近10年来厦门市湿地动态变化分析
厦门马銮湾北部湿地生态恢复与重构技术的研究

林业勘察设计
( 福建 ) 2 0 0 7年第 1 期
首要因 素 ; 海湾 周边 农 田 的化肥 和磷 流失 人湾 也是 重 要 污染 源 。据测 算 , 流人 湾 内的 总氮 量为 77 a磷 1t , / 为 01 a .t 。此 外 , / 马銮 湾 还需 接纳 杏林 工 业 区废 水 4 5万 ta / 。据 国家海 洋局 第 三 研究 所 19 96年 3月 监
湿地在抵御洪水、 调节径流、 控制污染 、 调节气候、 美化环境等方面起到重要作用, 它既是陆地上 的天然蓄
水库 , 又是众 多野生 动植物 资源 ( 特别 是 珍稀 水 禽 ) 繁殖 和越 冬地 。湿 地 与人 类 息息 相关 , 人类 拥 有 的 是 的宝 贵资源 , 湿地 被人们称 为“ 生命 的摇 篮 ” “ 球 之 肾” “ 、地 和 鸟类 的乐 园 ” 。厦 门马 銮湾湿 地 曾经 是个 美 丽富饶 的天 然湿地 , 目前其 水生 生态 环境 已经受到 严重 的破 坏 , 日益 发展 的经济环境 很不相 适应 。根 但 与 据 《 门市加 快海湾 型城市 建设 实 施 纲 要 》 马銮 湾地 区将作 为 厦 门 市 今后 重 点 建设 的 6个 主要 城 区之 厦 ,
右, 其余水域成为水产养殖地 。重构区水域为虾池鱼塘 , 供水产养殖。 l 溪流 : l4 2 马銮 湾区域 内柏瑶 山溪 、 青 溪 、 溪 、 芸溪 、 头溪 、 溪 、 溪 、 溪 、 头溪 深 后埔 过 埭 祥露 新墙 林东 船
等 9条溪 流 ( q新j 、 东溪 、 其 安溪 林 船头 溪 由于新 阳东区 的开发建 设而 被改 造 成排 洪 明渠 ) 分别 发源 于两 ,
甚 至 1 m 的流塑状 淤泥 。 2 1 1 3 土壤 ..
【地方试题】2024年厦门市初中毕业年级模拟考试地理试题(5月)

2024年厦门市初中毕业年级模拟考试地理本试卷共8页。
满分100分。
第I卷选择题本卷共25小题,每小题2分,共50分。
在每小题给出的四个选项中,只有一项是符合题目要求的。
“东山陆桥”是台湾海峡中的一条浅滩,在过去某个时期曾出露海面,直到7000年前,因冰川消融,被海水淹没。
图1示意台湾海峡深线等深线分布。
读图,完成1~2题。
图11.现代“东山陆桥”最深处海拔约为A.-30米B.-50米C.-70米D.-90米2.“东山陆桥”被海水淹没的原因可能是A.海面升降B.板块碰撞C.植被变化D.人类活动【参考答案】1.C 2.A2024年2月7日,中国第五个南极科考站——秦岭站顺利开站,填补了中国在南极罗斯海区域的考察空白。
秦岭站是一个集约高效、绿色低碳、智能先进的常年科考站。
图2为秦岭站照片。
读图,完成3~5题。
图23.秦岭站建筑主体设计时,较少考虑A.墙体保温B.风沙侵蚀C.建筑防火D.积雪掩埋4.为践行“绿色低碳"理念,秦岭站的供能适宜选择A.水能和煤炭B.水能和太阳能C.风能和煤炭D.风能和太阳能5.南极地区秦岭站的建设有助于A.捕捞海洋生物B.开采矿产资源C.改变生态环境D.开展气候研究【参考答案】3.B 4.D 5.D冷流降雪是高空强冷空气经过海洋,与来自海面的暖湿气流交汇后产生的降雪。
2023年12月20日,我国山东半岛多地遭遇冷流降雪。
图3示意山东半岛地形及本次冷流降雪的降雪量分布。
读图,完成6~8题。
图36.本次冷流降雪过程中,烟台的风力和风向是A.7级东南风B.7级西北风C.6级偏南风D.6级偏北风7.山东半岛冷流降雪的水汽主要来自A.渤海和黄海B.渤海和东海C.黄海和东海D.东海和南海8.烟台冷流降雪量远大于青岛的主要原因是A.人类活动频繁B.地形阻挡C.距离海洋更近D.纬度更高【参考答案】6.B 7.A 8.B澳大利亚地广人稀,人口老龄化严重。
图4示意澳大利亚人口分布。
读图,完成9~11题。
厦门市气候变化及其应对建议
厦门市气候变化及其应对建议作者:苏明峰来源:《海峡科学》2011年第07期[摘要]厦门正处于经济快速发展时期,应对气候变化形势严峻,任务艰巨。
该文在分析厦门市气候变化的事实和影响的基础上,提出厦门市应对气候变化的几点建议。
[关键词]厦门气候变化应对建议当前,气候变化正对自然生态系统和人类生存环境产生重大影响,气候变化所导致的气温增高、海平面上升、极端天气气候事件频繁发生等,受到国际社会的普遍关注。
气象观测数据表明:在全球变暖的大背景下,厦门地区气候也发生着明显的变化。
厦门正处于经济快速发展阶段,应对气候变化事关经济发展全局和人民群众的切身利益,如何应对并减缓气候变化对厦门经济、社会和生态环境的影响是当前乃至将来很长一段时间内的重要任务。
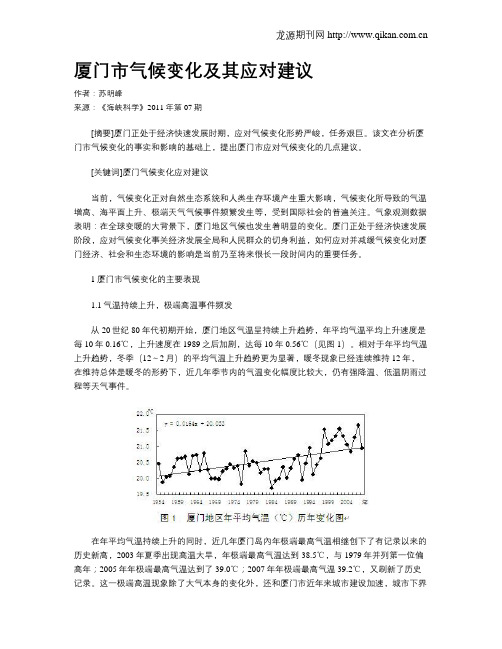
1厦门市气候变化的主要表现1.1气温持续上升,极端高温事件频发从20世纪80年代初期开始,厦门地区气温呈持续上升趋势,年平均气温平均上升速度是每10年0.16℃,上升速度在1989之后加剧,达每10年0.56℃(见图1)。
相对于年平均气温上升趋势,冬季(12~2月)的平均气温上升趋势更为显著,暖冬现象已经连续维持12年,在维持总体是暖冬的形势下,近几年季节内的气温变化幅度比较大,仍有强降温、低温阴雨过程等天气事件。
在年平均气温持续上升的同时,近几年厦门岛內年极端最高气温相继创下了有记录以来的历史新高,2003年夏季出现高温大旱,年极端最高气温达到38.5℃,与1979年并列第一位偏高年;2005年年极端最高气温达到了39.0℃;2007年年极端最高气温39.2℃,又刷新了历史记录。
这一极端高温现象除了大气本身的变化外,还和厦门市近年来城市建设加速,城市下界面改变造成的热岛效应有关。
城市下界面大面积水泥化使得厦门市区夏季最高气温要比周边郊区高2~3℃,城市热岛效应明显。
1.2 暴雨、干旱和强台风等极端天气气候事件增多尽管过去半个多世纪厦门年降水总量只有轻微的增加趋势,但极端降水事件却明显增多。
厦漳泉地区生态系统服务权衡与协同关系分析

厦漳泉地区生态系统服务权衡与协同关系分析胡其玉,陈松林(福建师范大学a.福建省亚热带资源与环境重点实验室;b.地理研究所,福州350007)摘要:将地理探测器引入生态系统服务权衡/协同关系的研究,并结合生态服务价值动态度、生态系统服务权衡协同度(ESTD),阐释2006—2016年厦漳泉地区生态系统服务变化特征和权衡/协同关系及其时空格局。
结果表明:①地区生态服务价值总体增长16.51%,其中林地、湿地、水域的生态服务价值增长明显;②食物生产、原料生产、土壤保持服务价值低于地区总体ESV动态度;③生态系统服务间作用强度会随对象的转换出现差异性,根据各类服务间作用的紧密程度划分核心和次核心“组团”;④地区生态系统服务间关系以协同为主,但食物生产与其他服务呈现明显的此消彼长,不同生态系统服务间权衡/协同性质及其程度均呈显著的空间分异。
