同源重组的分子机制_2
什么是同源重组的分子机制

什么是同源重组的分子机制关键信息项1、同源重组的定义2、参与同源重组的关键分子3、同源重组的启动条件4、同源重组过程中的关键步骤及相关机制5、同源重组的调控机制6、同源重组在生物体内的作用及意义1、同源重组的定义同源重组是指发生在两条同源DNA 序列之间的遗传物质交换过程。
这一过程对于维持基因组的稳定性、遗传多样性以及修复 DNA 损伤等方面具有重要意义。
11 同源重组的特点同源重组具有高度的准确性和特异性,它依赖于两条 DNA 链之间的同源性来实现遗传物质的交换。
12 同源重组与其他重组方式的区别与其他类型的重组(如位点特异性重组和转座重组)相比,同源重组涉及的区域更长,并且对同源序列的要求更高。
2、参与同源重组的关键分子21 重组酶重组酶是催化同源重组过程的关键酶类,如 RecA 家族的蛋白质。
211 RecA 蛋白的作用RecA 蛋白能够促进单链 DNA 与同源双链 DNA 进行链交换。
212 其他重组酶的功能除了 RecA 蛋白,还有一些其他的重组酶在同源重组中发挥着不同的作用。
22 单链结合蛋白单链结合蛋白能够稳定单链 DNA 结构,防止其重新形成双链。
23 核酸外切酶核酸外切酶在同源重组中参与切除DNA 链的末端,产生单链区域。
3、同源重组的启动条件31 DNA 损伤如 DNA 双链断裂是常见的启动同源重组的因素。
32 细胞周期阶段在特定的细胞周期阶段,如 S 期和 G2 期,同源重组的发生频率较高。
33 环境压力某些环境压力,如辐射、化学物质等,可能导致 DNA 损伤,从而启动同源重组。
4、同源重组过程中的关键步骤及相关机制41 产生 DNA 单链通过核酸内切酶或外切酶的作用,产生 DNA 单链。
411 单链的延伸和侵入单链 DNA 延伸,并侵入到同源双链 DNA 中,形成异源双链结构。
412 链交换和分支迁移在异源双链结构的基础上,发生链交换和分支迁移,促进遗传物质的交换。
42 形成 Holliday 连接体这是同源重组过程中的一个关键中间结构。
【遗传学】同源重组

【遗传学】同源重组同源重组是同源染色体间或同源序列之间某区段的重组,包括真核生物同源区段的交换、细菌同源区段的交换。
1.同源重组的基本条件1)具有同源区段同源重组常常发生在两个DNA分子的相同部位,但有时在具有同样或类似序列的不同部位间也发生重组,异位重组可引起DNA序列缺失、重复、倒位和其他DNA的重排现象。
但是不管同位重组还是异位重组,交换或重组序列必须相同或相似。
2)有联会发生3)DNA单链间能相互交换4)需蛋白因子参与2.同源重组的分子机制1)Holliday模型的重组过程(1)联会在晚偶线期和早粗线期染色体配对时,同源非姊妹染色单体的DNA分子联会在一起。
(2)酶切内切核酸酶识别DNA分子上的相应断裂点后,把磷酸二酯键切断,使两个同源非姊妹染色体DNA分子各有一条链断裂。
(3)交换重接两断链游离端的氢键断裂,离开互补单链。
落选局部放松,单链交换准备重接。
(4)形成交联桥结构在连接酶的作用下,断裂以交替方式与另一断裂点相互连接,行程一个交联桥,该结构称为Holloday中间体。
(5)分支迁移在重组接点处,单链可以从一条双螺旋进入另一条双螺旋,从而使重组接点像拉链一样滑来滑去,即重组接点可沿着两条双链移动,这种现象称为分支迁移。
某条单链上相当长的一段可以从一条DNA双螺旋上转移到另一条,形成长段的某种双链。
交联可靠拉链式活动,沿着配对DNA分子“传播”——桥迁,其中互补碱基间形成的氢键从一条链改变到另一条链,于是在两个亲本DNA分子间形成一段异源双链DNA,这种结构称Holliday结构。
(6)交联桥旋转交联的两臂环绕另外两臂旋转180°形成“十”字形。
(7)形成Holliday异构体在交联部分断开以消除交联体,回复两个线性DNA分子,形成Holliday结构的异构体。
(8)拆分沿左右或上下方向拆分Holliday中间体。
