行类似的实验:
抗生素筛选
G ene cloning and E xpression using Plasm id pB R 322
DNA聚合酶
能够把脱氧核糖核苷酸连续地加到双链DNA分子引物链的 3’-OH 末端,催化核苷酸的聚合作用,而不发生从引物模板上 解离的情况.
微生物在基因工程的兴起和发展过程中起着 不可替代的作用!
“微生物与基因工程”
一、基因工程的基本过程
1. 基因分离: a)分别提取供体DNA和载体DNA b)用专一性很强的限制性核酸内切酶分别切割供体和载体DNA
Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子, 特称为F′因子。
F′×F-与F+×F-的不同:给体的
部分染色体基因随F′一起转入受体细胞
a)与染色体发生重组; b)继续存在于F′因子上,
形成一种部分二倍体;
二 细菌的转导(transduction)
由噬菌体介导的细菌细胞间进行遗传交换的一种方式: 一个细胞的DNA通过病毒载体的感染转移到另一个细胞中
能将一个细菌宿主的部分染色体或质粒DNA 带到另一个细菌的噬菌体称为转导噬菌体
细菌转导的二种类型:
普遍性转导 局限性转导
1 普遍性转导(generalized transduction)
噬菌体可以转导给体细菌染色体的任何部分到
受体细胞中的转导过程
1951年,Joshua Lederberg和Norton Zinder为了证实大肠杆菌以外 的其它菌种是否也存在接合作用,用二株具不同的多重营养缺陷型 的鼠伤寒沙门氏菌进行类似的实验:
抗生素筛选
G ene cloning and E xpression using Plasm id pB R 322
DNA聚合酶
能够把脱氧核糖核苷酸连续地加到双链DNA分子引物链的 3’-OH 末端,催化核苷酸的聚合作用,而不发生从引物模板上 解离的情况.
基因的转移 PPT课件

25
二、转化的方法
转化大肠杆菌的方法有2类: 1. 感受态细胞法 将宿主细胞制备成感受态,这种细胞容 易接受外源DNA。 2. 电击法(电转化法) 利用电击的方法,在宿主细胞上瞬间打 “洞”,让外源DNA进入细胞内。
26
三 感受态细胞的制备
(一)基本概念 由于质粒载体缺失了mob 基因,不能 自行接合转移,必须改变E.coli 质膜的 通透性。 1. 感受态* Compenent 受体细胞经过诱导,产生一种短暂的吸 收外源DNA的生理状态。 2. 感受态细胞 Compenent cells 经物理和化学方法处理,受体细胞膜的 通透性改变,允许外源DNA 进入。
6.符合重组DNA操作的安全标准。
17
三、 E.coli受体细胞
1. 特性 用于转化的E.coli 细胞的特点
(1)限制修饰系统缺陷的突变体,即不含限制性 内切酶和甲基化酶的突变株 (Rˉ,Mˉ),
(2)允许外源DNA 进入E.coli体内,并稳定地遗 传给后代 。
18
2. 受体菌的条件与选择 符合生物安全性的要求。 宿主菌的限制酶和重组酶应为缺陷型。 宿主菌处于感受态。 转化率。 3. 常用E.coli菌株 DH5α、DH10B JM101~JM109 其它
8
体外包装好的重组噬菌体感染受体菌, 使受体菌发生溶菌,形成噬菌斑。每 g DNA能形成106噬菌斑。 当病毒从被感染的(供体)细胞释放出 来、再次感染另一(受体)细胞时,发 生在供体细胞与受体细胞之间的DNA 转移及重组。
9
图 转导作用
10
(3)体外包装的噬菌体的转导
① 体外包装 in vitro ing 定义:将重组的噬菌体DNA或Cosmid 质粒包装成具有感染能力的噬菌体颗 粒。