关 键 词:生态系统服务;权衡与协同;地理探测器;厦漳泉地区中图分类号:F062.2;X171.1 文献标志码:A 文章编号:10032363(2021)02?0145?06doi:10.3969/j.issn.1003?2363.2021.02.026收稿日期:2019-02-14;修回日期:2021-03-05基金项目:国家自然科学基金项目(41771136);福建省科技重大专项(2012NZ0002)作者简介:胡其玉(1993-),男,四川成都市人,硕士研究生,主要从事土地利用评价研究,(E mail)zhongguo999long@qq.com。
通信作者:陈松林(1964-),男,福建南安市人,教授,博士,主要从事土壤地理与土地资源研究,(E mail)slchen6@163.com。
0 引言 生态系统服务就是人类通过生态系统获取生产生活所需的各种收益[1]。
随着人类社会的快速发展,对生态系统结构和功能的扰动愈加强烈,因而需要开展生态系统管理。
而生态系统服务类型和驱动因素的多样性和各类服务间错综复杂的联系,解析多重生态系统服务间动态变化,即权衡与协同,对协调多种生态系统服务、实现多目标管理具有重要意义。
潮汐的转变案例分析

潮汐的转变案例分析潮汐的转变案例分析潮汐是大洋中引起海面高度周期性变化的现象,是由地球引力和惯性力引起的。
潮汐的转变是指海洋潮汐系统在某一地点的周期性变化与时间和空间的关系。
下面将对潮汐的转变进行案例分析。
潮汐的转变会受到多种因素的影响,包括月球和太阳的引力、地球自转的惯性作用、地形、气象等。
在具体的案例中,我将以中国厦门地区的潮汐转变为例进行分析。
厦门位于中国福建省东南部,背靠大陆,并与台湾隔海相望。
厦门地区的潮汐转变是受到月球和太阳的引力影响最为显著。
太阳和月球的引力是潮汐形成的主要原因之一,月球引力对潮汐影响远大于太阳引力,因为月球相对地球较近。
根据厦门的地理位置和潮汐转变规律,我们可以看到,厦门的潮汐周期大约为12小时25分钟。
这个周期是由月球绕地球公转一周所需时间决定的。
根据月球的周转,厦门的潮汐会有两次高潮和两次低潮。
在一天之内,潮汐会发生两次涨潮两次退潮的变化。
另外,在厦门地区,由于地形的影响,潮汐高低不均匀。
厦门地区有许多河道和浅滩,这些地理特征会引起潮水流速的变化,使潮汐高度不同。
同时,由于厦门地区是一个海岛城市,岛屿与海岸之间的地形也会对潮汐转变产生影响。
在岛屿上,由于海洋与陆地之间的夹角的变化,潮汐的高度也会有所不同。
最后,气象因素也会对潮汐转变产生影响。
例如,风力的大小和方向会改变潮汐的流向和速度,从而影响潮汐的转变。
此外,海洋表面的气压变化也会影响潮汐的高度。
综上所述,潮汐的转变是一个复杂的过程,受到多种因素的影响。
就厦门地区的潮汐转变来说,月球和太阳的引力、地形、气象等是影响潮汐转变的主要因素。
通过对潮汐转变的分析,我们可以更好地理解潮汐形成的原理和规律,为海洋资源的开发和利用提供一定的依据。
厦门同安湾下潭尾人工红树林湿地小型底栖动物群落结构

厦门同安湾下潭尾人工红树林湿地小型底栖动物群落结构陈昕韡;李想;曾佳丽;谭文娟;周细平;洪万树;蔡立哲【摘要】为研究厦门同安湾下潭尾人工红树林湿地小型底栖动物的群落结构,于2014年在下潭尾人工红树林湿地5个取样站进行了4个季节小型底栖动物定量取样,共获取了9个小型底栖动物类群,分别为自由生活海洋线虫、底栖桡足类、寡毛类、多毛类、涡虫类、有孔虫类、介形类、双壳类、星虫类,还有少许未定类群.其中,自由生活海洋线虫是优势类群,占总丰度的91.75%.下潭尾人工红树林湿地小型底栖动物平均丰度为(441.3±61.0) ind/(10 cm2),平均生物量为(555.8±104.6) μg/(10 cm2).单变量双因素方差分析(two-way ANOVA)表明:不同季节之间小型底栖动物丰度和生物量有极显著差异;不同取样站之间小型底栖动物丰度无显著差异,生物量有显著差异.小型底栖动物群落的类群均匀度指数(J')、多样性指数(H')和优势度指数(λ)的最高值均出现在光滩取样站.Pearson相关性分析表明:小型底栖动物的个体数与底温呈极显著负相关、与底盐呈显著负相关;夏、秋两季小型底栖动物的类群数以及夏季小型底栖动物丰度均与底盐呈显著相关.上述结果完善了我国人工红树林湿地小型底栖动物物种和生境多样性资料库,为滨海湿地公园管理、滩涂生态修复和红树林湿地管理提供了基础资料.%In order to study the community structure of meiofauna in artificial mangrove wetland of Xiatanwei located in Tong'an Bay,the meiofauna was quantitatively investigated at five stations in the four seasons of 2014.Nine meiofaunal groups were observed,including free-living marine Nematoda,benthic Copepoda,Oligochaeta,Polychaeta,Turbellaria,Foraminifera,Ostracoda,Bival via and Sipuncula.Among them,free-living marine Nematoda was the dominant group,accounting for 91.75 % of the total abundance.Theaverage abundance and biomass of meiofauna were (441.3 ± 61.0) ind/(10 cm2) and (555.8 ± 104.6) μg/(10 cm2),respectively.Univariate two-way ANOVA showed that there were highly significant differences in meiofauna abundance and biomass among seasons,and significant differences in biomass but not in meiofauna abundance among stations.The highest values of evenness index (J'),Shannon-Wiener index (H') and dominance index (λ) all appeared in the mudflat station.Meanwhile,Pearson correlation analysis indicated that temperature and salinity were the factors influencing the group number of meiofauna,and salinity was a crucial element that affected the abundance of meiofauna in summer.This study provides new data bank of meiofauna in artificial mangrove areas in China and supplies basic data of coastal wetland park management,intertidal zone ecological restoration and coastal mangrove wetland management.