如果左右切断,即上连、下连,则产生的两个异源双链的两侧基因为AB和ab,仍保持亲代类型;如果上下切断,即左连、右连,则两侧基因为Ab和ab两个重组类型。
同源重组的分子机制

同源重组的分子机制一、断裂重接模型(breakage joining model)C.D.Darlington 1936年提出。
在同源染色体联会时,由于染色体的缠绕而产生张力,两个相对染色单体在同一位置断裂,然后彼此和另一染色单体重新连接起来从而形成重组并消除这种张力。
二、基因转换现象Olive等广泛研究粪生粪壳菌g座位,g-决定子囊孢子灰色,g+决定子囊孢子的黑色,在g+×g-的杂交中,他们分析了20万子囊,发现0.06%是5∶3分离,0.05%是6∶2分离,0.008%是3∶1∶1∶3(或异常4∶4)分离。
图示. (1)一个孢子中的两个孢子有着不同的基因型。
(2)分离比例不是4∶4。
(3)邻近的基因A/a都呈现正常分离。
断裂重接模型则无法解释异常现象。
1930年,德国遗传学家H.温克勒把这种不规则分离现象解释为减数分裂过程中同源染色体联会时一个基因使相对位置上基因发生相应的变化所致,因而称就基因转变。
好象是由于一个基因转换为另一等位基因,所以称为基因转换(gene conversion)。
以后由于发现一个基因发生基因转变时,它两旁的基因常同时发生重组:在5∶3和6∶2分离的子囊中,大约有30%也在g座位的这边或那边发生重组;有基因转换的子囊中,基因转换和遗传重组都发生在同样两个单体的子囊比例竟高达90%。
所以认为基因转变是某种形式的染色体交换的结果。
因此,基因转变的研究,实质上也是染色体交换机制的研究。
三、同源重组的Holliday模型1964年,R. Holliday提出了重组的杂合DNA模型(hybrid DNA mode),并作修正。
图示.过程:A.同源的非姊妹染色体的DNA配对。
B-C.同源非姊妹染色单体DNA中两个方向相同的单链在DNA内切酶的作用下,在相同位置上同时切开。
D.切开单链交换重接,形成交联桥结构(cross-bridged structure)。
E.交联桥的位置可以靠拉链式活动,沿着配对DNA分子“传播”—桥迁(Bridge migration),其中互补碱基间形成的氢键从一条链改变另一条链。
遗传学中的同源重组机制

遗传学中的同源重组机制遗传学是研究遗传变异和遗传规律的学科,其研究范围从微观的分子水平到宏观的群体水平。
在遗传学中,同源重组机制是基因重组的一种重要方式,它在基因转移、基因突变的形成以及进化等方面起着重要作用。
同源染色体在遗传学中,同源染色体是指在相同物种中两条染色体的基因组成相同或相似。
同源染色体可以是同一物种的不同个体所拥有的染色体,也可以是不同物种之间的染色体。
例如,人类和大猩猩的染色体虽然不完全相同,但是它们具有高度的同源性。
同源重组机制的基本原理同源重组机制是指两条同源染色体之间互相交换DNA片段的过程。
同源重组机制主要在减数分裂时发生,以确保每个生殖细胞中基因组的正确分离和再组合。
同源重组机制的过程主要分为四个步骤:一、同源染色体的配对在减数分裂开始时,同源染色体首先进行配对。
在此过程中,同源染色体会相互搜索并配对,然后相互交换DNA片段。
二、DNA切割和重组在同源染色体配对后,相对应的染色体会互相“交换”某一范围内的DNA片段。
这个过程是由酶催化完成的,其中最重要的酶是转移酶(recombination enzyme),它能够切割两根不同染色体或同一染色体的DNA链,以将碎片重组为新的DNA序列。
三、交换在DNA片段切割和重组后,相互对应的同源染色体会互相“交换”DNA片段,形成新的染色体组合。
这个过程称为同源重组。
四、交叉互换当相互配对的同源染色体上出现非互补的序列时,交叉互换便会发生。
在交叉互换的过程中,同源染色体上的非同源DNA片段将被交换到另一条染色体上,从而形成一个重组后的DNA序列。