细菌的遗传与基因转移

● 05
第五章 垂直基因转移的意义
垂直基因转移的 过程
垂直基因转移是指细 菌通过有丝分裂或裂 殖进行基因传递。这 种过程能够帮助细菌 在繁殖和生长中传递 遗传信息,促进种群 的存活和演化。
垂直基因转移与遗传稳定性
保持遗传稳 定性
细菌种群
获得遗传信 息
后代细菌
垂直基因转移的演变
01、
重要作用
转导
噬菌体介导
细菌基因转移
遗传多样性
增加
基因交换
细菌之间
共轭
共轭是细菌之间通过 细胞接触传递质粒的 过程。这种方式使细 菌能够传递抗生素抗 性基因等重要遗传信 息,增加其生存能力 和抵抗力。共轭机制 在细菌族群中起着决 定性作用,是细菌遗 传及进化的重要途径。
水平基因转移的影响
适应基因
提高细菌的生存 能力
携带毒素基 因
对宿主细胞产生 毒性影响
质粒的复制
01、 独立于染色体的DNA复制过程
质粒可以独立完成复制过程
03、
依赖宿主细胞的复制机制
与宿主细胞的DNA复制过程相关联
02、 自主复制
依赖细菌自身的复制机制
04、
质粒的传递
质粒可以通过细菌之 间的接触传递给其他 细菌,导致基因交换。 这种传递方式使得细 菌的基因多样性增加, 更好地适应环境变化。
细菌的演化
传递有利基因
02、
促进进化
增长种群
进化演化
03、
影响细胞
增加多样性
改变遗传信息
04、
垂直基因转移的调控
01 多种调控机制
调节过程
02 适时进行
条件影响
03 提高效率
影响机制
2023年高中生物竞赛辅导课件:细菌基因转移和重组
L.Taturm E.coli 的多重营养缺陷型杂交实验
1958年获诺贝尔生理 学奖
一、细菌的接合作用
大肠杆菌的多重营养缺陷型杂交实验
两株多重营养缺陷型菌株单独不能生
长(左,右),只有在混合培养后才能
实
在平板上生长(中)
验
中间平板上长出的原养型菌落是两菌
证
株之间发生了遗传交换和重组所致!
据
(由营养缺陷恢复野生型表 型的菌株形成的菌落)
异常切割(低频):断裂和连接不是发 生在attP/attB 处,而是在原噬菌体临 近位点(包含gal 或bio基因)
正常的 切割
正常的 切割
λ噬菌体的溶源性反应:attP 位点与attB 位点 同源重组,噬菌体基因组整合到细菌染色体
异常切割(低频):断裂和连接不是发 生在attP/attB 处,而是在原噬菌体临 近位点(包含gal 或bio基因)
不同的微生物之间
微生物与高等动、植物之间
生物间基因的转移和交换是普遍存在的,是生物进化 的重要动力之一
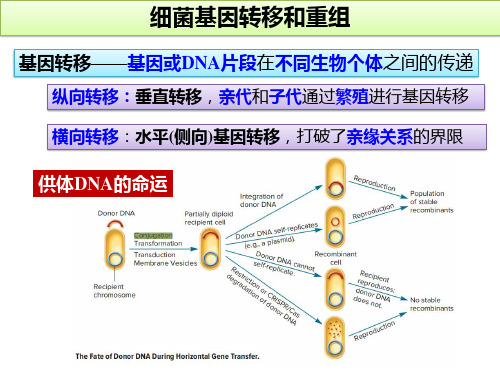
细菌基因转移和重组
细菌的三种水平基因转移形式
细菌的接合作用(Conjugation)
F+×F- 杂交、Hfr×F- 杂交、F’转导
细菌的转导(Transduction)
普遍性转导、局限性转导
细菌的遗传转化(Transformation)
二、细菌的转导(Transduction)
2、局限性转导
噬菌体总是携带同样的片段到受体细胞中
与普遍性转导的区别:
① 被转导的基因共价的与噬菌体DNA连接,与噬菌体 DNA一起进行复制、包装以及被导入受体细胞(普遍性 转导包装的全部是宿主菌的基因)
② 局限性转导颗粒携带特殊的染色体片段并将固定的个 别基因导入受体细胞(普遍性转导携带的宿主基因具有 随机性)
1958年获诺贝尔生理 学奖
一、细菌的接合作用
大肠杆菌的多重营养缺陷型杂交实验
两株多重营养缺陷型菌株单独不能生
长(左,右),只有在混合培养后才能
实
在平板上生长(中)
验
中间平板上长出的原养型菌落是两菌
证
株之间发生了遗传交换和重组所致!