【期刊名称】《厦门大学学报(自然科学版)》【年(卷),期】2017(056)003【总页数】8页(P351-358)【关键词】小型底栖动物;群落;人工红树林湿地;同安湾;下潭尾【作者】陈昕韡;李想;曾佳丽;谭文娟;周细平;洪万树;蔡立哲【作者单位】厦门大学环境与生态学院,福建厦门361102;厦门大学环境与生态学院,福建厦门361102;厦门大学环境与生态学院,福建厦门361102;厦门大学环境与生态学院,福建厦门361102;厦门大学嘉庚学院环境科学与工程学院,福建漳州363105;厦门大学海洋与地球学院,福建厦门361102;厦门大学环境与生态学院,福建厦门361102【正文语种】中文【中图分类】Q178.1同安湾位于福建省东南部沿海厦门岛北侧,东接翔安区,西、北部分别为同安区和集美区,南部为厦门岛,包括东咀港和浔江港海域[1].下潭尾海域位于同安湾顶,属于厦门市翔安区范围.有关同安湾红树林湿地底栖动物的研究,开始于集美凤林老红树林湿地(因城市建设已被填埋)大型底栖动物体内多环芳烃的研究[2]和大型底栖动物群落的研究[3],随后比较了同安湾潮间带红树林生境与非红树林生境的大型底栖动物群落[4],并进行了同安湾红树林树上大型底栖动物生态分布的研究[5].同安湾红树林湿地小型底栖动物的研究开始于2008年,主要报道了凤林红树林湿地自由生活海洋线虫群落[6],还比较了同安湾两处红树湿地的小型底栖动物丰度[7].对同安湾潮下带底栖动物的研究较早:2000年,方少华等[8]报道了浔江湾小型底栖生物数量;2007年,林俊辉等[9]报道了同安湾春季大型底栖生物的群落结构特征.小型底栖动物作为连接有机碎屑、初级生产和水层-底栖耦合的重要环节,是许多经济鱼、虾和贝类幼体时期的优质饵料[10];同时,小型底栖动物也是沉积物中有机碎屑的开发者和底栖细菌、微藻的主要消耗者,其摄食率大体与微生物生产量持平,调节微生物生产过程[11],在海洋生态系统中起着重要作用.2011年,厦门市在下潭尾海域启动了以红树林为主题的生态湿地公园建设,种植红树林42.8 hm2.本研究拟了解新的人工红树林湿地小型底栖动物动物群落结构以及主要环境因子对其的影响,旨在为我国人工红树林湿地建设和红树林湿地生态恢复提供基础资料.1.1 研究区域概况厦门市下潭尾滨海生态湿地公园位于厦门市翔安区下潭尾片区,南北以现有海岸线为界,东至塘厝水闸坝堤,西至东坑湾水道东侧口与赵厝岸线西侧连线,公园总规划面积400 hm2,其中滩涂面积125 hm2.已开工建设的下潭尾滨海生态湿地公园计划构建适宜红树林生长的人工滩涂岛5个,面积约44.8 hm2,滩涂治理面积15.5 hm2[12].红树林种植工程已于2012—2013年基本完成,种植树种以秋茄(Kandelia candel)为主,还包括桐花树(Aegiceras corniculatum)、白骨壤(Avicennia marina)和无瓣海桑(Sonneratia apetala)等.本研究在下潭尾人工红树林湿地布设A、B、C、D、E共5个取样站(图1),并于2014年2月(冬季)、5月(春季)、8月(夏季)、11月(秋季)进行了4个季节小型底栖动物的生态调查,每个取样站各取5个平行样.A、B、C、E取样站位于人工红树林内,D取样站位于光滩.1.2 样品采集和处理方法小型底栖动物的样品采集在低潮时进行.选取表面较平整且未受扰动的区域作为采样点,用内径为2.9 cm的注射器改造而成的采样管取样,取样深度约为9 cm.将采样管中的沉积物转移到250 mL塑料广口瓶,然后加入5%(体积分数)的甲醛溶液进行固定,固定后的所有样品带回实验室分选.分选前,先将样品摇匀;然后,将0.500 mm和0.042 mm孔径的网筛叠放,将摇匀的样品倒入网筛,用自来水冲洗样品,除去样品中的黏土、粉砂及其他杂质;接着,将0.042 mm孔径的网筛上残留的沉积物样品用密度为1.15 g/mL的Ludox-TM硅胶液分次转移到离心管中,Ludox-TM硅胶液的体积约为沉积物的3~4倍.混匀后,将样品以5 000 r/min的转速离心5 min.取悬浮液,向残留沉积物的离心管中再次加入Ludox-TM硅胶液重复离心1次.合并2次离心所得的悬浮液,倒入0.042 mm孔径的网筛,用自来水冲洗去除Ludox-TM硅胶液.之后,将样品转移到带平行线的培养皿中,在SMZ-168体式显微镜下对小型底栖动物进行分类、计数,将计数完的小型底栖动物保存于75%(体积分数)的乙醇溶液中以待后续操作.小型底栖动物的丰度(ind/(10 cm2),单位面积个体数)由直接计数得到.小型底栖动物的生物量(μg/(10 cm2),单位面积干重)测定方法采用换算法,即通过小型底栖动物各类群的个体平均干重乘以相应类群的丰度得到.自由生活海洋线虫(free-living marine Nematoda,简称线虫)、底栖桡足类(benthic Copepoda,简称桡足类)、寡毛类(Oligochaeta)、多毛类(Polychaeta)和介形类(Ostracoda)平均个体干重的测定方法以《海洋调查规范第6部分:海洋生物调查GB/T 12763.6—2007》[13]为依据,上述5个类群之外其他类群的平均个体干重参照Jario[14]、Widbom[15]和张志南等[16]的研究结果,见表1.2.1 小型底栖动物的类群组成2014年2—11月的4个季节中,共在下潭尾人工红树林湿地采集到9个类群的小型底栖动物,分别为线虫、桡足类、寡毛类、多毛类、涡虫类、有孔虫类、介形类、双壳类和星虫类,还有少许未定类群归为其他类.采集到的小型底栖动物中,线虫丰度百分比最高(91.75%),双壳类丰度百分比最低(0.02%),其他7个类群按丰度百分比由高到低依次为:桡足类(4.52%)、寡毛类(2.13%)、有孔虫类(0.57%)、涡虫类(0.45%)、多毛类(0.27%)、星虫类(0.17%)、介形类(0.05%),另其他类占0.07%.