同源重组在基因重组中的意义同源重组在基因重组中发挥着重要的作用。
基因重组是指基因片段的组合重组,形成新的基因序列,并在种群进化中起着关键作用。
同源重组机制不仅在有性生殖过程中发挥重要作用,也在无性生殖和DNA修复中发挥作用。
在进化过程中,同源重组机制也为物种的适应性进化提供了基本方式。
结论同源重组机制在遗传学中发挥着重要作用,是基因组进化和重组的基础。
同源重组

单链的侵入(strand invasion)由RecA蛋白介导。
RecA是一种单链DNA结合蛋白,参与大肠杆菌中所有 的同源重组事件。它催化一个双链DNA分子的3-末端 单链区侵入另一个双链DNA分子,形成异源双链区, 同时置换出同源单链,形成Holliday结构。
RecA介导的链交换可以分为三个阶段:RecA聚集在
SSB所结合的单链DNA上,而Mus81蛋白可能是拆分 Holliday中间体的酶。
二、位点特异性重组
位点特异性重组是发生在DNA上特定序列之间的重组,不依
赖于DNA顺序的同源性,由能识别特异DNA序列的蛋白质介导,
并不需要RecA蛋白和单链DNA。 λ噬菌体DNA侵入大肠杆菌细胞后,面临着裂解生长和溶原
在断裂处,MRX酶复合体利用其5’→3’的外切酶活性
降解DNA,生成3’单链末端,其长度通常可达1 Kb或更 长。MRX酶复合体由Mre11,Rad50和Xrs2三个亚基组成, 并以三个亚基的首字母命名。MRX酶复合体还被认为能 除去与DNA相连的Spo11。
在真核细胞中已经发现了两种与细菌RecA蛋白同源
分支迁移由结合在Holliday分支点上的RuvA和RuvB蛋 白催化。RuvA以四聚体的形式识别并结合到Holliday 分支点上。RuvB蛋白为催化分支迁移的解旋酶,但
RuvB自身不能有效地与DNA结合,它需要与RuvA一起
起作用。RuvC蛋白催化断裂反应,切开极性相同的两 条单链,拆分Holliday 中间体。
物种生存具有重要意义。通过重组实现基因的重新组合使
物种能够更快地适应环境,加快进化的过程。此外,DNA 重组还参与许多重要的生物学过程,比如重组在DNA损伤 修复和突变中发挥重要作用。
高中生物第十一节:基因重组与基因转位——基因重组概念
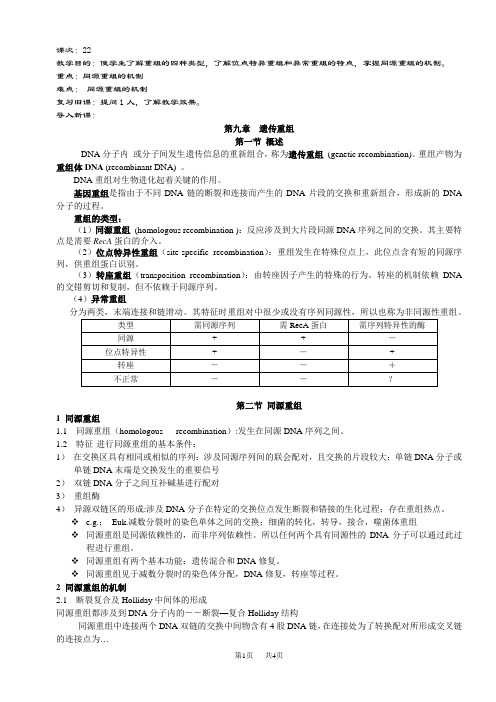
课次:22教学目的:使学生了解重组的四种类型,了解位点特异重组和异常重组的特点,掌握同源重组的机制。
重点:同源重组的机制难点:同源重组的机制复习旧课:提问1人,了解教学效果。
导入新课:第九章遗传重组第一节概述DNA分子内或分子间发生遗传信息的重新组合,称为遗传重组(genetic recombination)。
重组产物为重组体DNA (recombinant DNA) 。
DNA重组对生物进化起着关键的作用。
基因重组是指由于不同DNA链的断裂和连接而产生的DNA片段的交换和重新组合,形成新的DNA 分子的过程。
重组的类型:(1)同源重组(homologous recombination ):反应涉及到大片段同源DNA序列之间的交换。
其主要特点是需要RecA蛋白的介入。
(2)位点特异性重组(site-specific recombination):重组发生在特殊位点上,此位点含有短的同源序列,供重组蛋白识别。
(3)转座重组(transposition recombination):由转座因子产生的特殊的行为。
转座的机制依赖DNA 的交错剪切和复制,但不依赖于同源序列。
(4)异常重组分为两类,末端连接和链滑动。
其特征时重组对中很少或没有序列同源性,所以也称为非同源性重组。
第二节同源重组1 同源重组1.1 同源重组(homologous recombination):发生在同源DNA序列之间。
1.2 特征-进行同源重组的基本条件:1)在交换区具有相同或相似的序列:涉及同源序列间的联会配对,且交换的片段较大;单链DNA分子或单链DNA末端是交换发生的重要信号2)双链DNA分子之间互补碱基进行配对3)重组酶4)异源双链区的形成:涉及DNA分子在特定的交换位点发生断裂和错接的生化过程;存在重组热点。