据
(由营养缺陷恢复野生型表 型的菌株形成的菌落)
异常切割(低频):断裂和连接不是发 生在attP/attB 处,而是在原噬菌体临 近位点(包含gal 或bio基因)
正常的 切割
正常的 切割
λ噬菌体的溶源性反应:attP 位点与attB 位点 同源重组,噬菌体基因组整合到细菌染色体
异常切割(低频):断裂和连接不是发 生在attP/attB 处,而是在原噬菌体临 近位点(包含gal 或bio基因)
不同的微生物之间
微生物与高等动、植物之间
生物间基因的转移和交换是普遍存在的,是生物进化 的重要动力之一
细菌基因转移和重组
细菌的三种水平基因转移形式
细菌的接合作用(Conjugation)
F+×F- 杂交、Hfr×F- 杂交、F’转导
细菌的转导(Transduction)
普遍性转导、局限性转导
细菌的遗传转化(Transformation)
二、细菌的转导(Transduction)
2、局限性转导
噬菌体总是携带同样的片段到受体细胞中
与普遍性转导的区别:
① 被转导的基因共价的与噬菌体DNA连接,与噬菌体 DNA一起进行复制、包装以及被导入受体细胞(普遍性 转导包装的全部是宿主菌的基因)
② 局限性转导颗粒携带特殊的染色体片段并将固定的个 别基因导入受体细胞(普遍性转导携带的宿主基因具有 随机性)
第8章-微生物遗传-2_PPT幻灯片

• 转化DNA的最低浓度:在群体中含有15%感受态细胞时, 0.1gDNA/ml细胞悬液即可有效发生转化
1、自然遗传转化(简称自然转化)
1928年,Griffith发现肺炎链球菌(Streptococcus pneumoniae) 的转化现象
目前已知有二十多个种的细菌具有自然转化的能力
进行自然转化,需要二方面必要的条件:
– 感受态由细胞的遗传性决定,但同时也受环境因子的影 响:cAMP、Ca2+等最明显。如用CaCl2 处理E. coli可以 诱发其产生感受态。
• 感受态细胞的比例:当处于感受态高峰时,群体中呈感受态 的细胞因菌种而不同.
– Bacillus subtilis不超过10~15%
– Streptococcus pneuБайду номын сангаасonia和Haemophilus influenzae(流 感嗜血杆菌)达到100%
人工转化(artificial transformation)
转化(transformation)
1、转化及其发现:
R型活菌+S型死菌→ →S型活菌 ➢定义:受体菌自然或在人工技术作用下直接摄取来自供体菌 的游离DNA片段,并把它整合到自己的基因组中,而获得部 分新的遗传性状的基因转移过程,称为转化。转化后的的受 体菌称为转化子(transformant)。 ➢有关名词:
3、转化的类型:
根据感受态建立的方式,可以分为:
①自然遗传转化natural genetic transformation ②人工转化artificial transformation
感受态:
• 出现时间:只在细菌生长的某一时期出现;不同菌种的感受 态出现在不同生长时期
– Streptococcus pneumoniae的感受态出现在生长曲线中的 指数期的中期;Bacillus的一些种则往往出现在指数期末 及稳定期的初期。
1、自然遗传转化(简称自然转化)
1928年,Griffith发现肺炎链球菌(Streptococcus pneumoniae) 的转化现象
目前已知有二十多个种的细菌具有自然转化的能力
进行自然转化,需要二方面必要的条件:
– 感受态由细胞的遗传性决定,但同时也受环境因子的影 响:cAMP、Ca2+等最明显。如用CaCl2 处理E. coli可以 诱发其产生感受态。
• 感受态细胞的比例:当处于感受态高峰时,群体中呈感受态 的细胞因菌种而不同.