分析各取样站的不同类群丰度百分比,结果如图2所示:在5个取样站均有发现线虫、桡足类、寡毛类、多毛类、涡虫类、有孔虫类和星虫类7个小型底栖动物类群,其中线虫的丰度百分比在各取样站均达到了80%以上;A、D和E取样站丰度百分比居第二位的是桡足类,而B、C取样站为寡毛类.A取样站共采集到8个小型底栖动物类群,除上述7个共有类群外,增加了其他类;在B、C、D取样站均采集到9个小型底栖动物类群,除上述7个共有类群外,增加了介形类和双壳类,还有少许未定类群即归为其他类;E取样站获得8个小型底栖动物类群,除上述7个共有类群外,增加了介形类.利用PRIMER 5.0软件对下潭尾人工红树林湿地小型底栖动物的类群组成进行多维尺度分析(multi-dimensional scaling,MDS).分析结果(图3)显示:Stress=0.09,表明图形中显示的样本间关系可信;从季节上看,冬季小型底栖动物的类群组成最相似,夏季的差异最大.2.2 小型底栖动物的平均丰度和平均生物量分析各季节各取样站的小型底栖动物丰度,结果如图4所示:丰度最高值出现在冬季的E取样站,为(855.9±325.1) ind/(10 cm2);最低值则出现在夏季的D取样站,为(190.3±60.1) ind/(10 cm2).从全年平均值上看:E取样站小型底栖动物的丰度最高,为(514.0±282.5) ind/(10 cm2);B、C、A取样站依次递减,分别为(471.5±254.1) ind/(10 cm2),(464.7±182.2) ind/(10 cm2)和(388.6±129.1) ind/(10 cm2);D取样站最低,为(368.0±164.0) ind/(10 cm2).春季小型底栖动物的丰度最高值出现在B取样站,夏季丰度最高值出现在C取样站,春、夏两季丰度最低值均出现在D取样站;秋、冬两季丰度最高值均出现在E取样站,秋季最低值出现在B取样站,冬季最低值出现在A取样站.单变量双因素方差分析(two-way ANOVA)结果表明:不同季节间小型底栖动物的丰度呈极显著差异(p<0.01),不同取样站间小型底栖动物的丰度无显著差异(p>0.05),季节×取样站间小型底栖动物的丰度无显著差异(p>0.05).分析各季节各取样站的小型底栖动物生物量,结果如图5所示:生物量的最高值出现在秋季的E取样站((1 015.6±33.5) μg/(10 cm2)),最低值出现在夏季的B取样站((168.6±26.3) μg/(10 cm2));从全年平均值上看:E取样站小型底栖动物的生物量最高,为(709.9±315.9) μg/(10 cm2);B、C、D取样站依次递减,分别为(575.3±311.8) μg/(10 cm2),(548.8±258.8) μg/(10 cm2)和(526.1±211.4)μg/(10 cm2);A取样站最低,为(419.1±106.3) μg/(10 cm2).春季小型底栖动物的生物量最低值出现在D取样站,最高值出现在B取样站;夏季生物量最低值出现在B取样站,秋、冬两季最低值均出现在A取样站,夏、秋、冬三季最高值均出现在E取样站.Two-way ANOVA结果表明:不同季节间小型底栖动物的生物量呈极显著差异(p<0.01),不同取样站间小型底栖动物的生物量呈显著差异(p<0.05),季节×取样站间小型底栖动物的生物量无显著差异(p>0.05).2.3 小型底栖动物类群的多样性利用PRIMER5.0软件计算下潭尾人工红树林湿地各季节各取样站小型底栖动物的类群数(S)、个体数(N)、类群丰富度指数(d)、均匀度指数(J′)、香农-威纳多样性指数(H′)和优势度指数(λ).计算结果如表2所示:d的最高值出现在秋季的B取样站(0.825);J′、H′和λ的最高值均出现在夏季的D取样站,分别是0.443,0.891和0.330;d、J′、H′和λ的最低值均出现在夏季的B取样站,分别是0.134,0.104,0.069和0.018.2.4 小型底栖动物类群与环境因子的相关性将下潭尾人工红树林湿地4个季节小型底栖动物的类群数和丰度与底温、底盐进行Pearson相关性分析,结果(表3)表明:夏季的类群数与底盐呈显著负相关(p<0.05);秋季的类群数与底盐呈显著正相关(p<0.05);夏季的丰度与底盐呈显著正相关(p<0.05);其他各参数之间无显著相关性.对下潭尾人工红树林湿地小型底栖动物的类群的多样性参数与底温、底盐进行Pearson相关性分析,结果(表4)表明:个体数与底温呈极显著负相关(p<0.01),与底盐呈显著负相关(p<0.05);其余各参数之间无显著相关性(p>0.05).3.1 底温和底盐对小型底栖动物的影响小型底栖动物的分布受物理、化学和生物方面多种因素的影响,如气温、沉积物粒度、泥温、底盐、叶绿素a含量、水深以及自身繁殖特点等.Palmer等[17]的研究显示小型底栖动物的生殖和发育都与温度呈正相关;吴辰[18]研究了湛江高桥红树林的小型底栖动物,发现春、夏、秋三季小型底栖动物丰度与温度有显著相关性;卓异[19]发现泉州湾潮间带红树林区夏季小型底栖动物的丰度与温度有极显著负相关性;而本研究中各季节小型底栖动物的丰度与底温均无显著相关性,这与上述学者的研究结果不同.Palmer等[17]认为冬季水体的底温降到全年较低水平,在经过秋季小型底栖动物的丰度高峰期后,沉积物中有机质消耗殆尽,因此小型底栖动物的丰度为全年较低,吴辰[18]在湛江高桥的研究结果支持以上结论.但本研究的结果却是冬季小型底栖动物的丰度较高,与文献[19-21]的研究结果一致.这可能是因为影响小型底栖动物丰度和分布的因素除了温度外,还有盐度、光照、降水量等.同时,气候条件、红树植物类型、人为扰动程度和沉积物等因素的差异也会造成小型底栖动物丰度的差异.Ingole等[22]研究了盐度对热带河口砂质潮间带小型底栖动物结构的影响,结果表明小型底栖动物的丰度波动与盐度波动有显著相关性,无脊椎动物群落分布与不同的盐度区域有直接的关联性.本研究中夏、秋两季小型底栖动物的类群数与底盐有显著相关性,其中夏季小型底栖动物丰度与底盐呈显著正相关.卓异[19]发现泉州湾潮间带夏季小型底栖动物丰度与盐度呈显著正相关,其他季节各参数与盐度无显著相关性;吴辰[18]在湛江高桥红树林区发现春季小型底栖动物丰度与盐度呈极显著负相关,其他季节均无显著相关性.Ingole等[22]的研究地点为热带的河口砂质潮间带,但上述研究区域为亚热带红树林区.Barbara等[23]的研究显示,砂质沉积物中沉积环境异质性高,许多物种生活于砂间间隙,环境变化时小型底栖动物能迅速响应.因此,在砂质潮间带小型底栖动物对于盐度的变化较为敏感,而由于大多数红树林是生长在细质的淤泥质滩涂上,且一般红树林土壤是初生土壤,土壤由粉粒和黏粒组成,含有大量有机质[24],与砂质潮间带有较大不同,所以红树林区小型底栖动物与盐度的相关性有待进一步研究.3.2 不同红树林区小型底栖动物群落结构的差异由于红树林中具有丰富的单宁酸和有机质,红树林中的小型底栖动物与非红树林区有着显著的差异[25],且小型底栖动物在红树林生态系统中起着非常重要的作用,世界各地的底栖生物学家相继在不同红树林区域开展了研究工作.