e.g.:Euk.减数分裂时的染色单体之间的交换;细菌的转化,转导,接合,噬菌体重组同源重组是同源依赖性的,而非序列依赖性。
同源重组法分子克隆 -回复

同源重组法分子克隆 -回复同源重组法是分子克隆技术中的一种重要方法,其基本原理是利用DNA的同源性重组来插入外源DNA序列到宿主DNA中。
同源重组法在基因克隆、遗传工程等领域得到了广泛应用。
本文将详细介绍同源重组法的原理、步骤及应用。
一、同源重组法的原理同源重组法的原理基于DNA分子的自身结构和功能,DNA分子在某些条件下能够进行重组、修复和重复。
同源重组是指两个DNA分子之间具有相似序列(同源)的区域进行交换而形成的DNA分子重组。
同源重组法基于此原理,通过在宿主DNA中引入重组的同源片段,将外源DNA序列插入到宿主DNA中。
同源重组法的原理可以分为两个步骤:相互间接断裂和互补配对。
两个DNA分子的同源片段同时发生间接断裂,获得可供基因重组的末端。
接下来,由于互补配对的作用,从两个DNA分子中间的同源片段在一定条件下进行配对,形成插入、缺失、互换等不同类型的重组产物。
1. 构建载体DNA:载体DNA是将外源DNA插入到宿主DNA中的重要工具,构建载体DNA 需要选择有适当限制酶切位点的载体和外源DNA。
一般来说,常用的载体包括质粒、噬菌体、噬菌体样颗粒等。
2. 制备DNA片段:外源DNA片段可以通过PCR扩增、酶切和DNA合成等技术制备。
需要注意的是,PCR扩增要确保扩增的DNA片段与宿主DNA具有一定的同源性。
3. 利用限制酶切割载体和外源DNA:根据预定的酶切位点设计限制酶切位点并进行酶切。
4. 进行杂交和拼接:将外源DNA片段与载体DNA杂交,并通过互补配对将DNA片段与载体DNA进行拼接。
5. 转化大肠杆菌:利用化学方法或电击法将构建好的载体DNA转化到大肠杆菌中,转化后得到含外源DNA的菌落。
6. 筛选阳性菌落:利用选择性培养基和荧光素酯分析方法等技术筛选阳性菌落。
7. 测序鉴定:对筛选出的阳性菌落进行测序,并鉴定插入的外源DNA序列是否正确。
同源重组法是分子克隆领域中一种非常实用的技术。
同源重组技术的原理和应用

同源重组技术的原理和应用同源重组技术(Homologous recombination technology,HRT)是一种常用的基因编辑技术,它能够在特定部位改变DNA序列,用于治疗某些遗传性疾病、研究基因表达调控和蛋白质结构等方面。
本文将介绍同源重组技术的原理和应用。
1. 同源重组技术的原理同源重组技术是利用质粒、病毒等载体携带的外源基因通过靶向指向的方式将其导入到细胞或生物体中,从而达到改变生物体基因组的目的。
具体来说,同源重组技术是基于DNA的相互作用原理进行的。
DNA由四种碱基(腺嘌呤、胸腺嘧啶、鸟嘌呤和胞嘧啶)构成,它们之间形成了氢键,使得两条互补的DNA链可以通过这些氢键相互结合。
在同源重组中,DNA分子的另一端则通过DNA酶和锌指核酸酶来实现切割和精准定位。
一旦发现了具备相同序列的区域,这些酶就会将外源基因定向到位点,与染色体上的同源序列组成DNA双链,从而取代了原有的序列,以达到修复或替换某些基因的效果。
2. 同源重组技术的应用同源重组技术的应用广泛,其中最重要的是对基因的编辑和修复。
以下将介绍几种常见的同源重组技术应用:(1)质粒介导同源重组质粒介导同源重组是一种常用的基因工程技术。
这种技术主要是利用菌单倍体的同源重组能力,通过转化质粒来实现有选择性地在DNA的特定区域插入新基因。
这种技术特别适用于菌类以及一些单细胞真菌和原生生物。
(2)病毒介导同源重组病毒介导同源重组则运用了病毒自身的重组机制特征,对其进行了改造,使其能够以带选择性的方式在目标细胞中整合外源基因。
这种方法目前已经广泛应用于人类基因治疗领域,尤其在修复致病基因和引入新基因方面取得了显著进展。