– Bacillus subtilis不超过10~15%
– Streptococcus pneuБайду номын сангаасonia和Haemophilus influenzae(流 感嗜血杆菌)达到100%
人工转化(artificial transformation)
转化(transformation)
1、转化及其发现:
R型活菌+S型死菌→ →S型活菌 ➢定义:受体菌自然或在人工技术作用下直接摄取来自供体菌 的游离DNA片段,并把它整合到自己的基因组中,而获得部 分新的遗传性状的基因转移过程,称为转化。转化后的的受 体菌称为转化子(transformant)。 ➢有关名词:
3、转化的类型:
根据感受态建立的方式,可以分为:
①自然遗传转化natural genetic transformation ②人工转化artificial transformation
感受态:
• 出现时间:只在细菌生长的某一时期出现;不同菌种的感受 态出现在不同生长时期
– Streptococcus pneumoniae的感受态出现在生长曲线中的 指数期的中期;Bacillus的一些种则往往出现在指数期末 及稳定期的初期。
微生物遗传学第四章 细菌转移(2)

2020/11/25
1. 接合现象的发现与证实
1946年,J.Lederberg & Tatum的大肠杆菌杂交试 验:
材料:大肠杆菌(E. coli) K12菌株的两个营养缺陷型品系:
菌株A—甲硫氨酸缺陷型met-和生物素缺陷型bio-; 菌株B—苏氨酸缺陷型thr-和亮氨酸缺陷型leu-。
2020/11/25
2.F质粒的发现
证明了细菌的接合是遗传物质的单向转移后, Hayes偶然发现了作为原始供体的A菌在冰箱里 存放了一年后出现一种变种,变种和正常的B菌 杂交时缺乏将遗传物质传给B菌株的能力。
他把这个不育变种的一个Strr 突变型分离出来, 并把它和可育的Strs A菌株一起繁殖,将其涂布 在含有链霉素的平板上,分离后再和B菌株杂交, 结果使不育的菌株回复了可育性(大约1/3恢 复)。
2020/11/25
有学者认为,具有性菌毛的细胞可以叫做 雄性,这种细丝状的菌毛像一种分子阴茎 ,与缺乏性菌毛的雌性细胞交合(德迪夫 1999)。
威廉斯( 2001)的观点:“在细菌和病毒 以及在所有高等生命体的主要类型中,遗 传重组现象的存在表明,性别的分子基础 是来自远古的进化演变的产物”。
14.11MCB 140 2/16/05 15
草履虫
MCB 140 2/16/05 16
W.Hayes的实验(1952)
(A)Strs (B)Strr
(A)Strr (B)Strs
A: met- bio- thr+ thi+ B: met+bio+ thr- thi-
(B)Strr
(A)Strr
⑥ 其特性类似于染色体,但染色体基因转移的频率不超过 10-6,F因子转移的频率高达70%以上。
1. 接合现象的发现与证实
1946年,J.Lederberg & Tatum的大肠杆菌杂交试 验:
材料:大肠杆菌(E. coli) K12菌株的两个营养缺陷型品系:
菌株A—甲硫氨酸缺陷型met-和生物素缺陷型bio-; 菌株B—苏氨酸缺陷型thr-和亮氨酸缺陷型leu-。
2020/11/25
2.F质粒的发现
证明了细菌的接合是遗传物质的单向转移后, Hayes偶然发现了作为原始供体的A菌在冰箱里 存放了一年后出现一种变种,变种和正常的B菌 杂交时缺乏将遗传物质传给B菌株的能力。
他把这个不育变种的一个Strr 突变型分离出来, 并把它和可育的Strs A菌株一起繁殖,将其涂布 在含有链霉素的平板上,分离后再和B菌株杂交, 结果使不育的菌株回复了可育性(大约1/3恢 复)。
2020/11/25
有学者认为,具有性菌毛的细胞可以叫做 雄性,这种细丝状的菌毛像一种分子阴茎 ,与缺乏性菌毛的雌性细胞交合(德迪夫 1999)。
威廉斯( 2001)的观点:“在细菌和病毒 以及在所有高等生命体的主要类型中,遗 传重组现象的存在表明,性别的分子基础 是来自远古的进化演变的产物”。
14.11MCB 140 2/16/05 15
草履虫
MCB 140 2/16/05 16
W.Hayes的实验(1952)
(A)Strs (B)Strr
(A)Strr (B)Strs
A: met- bio- thr+ thi+ B: met+bio+ thr- thi-
(B)Strr
(A)Strr
⑥ 其特性类似于染色体,但染色体基因转移的频率不超过 10-6,F因子转移的频率高达70%以上。
微生物学遗传与变异ppt课件

β- 棒状噬菌体
含毒素基因
编码毒素蛋白
• 毒力减弱—— 有毒菌株变异为弱毒或无毒菌株
卡介苗 Bacillus of Calmette- Guerin,BCG : 卡介二氏用有毒的牛 结核分枝杆菌在含甘油、马铃薯的培养基上,经13年连续230次传 代所获得的一毒力减弱但保留有抗原性的变异株。是 用于人工免疫 以预防结核病的活疫苗。