小型底栖动物的分布很广泛,在海洋任何沉积物中几乎都有分布,但是不同区域和不同生境,其密度、类群组成、生物量和多样性都有非常大的区别[26].我国研究红树林区小型底栖动物的科研人员相对较少,现有的研究区域主要集中在福建、海南、深圳和香港等地;国际上,近10年关于红树林区小型底栖动物的报道主要集中在越南、古巴、印度、加勒比海、红海和西里伯斯海.lafsson[27]、Armenteros等[28]和Sabine[29]的早期研究表明:红树林小型底栖动物主要包括线虫、桡足类、多毛类、寡毛类、猛水蚤类、介形类、动吻类、原足类、涡虫类、纽虫类、海螨类、轮虫类这12 个类群,其中线虫是较主要的类群,其次是桡足类.但近10年国际上对红树林区小型底栖动物的研究[25,28,30]显示,各地区小型底栖动物的类群数为5~12个,除了上述12个类群,还发现了有孔虫类、双壳类、纤毛虫类、刺胞动物、颚口动物、昆虫类、苔藓虫类、肉鞭毛虫类、被囊类、腹足类、甲壳类这11个类群,类群数相对早期研究有了较大的增加.在中国红树林区域的研究中,同样发现了更多的小型底栖动物类群.目前,仅福建红树林区报道的小型底栖动物类群就有20个,分别是线虫、桡足类、多毛类、寡毛类、介形类、动吻类、海螨类、轮虫类、原足类、纽虫类、昆虫类、枝角类、涡虫类、缓步类、有孔虫类、双壳类、原生动物、星虫类、端足类和腹足类[19-21,31].其中,曹婧[20]在漳江口红树林区发现小型底栖动物14类,为福建已报道的区域中类群数较多的;本研究在下潭尾人工红树林湿地鉴定出小型底栖动物9类,低于漳江口红树林地区;而周细平[31]在集美凤林红树林和翔安山后亭红树林均只发现小型底栖动物5类,为福建已报道的区域中类群数较少的.Alongi[32]研究发现砂质海滩小型底栖动物的丰度一般可达(1 000~8 000) ind/(10 cm2),而红树林区则普遍低于500 ind/(10 cm2).但是许多研究显示红树林区小型底栖动物丰度高于500 ind/(10cm2).本研究在同安湾人工红树林湿地获得的小型底栖动物平均丰度为(441.3±61.0) ind/(10 cm2),对比国内报道的(397.6~2 229.9) ind/(10cm2)[6,18-21,31,33]及国外近10年报道的越南、古巴、印度、加勒比海、红海和西里伯斯海等地区的(108.6~2 474) ind/(10 cm2)[25,28,30],处于较低水平.周细平[31]在集美凤林人工红树林区和翔安山后亭人工红树林区只发现小型底栖动物类群5个,远低于本研究区域的9个,但其小型底栖动物年平均丰度(分别是(1 548±512) ind/(10 cm2)和(892±19) ind/(10 cm2))均高于本研究区域((441.3±61.0) ind/(10 cm2)).这可能是因为本研究区域内红树的树龄均不超过2龄,属于新生红树林区,红树凋落物较少,红树林的集约度和郁闭度均低于成熟红树林区,小型底栖动物所处的生态系统尚未完全成熟,为不同类群的生长繁殖提供了可能,同时也制约着小型底栖动物各类群数量的单一增长.另外,有报道指出红树林水域富营养化会造成底栖生物多样性及种类数下降[34];且周边居民频繁采挖红树林中的经济种类,扰动沉积物,影响红树林的生境,同样会导致小型底栖动物的丰度和多样性降低.物种所处环境对其生理机能、分布及种群密度都有极大影响[22].大量资料表明,影响小型底栖动物群落的因素很多,包括气候条件、人为扰动、底质类型、温度、盐度、pH 值、有机物以及一些无机元素的含量等[35],因此造成不同红树林区小型底栖动物群落结构及丰度差异的原因是复杂多样的,难以归结为某种具体的原因,相关研究亟待开展.本研究通过对厦门同安湾下潭尾人工红树林湿地的小型底栖动物定量采集所获数据进行分析,发现影响人工红树林湿地小型底栖动物群落结构的因素复杂多样,且可能与树龄存在一定的关系.这些结果完善了我国人工红树林湿地小型底栖动物物种和生境多样性资料库,为滨海湿地公园管理、滩涂生态修复和红树林湿地管理提供了基础资料.【相关文献】[1] 鲍晶晶,蔡锋,任建业,等.厦门同安湾地貌特征研究[J].应用海洋学学报,2013,32(4):499-508.[2] 蔡立哲,马丽,袁东星,等.九龙江口红树林区底栖动物体内的多环芳烃[J].海洋学报(中文版),2005,27(5):112-118.[3] 林秀春,蔡立哲,马丽,等.厦门凤林红树林湿地大型底栖动物群落[J].生物多样性,2006,14(2):128-135.[4] 周细平,蔡立哲,傅素晶,等.福建同安湾潮间带红树林生境与非红树林生境大型底栖动物群落比较[J].生物多样性,2010,18(1):60-66.[5] 周细平,蔡立哲,傅素晶.福建同安湾红树林树上大型底栖动物的生态分布[J].泉州师范学院学报,2014,32(6):5-9.[6] 郭玉清.厦门凤林红树林湿地自由生活海洋线虫群落的研究[J].海洋学报(中文版),2008,30(4):147-153.[7] ZHOU X P,CAI L Z,FU S parison of meiofaunal abundance in two mangrove wetlands in Tong′an Bay,Xiamen,China[J].Journal of Ocean University ofChina,2015,14(5):816-822.[8] 方少华,吕小梅,张跃平.厦门浔江湾小型底栖生物数量分布及生态意义[J].台湾海峡,2000,19(4):474-477.[9] 林俊辉,柯才焕,郑凤武,等.同安湾春季大型底栖生物的群落结构特征[J].厦门大学学报(自然科学版),2007,46(增1):155-160.[10] 张志南,周红.国际小型底栖生物研究的某些进展[J].中国海洋大学学报(自然科学版),2004,34(5):799-806.[11] 黄勇.南黄海小型底栖生物生态学和海洋线虫分类研究[D].青岛:中国海洋大学,2005:1-2.[12] 李想.同安湾人工红树林湿地小型底栖动物群落研究[D].厦门:厦门大学,2015:5.[13] 国家海洋局.海洋调查规范第6部分海洋生物调查:GB/T 12763.6—2007[S].北京:中国标准出版社,2007.[14] JARIO J V.Nematode species composition and seasonal flucturation of a sublittoral meiofluna community in the German Bight[J].Verff Inst MeeresforschBremerh,1975,15:283-337.