(3)基因组编辑基因组编辑技术可以通过同源重组来治疗遗传性疾病。
比如,在用于治疗Friedreich's ataxia(FA)的基因治疗研究中,研究团队采用基因克隆技术构建了能够靶向FA基因的重组质粒,并通过同源重组的方式将其导入细胞中。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
哈工大-遗传学
第四章 同源重组的分子机制
断裂重接模型和模板选择复制模型存在的问题
断裂重接模型:不能解释基因转变和极化子现象; 模板选择复制模型:
(1) 违背了半保留复制; (2) DNA复制在S期,重组在偶线期,不应同时发生; (3)不能解释3线和4线交换;
哈工大-遗传学
第六章 同源重组的分子机制
3 + pdxp + + pdx + + +
4 pdx + + pdxp pdx +
哈工大-遗传学
第六章 同源重组的分子机制
pdx +
--
-+
-+
-+
++
++
+-
+-
-+
-+
-+
基因转-变+(gene conver-sio-n):一个基因-转+变为它的等位基
因的遗+传+学现象(源于+ 基+ 因内重组)。+ +
上一课重点内容回顾
• 连锁与交换规律
两点测交 三点测交
**相关概念:交换值、重组率、遗传干涉、并发系数、 染色单体干涉等
• 四分子遗传分析
着丝粒作图 两个连锁基因作图
**相关概念:顺序四分子、第一次分裂分离(MI模式)、
第二次分裂分离(MII模式)等
上一课重点内容回顾
着丝粒作图原理与方法
顺序四分子遗传分析
3. 切开的单链交换重接;
4. 形成交联桥结构; 5. 分支迁移:形成一大段异源 4
双链DNA(Holliday结构)
5
7
6. 绕交联桥旋转1800,形成
Holliday异构体;
7. 切开与重接:
左右切,形成非重组体
上下切,形成重组体。
支持Holliday遗传重组模型的证据: 1、形态学上,Holliday中间体(Chi 结构)的电镜照片;
例:同源染色体非姐妹单体交换; 细菌的转化、转导、接合; 噬菌体的重组…
条件:2个DNA分子序列同源;
不同物种同源重组所需的最小的同源序列不同;
哈工大-遗传学
同源重组的分子机制
解释同源重组分子机制的模型:
Байду номын сангаас
断裂和重接模型 1937年,Darlington提出;
模板选择复制模型(copy choice ) Belling J.首先提出,1933年又撤回; 1948年,Hershey再次提出;
2、基因转变的分子机制
①在5:3和6:2分离的子囊中,约有30%也在g基因两侧 发生重组;
②发生基因转变的子囊中,基因转变和遗传重组都发生 于同样的两个染色单体上的子囊的比例高达90%;
基因转变的实质:重组过程中留下的局部异源双 链区,在细胞内的修复系统识别下不同的修复产 生的结果;
哈工大-遗传学
Holliday 模型中,由于重组而产生的异源双链区存在 不配对碱基,可被细胞内的修复系统能够识别并以一条链 为模板进行切除修复:
(1). 两个杂种分子均未得到校正,有丝分裂后分离形成4:4 或 3:1:1:3的异常孢子分离比,属于半染色单体转变;
(2). 一个杂种分子得到校正,另一条未校正,有丝分裂后 分离形成 5:3 或 3:5 的分离比,属于半染色单体转变;
1、异常分离与基因转变 粗糙脉孢菌: pdxp:酸度敏感的VB6依赖型 pdx: 酸度不敏感的VB6依赖型
哈工大-遗传学
第六章 同源重组的分子机制
+ pdxp
× pdx +
585个子囊中
孢子对
①
子囊
②
③
④
1 2
? + pdxp pdx +
+ + pdx +
+ + pdx + + pdxp + pdxp
+-
+-
+-
-+
-+
-+
-+
+-
++
+-
+-
染色单体转变粪生粪壳菌(Oliv半e)染: 色g+单×体g转- 变
+
+
+
+
+
+
+
+
+
-
-
-
+
+
+
+
+
+
+
?