• 染色体DNA chromosome • 质粒 plasmid • 转位因子 transposable element • 噬菌体 phage
• 染色体DNA
chromosome
• 无内含子 • 重复序列少 • 功能相关基因组
成操纵子
• 病原菌中存在
毒力島(pathogenecity Island)
1.形态结构: EM 下 有 三 种 基 本 形 态 :
蝌蚪型 微球形 丝形
2.化学组成:
• 噬菌体由核酸和蛋白质组成。 • 核酸是噬菌体的遗传物质,根据其组成可为DNA噬
菌体和RNA噬菌体。
• 蛋白质是噬菌体头部衣壳及尾部的主要组成成份,
能保护噬菌体核酸,决定其外形和表面特征。
噬菌体与细菌的关系
4、耐药性变异variation of virulence
细菌对某种抗生素或药物由敏感变为不敏感即为细菌
的耐药现象。
多重耐药株:同时耐受多种抗生素的菌株。 抗生素依赖菌株:如痢疾志贺菌的赖链霉素菌株。
抗生素
抗生素
敏感
耐药
细菌遗传变异的物质基础
material basis of bacterial heredity and variation
转导频率
普遍性转导
局限性转导
裂解期
含毒素基因
编码毒素蛋白
• 毒力减弱—— 有毒菌株变异为弱毒或无毒菌株
卡介苗 Bacillus of Calmette- Guerin,BCG : 卡介二氏用有毒的牛 结核分枝杆菌在含甘油、马铃薯的培养基上,经13年连续230次传 代所获得的一毒力减弱但保留有抗原性的变异株。是 用于人工免疫 以预防结核病的活疫苗。
• 染色体DNA chromosome • 质粒 plasmid • 转位因子 transposable element • 噬菌体 phage
• 染色体DNA
chromosome
• 无内含子 • 重复序列少 • 功能相关基因组
成操纵子
• 病原菌中存在
毒力島(pathogenecity Island)
1.形态结构: EM 下 有 三 种 基 本 形 态 :
蝌蚪型 微球形 丝形
2.化学组成:
• 噬菌体由核酸和蛋白质组成。 • 核酸是噬菌体的遗传物质,根据其组成可为DNA噬
菌体和RNA噬菌体。
• 蛋白质是噬菌体头部衣壳及尾部的主要组成成份,
能保护噬菌体核酸,决定其外形和表面特征。
噬菌体与细菌的关系
4、耐药性变异variation of virulence
细菌对某种抗生素或药物由敏感变为不敏感即为细菌
的耐药现象。
多重耐药株:同时耐受多种抗生素的菌株。 抗生素依赖菌株:如痢疾志贺菌的赖链霉素菌株。
抗生素
抗生素
敏感
耐药
细菌遗传变异的物质基础
material basis of bacterial heredity and variation
转导频率
普遍性转导
局限性转导
裂解期
细菌基因转移和基因重组

整合就是指单链的转化DNA与受体DNA对应位点的置换,从而稳定
地掺入到受体DNA中的过程。
实际上就是一个遗传重组的过程。因而研究整合的分子机制事实上
也为遗传重组的分子机制作出了贡献。
二、转化因子的特性及感受态
(一)特性: 1、转化因子必需是双链DNA(见图) 2、单链DNA使细胞转化 3、转化因子的大小从0.5-15 kb
1
2
3
转化效率的高低决定于3个因素: 受体细胞的感受态,决定转化因子能否进 入细胞。 受体细胞的限制酶,决定转化因子在整合 前是否被降解。 供体与受体DNA的同源性,决定转化因子 能否被整合。
五、人工转化的方法
1. Ca2+ 等阳离子介导的转化 2. PEG介导的转化 3. 电穿孔法 4. 基因枪转化
该试验说明什么问题呢?
对正反杂交实验的解释:
A实验:菌株A被杀死,仍有子代,故认为菌株 A为供体(donors),相当于雄性亲本, 表示为F+ 。 B实验:菌株B被杀死,没有子代,故认为菌株 B为受体(recipients),相当于雌性亲 本,表示为F- 。
结论:遗传重组是一种 单向过程。 A:供体;B:受体
一、细菌接合的发现与证实
1、发现 Yalu University :Lederberg(1925 - ) Tatum(1909 - 1975) 实验材料: E.coli k12 58-161 met – bio – thr + leu + E.coli k12 w677 met + bio + thr – leu –
对外源DNA的摄取有特异性主要由特异
性摄取序列所决定。 例如:流感嗜血菌的特异摄取序列:
地掺入到受体DNA中的过程。
实际上就是一个遗传重组的过程。因而研究整合的分子机制事实上
也为遗传重组的分子机制作出了贡献。
二、转化因子的特性及感受态
(一)特性: 1、转化因子必需是双链DNA(见图) 2、单链DNA使细胞转化 3、转化因子的大小从0.5-15 kb
1
2
3
转化效率的高低决定于3个因素: 受体细胞的感受态,决定转化因子能否进 入细胞。 受体细胞的限制酶,决定转化因子在整合 前是否被降解。 供体与受体DNA的同源性,决定转化因子 能否被整合。
五、人工转化的方法
1. Ca2+ 等阳离子介导的转化 2. PEG介导的转化 3. 电穿孔法 4. 基因枪转化
该试验说明什么问题呢?