[15] WIDBOM B.Determination of average individual dry weight and ash-free dry weight in different sieve fractions of marine meiofauna[J].Marine Biology,1984,84(1):101-108. [16] 张志南,周红,于子山,等.胶州湾小型底栖生物的丰度和生物量[J].海洋与湖沼,2001,32(2):139-147.[17] PALMER M A,COULL B C.The prediction of development rate and the effect of temperature for the meiobenthic copepod,Microarthridion littorale (Poppe)[J].Journal of Experimental Marine Biology and Ecology,1980,48(1):73-83.[18] 吴辰.湛江高桥潮间带小型底栖动物群落的生境多样性研究[D].厦门:厦门大学,2013:34-38.[19] 卓异.泉州湾潮间带不同生境小型底栖动物群落的多样性研究[D].厦门:厦门大学,2014:22-26.[20] 曹婧.福建漳江口红树林和盐沼湿地小型底栖动物的研究[D].厦门:厦门大学,2012:35-36.[21] 常瑜.福建省红树林湿地海洋线虫多样性及分类学的研究[D].厦门:集美大学,2014:11-16.[22] INGOLE B S,PARULEKAR A H.Role of salinity in structuring the intertidal meiofauna ofa tropical estearine beach:field evidence[J].Indian Journal of Marine Sciences,1998,27:356-361.[23] BARBARA K A,BUSS E A.Augmentative applications of Steinernema scapterisci (Nematoda:Steinernematidae) for mole cricket (Orthoptera:Gryllotalpidae) control on golf courses[J].Florida Entomologist,2006,89(2):257-262.[24] 刘均玲,黄勃.红树林生态系统小型底栖动物研究进展[J].海洋科学,2012,36(10):118-122.[25] GOBARDHAN S,SANDALIN R S,ANSARI Z A.Meiofauna-mangrove interaction:a pilot study from a tropical mangrove habitat[J].Cahiers de Biologie Marine,2013,54(3):349-358.[26] 张玉红.台湾海峡及邻近海域小型底栖动物密度和生物量研究[D].厦门:厦门大学,2009:54-56.[27] LAFSSON E.Meiobenthos in mangrove areas in eastern Africa with emphasis on assemblage structure of free-living marine nematodes[J].Hydrobiologia,1995,312(1):47-57.[28] ARMENTEROS M I,MARTIN J P,WILLIAMS B,et al.Spatial and temal variations of meiofaunal communities from the western sector of the gulf ofBatabanó,Cuba.Ⅰ.Mangrove systems[J].Estuaries Coasts,2006,29:124-132.[29] SABINE D.Abundance and distribution of small infauna in mangroves of Missionary Bay,north Queensland,Australia[J].Revista de Biologia Tropical,2001,49(2):535-544. [30] BALASUBRAMANAIAN T,BANDANA D,AYYAPPAN S,et al.Benthic meiofaunal composition and community structure in the Sethukuda mangrove area and adjacent open sea,east coast of India[J].Ocean Science Journal,2011,46 (2):63-72.[31] 周细平.同安湾人工种植红树林对底栖动物生态效应研究[D].厦门:厦门大学,2007:67-70.[32] ALONGI D M.Inter-estuary variation and intertidal zonation of free-living nematode communities in tropical mangrove systems[J].Marine Ecology ProgressSeries,1987,40:103-114.[33] 刘均玲,黄勃,梁志伟.东寨港红树林小型底栖动物的密度和生物量研究[J].海洋学报(中文版),2013,35(2):187-192.[34] MOKIEVSKY V O,TCHESUNOV A V,UDALOV A A,et al.Quantitative distribution of meiobenthos and the structure of the free-living nematode community of the mangrove intertidal zone in Nha Trang Bay (Vietnam) in the South China Sea[J].Russian Journal of Marine Biology,2011,37(4):272-283.[35] COULL B C.Role of meiofauna in estuarine soft-bottom habitats[J].Australian Journal of Ecology,1999,24(4):327-343.。
厦门城市湿地资源的生态敏感性评价及保护对策
厦门城市湿地资源的生态敏感性评价及保护对策作者:林文辉来源:《海峡科学》2007年第06期【摘要】通过对厦门城市湿地资源退化原因分析,采用专家咨询法对指标体系进行咨询,建立湿地敏感程度指标体系,并且构建了湿地生态敏感性评价指数ESA,获得厦门湿地敏感性评估值在31-77之间。