-
-
-
-
6:2 (或 2:6)
120/200000
5:3 (或 3:5)
100/200000
+
+
+
+
?
-
+
?
-
-
-
-
3:1:1:3
16/200000
霍利迪(Holliday)模型 1964年Robin Holliday提出;
Meselson-Radding模型 1975年Meselson Radding提出;
哈工大-遗传学
同源重组的分子机制
1
2.
断裂重接模型
3.
4.
1.
2.
3.
4.
模板选择复制 模型
图 23-3 断裂重接模型和模板选择复制模型的比较
四分子分析
两个基因作图原理与方法
非顺序四分子遗传分析
相关概念:四分子、顺序四分子、第一次分裂分离(MI模式)、 第二次分裂分离(MII模式)、子囊型(PD、NPD和T)
第六章 同源重组的分子机制
哈工大-遗传学
第六章 同源重组的分子机制
一、概念
同源重组:依赖大范围的DNA同源序列的联会,重组过 程中,两个染色体或DNA分子交换对等的部分。
第六章 同源重组的分子机制
哈工大-遗传学
第六章 同源重组的分子机制
支持Holliday遗传重组模型的证据: 1、形态学上,Holliday中间体(Chi 结构)的电镜照片; 2、异源双链形成时,产生碱基错配导致与重组相关联 的基因转变的发生;
哈工大-遗传学
第六章 同源重组的分子机制
(二)、基因转变及其分子机理
二、Holliday模型
依据:基因转变 证据:细菌重组chi结构
哈工大-遗传学
第六章 同源重组的分子机制
(一)、过程
1
1. 同源的非姐妹染色单体联会;
6
2. DNA内切无酶论在非H姐o妹ll染id色a单y 结2构断裂是否导致旁侧遗传标
体的相同位置同时切开两个
旋转1800
方记向的相同重的组一条,单它链们; 都含有3 一个异源双链区。
第六章 同源重组的分子机制
哈工大-遗传学
第六章 同源重组的分子机制
左右切
异源双链区
D/d
A
G
B
A
A
B
a
C
b
a
T
b
D/d
上下切
异源双链区
D/d
A
G
b
A
A
b
a
T
B
a
C
B
D/d
A
G
B
A
A
B
a
C
b
a
T
b
A
G
b
A
A
b
a
T
B
a
C
B
哈工大-遗传学
第六章 同源重组的分子机制
哈工大-遗传学
第六章 同源重组的分子机制
(3). 两个杂种分子都被校正到同一种类型,有丝分裂后分 离形成 6:2或2:6 的分离比,属于染色单体转变;
(4). 两个杂种分子都被校正到不同类型,有丝分裂后分离 形成 4:4 分离比,未出现基因转变;
哈工大-遗传学
第六章 同源重组的分子机制
Holliday模型同样适用于环状DNA间的重组 两个环状DNA分子配对、断裂、重接形成“8”字
型结构中间物,根据切割的位置不同,可分别形成两 个亲本环、大的单体环或者是滚环结构,也可以形成 “χ” 结构。
哈工大-遗传学
第六章 同源重组的分子机制
2
哈工大-遗传学