对正反杂交实验的解释:
A实验:菌株A被杀死,仍有子代,故认为菌株 A为供体(donors),相当于雄性亲本, 表示为F+ 。 B实验:菌株B被杀死,没有子代,故认为菌株 B为受体(recipients),相当于雌性亲 本,表示为F- 。
结论:遗传重组是一种 单向过程。 A:供体;B:受体
一、细菌接合的发现与证实
1、发现 Yalu University :Lederberg(1925 - ) Tatum(1909 - 1975) 实验材料: E.coli k12 58-161 met – bio – thr + leu + E.coli k12 w677 met + bio + thr – leu –
对外源DNA的摄取有特异性主要由特异
性摄取序列所决定。 例如:流感嗜血菌的特异摄取序列:
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Hfr菌株的F因子插入到染色体DNA上,因此只要发生接合转移转移 过程,就可以把部分甚至全部细菌染色体传递给F-细胞并发生重组 ,由此而得名为高频重组菌株。
Hfr菌株仍然保持着F+细胞的特征,具有F性菌毛,并象F+一样与 F-细胞进行接合。所不同的是,F因子的先导区(leading region)结 合着染色体DNA向受体细胞转移,F因子除先导区以外,其余绝 大部分是处于转移染色体的末端,由于转移过程常被中断,因此F 因子不易转入受体细胞中,故Hfr×F-杂交后的受体细胞(或接合 子)大多数仍然是F-。
第七章 微生物遗传和变异
一 细菌的接合作用(conjugation)
接合作用:通过细胞与细胞的直接接触而产生的遗 传信息转移和重组过程
证实接合过程需要细胞间的直接接触的 “U”型管实验( Bernard Davis,1950 )
2. 机制
接合作用是由一种被称为F因子的质粒介导 F因子的分子量通常为5×107 Da ,上面有编码细菌产生 性菌毛(sex pili)及控制接合过程进行的20多个基因。
d)F′菌株,Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子,特称为F′因 子。 细胞表面同样有性菌毛。
1) F+×F-杂交
F+菌株的F因子向F-细胞转移,但含F因子的宿主细胞 的染色体DNA一般不被转移。
杂交的结果:给体细胞和受体细胞均成为F+细胞
2)Hfr ×F-杂交
三 细菌的遗传转化(genetic transformation)
定义:游离DNA分子(质粒和染色体DNA)被自然或人工
感受态细胞摄取,并得到表达的基因转移过程。
1、转化
1928年,Griffith发现肺炎链球菌(Streptococcus pneumoniae) 的转化现象
目前已知有二十多个种的细菌具有自然转化的能力
Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子, 特称为F′因子。
F′×F-与F+×F-的不同:给体的
部分染色体基因随F′一起转入受体细胞
a)与染色体发生重组; b)继续存在于F′因子上,
形成一种部分二倍体;
二 细菌的转导(transduction)
由噬菌体介导的细菌细胞间进行遗传交换的一种方式: 一个细胞的DNA通过病毒载体的感染转移到另一个细胞中
含有F因子的细胞:“雄性”菌株(F+),其细胞表面有性菌毛 不含F因子的细胞:“雌性”菌株(F-),细胞表面没有性菌毛
F因子的四种细胞形式
a)F-菌株, 不含F因子,没有性菌毛,但可以通过 接合作用接 收F因子而变成雄性菌株(F+);
b)F+菌株, F因子独立存在,细胞表面有性菌毛。 c)Hfr菌株,F因子插入到染色体DNA上,细胞表面有性菌毛。
能将一个细菌宿主的部分染色体或质粒DNA 带到另一个细菌的噬菌体称为转导噬菌体
细菌转导的二种类型:
普遍性转导 局限性转导
1 普遍性转导(generalized transduction)
噬菌体可以转导给体细菌染色体的任何部分到
受体细胞中的转导过程
1951年,Joshua Lederberg和Norton Zinder为了证实大肠杆菌以外 的其它菌种是否也存在接合作用,用二株具不同的多重营养缺陷型 的鼠伤寒沙门氏菌进行类似的实验:
普 遍 性 转 导 过 程
2 局限性转导(specialized transduction) 把供体菌的少数特定基因转移到受体菌中的过程
温和噬菌体感染
整合到细菌染色体的特定位点上
宿主细胞发生溶源化
溶源菌因诱导而发生裂解时, 在前噬菌体二侧的少数宿主 基因因偶尔发生的不正常切 割而连在噬菌体DNA上
– 3、 DNA双链中的一条单链逐步降解,同时,另一条 单链逐步进入细胞。
– 4、 转化DNA单链与受体菌染色体组上的同源区段配 对,接着受体染色体组的相应单链片段被切除,并被外 来的单链DNA所交换和取代,于是形成了杂种DNA区 段。
– 5、受体菌染色体组进行复制,杂合区段分离成两个, 其中之一类似供体菌,另一类似受体菌。当细胞分裂后, 此染色体分离形成了一个转化子。
转化全过程
转化整合过程
转化过程的特点:
a)对核酸酶敏感; b)不需要活的DNA供体细胞; c)转化是否成功及转化效率的高低主要取决于 转化(DNA)给体菌株和转化受体菌株之间的 亲源关系; d)通常情况下质粒的自然转化效率低
进行转化,需要二方面必要的条件:
转化因子:外源游离dsDNA分子 受体细胞处于感受态:最容易接受外源DNA的一种生理状态
以革兰氏阳性的肺炎链球菌为材料,其转化过程大体是:
– 1、双链DNA片段与感受态受体菌的细胞表面特定位 点结合。
– 2、在位点上的DNA发生酶促分解,形成平均分子量 为(4~5) x 106D的DNA片段。
染色体上越靠近F因子的先导区的基因,进入的机会 就越多,在F-中出现重组子的的时间就越早,频率也高。
F因子不易转入受体细胞中,故Hfr×F杂交后的受体细胞(或称接合子)大多 数仍然是F-。
3细)胞F基′因×的F这-杂种交转移过程又常称为性导(sexduction)、F因子转导
(F-duction),或F因子媒介的转导(F-mediated transduction)。
部分缺陷的温和噬菌体
把供体菌的少数特定裂解时的 不正常切割:包含gal或 bio基因
(几率一般仅有10-6)
3 局限性转导与普遍性转导的主要区别:
a)普遍性转导是误包;局限性转导是误切。 b)局限性转导颗粒携带特定的染色体片段并将固定的个别基因
导入受体,故称为局限性转导。而普遍性转导携带的宿主基 因具有随机性,包装的可能全部是宿主菌的基因。
用“U”型管进行同样的实验时,在给体和受体细胞 不接触的情况下,同样出现原养型细菌!
沙门氏菌LT22A是携带P22噬菌体的溶源性细菌 另一株是非溶源性细菌
基因的传递很可能是由可透过“U”型管滤板 的
P22噬菌体介导的
一个表面看起来的常规研究却导致 一个惊奇和十分重要发现的重要例证!
(普遍性转导这一重要的基因转移途径的发现)
Hfr菌株仍然保持着F+细胞的特征,具有F性菌毛,并象F+一样与 F-细胞进行接合。所不同的是,F因子的先导区(leading region)结 合着染色体DNA向受体细胞转移,F因子除先导区以外,其余绝 大部分是处于转移染色体的末端,由于转移过程常被中断,因此F 因子不易转入受体细胞中,故Hfr×F-杂交后的受体细胞(或接合 子)大多数仍然是F-。
第七章 微生物遗传和变异
一 细菌的接合作用(conjugation)
接合作用:通过细胞与细胞的直接接触而产生的遗 传信息转移和重组过程
证实接合过程需要细胞间的直接接触的 “U”型管实验( Bernard Davis,1950 )
2. 机制
接合作用是由一种被称为F因子的质粒介导 F因子的分子量通常为5×107 Da ,上面有编码细菌产生 性菌毛(sex pili)及控制接合过程进行的20多个基因。
d)F′菌株,Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子,特称为F′因 子。 细胞表面同样有性菌毛。
1) F+×F-杂交
F+菌株的F因子向F-细胞转移,但含F因子的宿主细胞 的染色体DNA一般不被转移。
杂交的结果:给体细胞和受体细胞均成为F+细胞
2)Hfr ×F-杂交
三 细菌的遗传转化(genetic transformation)
定义:游离DNA分子(质粒和染色体DNA)被自然或人工
感受态细胞摄取,并得到表达的基因转移过程。
1、转化