在ArcGis支持下,模拟厦门湿地敏感性的空间分布,并按照敏感性分布情况提出相应保护对策措施。
【关键词】湿地资源生态敏感性评价空间分布对策1引言城市湿地是城市重要的生态基础,是城市可持续发展所依赖的重要的自然生态系统,是城市及其居民能持续获取自然服务的基础。
厦门湿地类型多样,分布广泛。
近年来,经济建设的快速发展及城市化和工业化的同时也致使厦门海岸湿地过度开发,污染加剧,湿地生境受到严重破坏。
为加强湿地的合理开发利用与管理,确保社会、经济、环境的可持续发展,本文从分析厦门海岸湿地面临的资源环境压力分析入手,探讨湿地的“压力-状态-响应”之间的互动关系,利用一定的数学模型,对湿地敏感性程度进行定性与定量相结合综合评估,并借助GIS空间分析模块将湿地退化产生的效应在空间上表达,提出制定厦门湿地资源的保护规划和湿地功能区保护对策。
2厦门湿地退化的主要环境压力分析在城市化过程中湿地景观格局发生了变化,究其原因,既有自然因素,也有人为干扰,实质上是人—地系统动力学作用的结果。
厦门湿地面临的压力因素主要有如下几个方面。
2.1 围海造地围海造地是湿地面积减少的主要原因之一。
厦门市海湾型城市建设的土地资源十分有限,城市发展用地需要过大。
填海造地,填湾架桥等工程不断的占用大面积的海岸、滩涂、浅海水域等湿地。
围海造地减缓了厦门市用地矛盾,促进了种植业、盐业、养殖业的多种经营,但是也致使湿地尤其是浅海水域和滩涂面积的减少。
2.2 海岸工程海岸工程包括港口码头建设、海堤修筑、跨海通道工程等。
海岸工程建设不断,导致海域纳潮面积大大缩小,水动力条件改变。
厦门凤林红树林湿地自由生活海洋线虫群落的研究
MD S标序 分析结果 推 断 自由生 活海洋线 虫的群 落结构 与不 同 的红树植 物形成 的沉积物 有关 。 关键 词 : 红树 林 ; 型底栖 动物 ; 小 自由生活海 洋线虫 ; 落结构 ; 食类 型 群 摄
中图 分 类号 : 7 . Q1 8 1 文献 标 识 码 : A 文童 编 号 :2 34 9 ( 0 8 0 — 1 70 0 5—1 3 20 )40 4—7
段 , 不 同 红 树 种 类 种 植 区 沉 积 物 中 自 由 生 活 海 洋 对
群落 , 是处 于陆地生 态 系统 与海 洋 生 态 系统过 渡 带
的 一 类 特 殊 湿 地 生 态 系 统 , 热 带 亚 热 带 海 岸 带 的 是
线虫 的群 落结构 、 生物多样性 变化 , 势种类 以及 摄 优
性有 过报道 。旧 区( 为 D 断面 ) 树 树 种 为 纯 白 称 红
骨 壤 ( i n i rn ) 植 株 呈 低 矮 丛 状 ,9 9 Avc nama ia , e 18 —
带和亚 热带红树林 群 落 中小 型底 栖 动物 的研 究 , 尤 其 是对 自由生 活海 洋线 虫 的研 究 在不 断 深入 。 国内 , 加强红树植 被恢复 的同时 , 栖动物 多样性 在 底
郭玉 清
(.集美 大 学 水 产学 院 , 1 福建 厦 门 3 12 ;. 美 大学 福 建 省 高校 水 产 科 学 技术 与食 品 安 全重 点 实 验 室 , 建 厦 6 0 12 集 福
门 3 12 ) 6 0 1
摘 要 : 0 4年 冬季在 集美凤林 类似底 质类型 、 20 盐度 和潮位 的海湾地 段 4个断面 , 就不 同红树 植物林 中的小型底栖 动物数量和 自由生活海洋 线虫群 落进 行 了研 究。结 果表 明: 海洋 线虫 是凤林 红 树林 中小 型底 栖动物 中的绝对优 势类群 , 占到小 型底栖 动物 的 7 . %~9 . ; 丰度 来看 , 61 63 从 旧区 白骨
基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建——以厦门市为例
第44卷第6期2024年3月生态学报ACTAECOLOGICASINICAVol.44,No.6Mar.,2024基金项目:国家重点研发计划项目(2022YFF1301300)收稿日期:2023⁃04⁃14;㊀㊀网络出版日期:2023⁃12⁃22∗通讯作者Correspondingauthor.E⁃mail:lntang@iue.ac.cnDOI:10.20103/j.stxb.202304140767李倩瑜,唐立娜,邱全毅,李寿跳,徐烨.基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建 以厦门市为例.生态学报,2024,44(6):2284⁃2294.LiQY,TangLN,QiuQY,LiST,XuY.ConstructionofurbanecologicalsecuritypatternbasedonMSPAandMCRModel:acasestudyofXiamen.ActaEcologicaSinica,2024,44(6):2284⁃2294.基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建以厦门市为例李倩瑜1,2,3,唐立娜1,∗,邱全毅1,李寿跳1,2,3,徐㊀烨1,21中国科学院城市环境研究所城市环境与健康重点实验室,厦门㊀3610212中国科学院大学,北京㊀1000493福建农林大学,福州㊀350002摘要:城市化进程的快速发展加剧了生态系统的退化㊂如何扭转生态系统的退化,同时满足人类日益增长的生态系统服务需求,成为当前的一个研究热点㊂生态安全格局的构建在一定程度上可平衡城市发展与生态环境保护之间的关系,对于保障区域生态安全㊁提升生态系统功能具有重大意义㊂以厦门市为例,基于 生态源地识别 阻力面构建 生态廊道提取 的基本框架构建陆域生态安全格局㊂结合生态系统服务重要性评价和形态学空间格局分析识别生态源地,该方法兼顾了生态结构和功能,使得所识别的生态源地更具全面性㊂选取土地利用类型㊁高程和坡度构建生态综合阻力面,并用人类居住合成指数修正生态综合阻力面,以减少主观赋值的影响,识别各土地利用类型内部的差异,使生态阻力面的构建更加合理㊂在此基础上通过最小累积阻力模型提取生态廊道,利用重力模型量化潜在生态廊道的相对重要性,并根据重力模型结果划分重要性等级㊂研究结果表明,厦门市的生态安全格局由14个生态源地㊁21条生态廊道㊁15个生态节点及若干个踏脚石所组成㊂生态源地主要集中在研究区的西部和北部,以林地和草地为主,面积合计为558.64km2㊂生态廊道长约159.40km,其中,关键生态廊道9条,一般生态廊道12条㊂生态廊道呈现出东西方向联系较为密切,南北方向联系不足的特点㊂根据对区域生态安全的贡献度,将生态安全格局划分为3个管控区进行分级管控㊂将研究结果与厦门市当前的实施计划进行对比分析,虽然结果有所差别,但总体上相对一致,造成差异的主要原因在于两者所采用的研究数据及方法不同㊂因此,研究认为将生态系统服务重要性评价和形态学空间格局分