1928年,Griffith发现肺炎链球菌(Streptococcus pneumoniae) 的转化现象
目前已知有二十多个种的细菌具有自然转化的能力
Hfr菌株内的F因子因不正常切割而脱离染色体时, 形成游离的但携带一小段染色体基因的F因子, 特称为F′因子。
F′×F-与F+×F-的不同:给体的
部分染色体基因随F′一起转入受体细胞
a)与染色体发生重组; b)继续存在于F′因子上,
形成一种部分二倍体;
二 细菌的转导(transduction)
由噬菌体介导的细菌细胞间进行遗传交换的一种方式: 一个细胞的DNA通过病毒载体的感染转移到另一个细胞中
含有F因子的细胞:“雄性”菌株(F+),其细胞表面有性菌毛 不含F因子的细胞:“雌性”菌株(F-),细胞表面没有性菌毛
F因子的四种细胞形式
a)F-菌株, 不含F因子,没有性菌毛,但可以通过 接合作用接 收F因子而变成雄性菌株(F+);
b)F+菌株, F因子独立存在,细胞表面有性菌毛。 c)Hfr菌株,F因子插入到染色体DNA上,细胞表面有性菌毛。
能将一个细菌宿主的部分染色体或质粒DNA 带到另一个细菌的噬菌体称为转导噬菌体
细菌转导的二种类型:
普遍性转导 局限性转导
1 普遍性转导(generalized transduction)
噬菌体可以转导给体细菌染色体的任何部分到
受体细胞中的转导过程
1951年,Joshua Lederberg和Norton Zinder为了证实大肠杆菌以外 的其它菌种是否也存在接合作用,用二株具不同的多重营养缺陷型 的鼠伤寒沙门氏菌进行类似的实验:
普 遍 性 转 导 过 程
2 局限性转导(specialized transduction) 把供体菌的少数特定基因转移到受体菌中的过程
温和噬菌体感染
整合到细菌染色体的特定位点上
宿主细胞发生溶源化
溶源菌因诱导而发生裂解时, 在前噬菌体二侧的少数宿主 基因因偶尔发生的不正常切 割而连在噬菌体DNA上
– 3、 DNA双链中的一条单链逐步降解,同时,另一条 单链逐步进入细胞。
– 4、 转化DNA单链与受体菌染色体组上的同源区段配 对,接着受体染色体组的相应单链片段被切除,并被外 来的单链DNA所交换和取代,于是形成了杂种DNA区 段。
– 5、受体菌染色体组进行复制,杂合区段分离成两个, 其中之一类似供体菌,另一类似受体菌。当细胞分裂后, 此染色体分离形成了一个转化子。
转化全过程
转化整合过程
转化过程的特点:
a)对核酸酶敏感; b)不需要活的DNA供体细胞; c)转化是否成功及转化效率的高低主要取决于 转化(DNA)给体菌株和转化受体菌株之间的 亲源关系; d)通常情况下质粒的自然转化效率低
进行转化,需要二方面必要的条件:
转化因子:外源游离dsDNA分子 受体细胞处于感受态:最容易接受外源DNA的一种生理状态
以革兰氏阳性的肺炎链球菌为材料,其转化过程大体是:
– 1、双链DNA片段与感受态受体菌的细胞表面特定位 点结合。
– 2、在位点上的DNA发生酶促分解,形成平均分子量 为(4~5) x 106D的DNA片段。
染色体上越靠近F因子的先导区的基因,进入的机会 就越多,在F-中出现重组子的的时间就越早,频率也高。
F因子不易转入受体细胞中,故Hfr×F杂交后的受体细胞(或称接合子)大多 数仍然是F-。
3细)胞F基′因×的F这-杂种交转移过程又常称为性导(sexduction)、F因子转导
(F-duction),或F因子媒介的转导(F-mediated transduction)。
部分缺陷的温和噬菌体
把供体菌的少数特定裂解时的 不正常切割:包含gal或 bio基因
(几率一般仅有10-6)
3 局限性转导与普遍性转导的主要区别:
a)普遍性转导是误包;局限性转导是误切。 b)局限性转导颗粒携带特定的染色体片段并将固定的个别基因
导入受体,故称为局限性转导。而普遍性转导携带的宿主基 因具有随机性,包装的可能全部是宿主菌的基因。
用“U”型管进行同样的实验时,在给体和受体细胞 不接触的情况下,同样出现原养型细菌!
沙门氏菌LT22A是携带P22噬菌体的溶源性细菌 另一株是非溶源性细菌
基因的传递很可能是由可透过“U”型管滤板 的
P22噬菌体介导的
一个表面看起来的常规研究却导致 一个惊奇和十分重要发现的重要例证!
(普遍性转导这一重要的基因转移途径的发现)