析㊁最小累积阻力模型和重力模型结合,可为生态安全格局的构建提供科学依据㊂关键词:生态安全格局;生态源地;生态阻力面;生态廊道ConstructionofurbanecologicalsecuritypatternbasedonMSPAandMCRModel:acasestudyofXiamenLIQianyu1,2,3,TANGLina1,∗,QIUQuanyi1,LIShoutiao1,2,3,XUYe1,21KeyLaboratoryofUrbanEnvironmentandHealth,InstituteofUrbanEnvironment,ChineseAcademyofSciences,Xiamen361021,China2UniversityofChineseAcademyofSciences,Beijing100049,China3FujianAgricultureandForestryUniversity,Fuzhou350002,ChinaAbstract:Therapiddevelopmentofurbanizationaggravatesthedegradationofecosystem.Howtoreversethedegradationofecosystemandmeettheincreasingdemandforecosystemserviceshasbecomeahotresearchtopic.Tosomeextent,theconstructionofecologicalsecuritypatterncanbalancetherelationshipbetweenurbandevelopmentandeco⁃environmentalprotection,whichisofgreatsignificanceforensuringregionalecologicalsecurityandpromotingecosystemfunction.Inthisstudy,weusedXiamenasanexample,andtheframeworkof ecologicalsourceidentification⁃constructionofecologicalresistancesurface⁃extractionofecologicalcorridors wasadoptedtoconstructtheecologicalsecuritypatternoflandarea.Theassessmentoftheecosystemservicesimportanceandmorphologicalspatialpatternanalysiswerecombinedtoidentifytheecologicalsource.Thismethodtookbothecologicalstructuresandfunctionsintoaccount,makingtheidentifiedecologicalsourcemorecomprehensive.Theecologicalcomprehensiveresistancesurfacewasconstructedbylandusetype,elevationandslope,andcorrectedbyhumansettlementindextoreducetheinfluenceofsubjectiveassignments,identifythedifferenceswithineachlandusetype,andmaketheconstructionofecologicalresistancesurfacemorereasonable.Onthisbasis,theminimumcumulativeresistancemodelwasusedtoextractecologicalcorridors,andthegravitymodelwasusedtoquantifytherelativeimportanceofpotentialecologicalcorridors,andthentoclassifytheimportanceofecologicalcorridors.TheresultsshowedthattheecologicalsafetypatternofXiamencityconsistedof14ecologicalsources,21ecologicalcorridors,15ecologicalnodes,andseveralsteppingstones.Thedistributionofecologicalsourcesmainlyconcentratedinthewestandnorthofthestudyarea,dominatedbywoodlandandgrassland,andthetotalareawas558.64km2.Thelengthoftheecologicalcorridorwereabout159.40km,ofwhich9werekeyecologicalcorridorsand12weregeneralecologicalcorridors,whichhadthecharacteristicsofcloseconnectionbetweeneastandwestandinsufficientconnectioninnorth⁃southdirection.Accordingtothecontributiontotheregionalecologicalsecurity,theecologicalsecuritypatternwasdividedintothreecontrolzonesforhierarchicalcontrol.ComparingthefindingswiththecurrentimplementationplanofXiamen,althoughtheresultsweredifferent,theywererelativelyconsistentoverall,andthemainreasonforthedifferencewasthedifferentresearchdataandmethodsusedinthetwostudies.Therefore,thisstudyconcludedthatthecombinationofecosystemserviceimportanceassessmentwithmorphologicalspatialpatternanalysis,minimumcumulativeresistancemodel,andgravitymodelcouldprovideascientificbasisfortheconstructionofecologicalsecuritypattern.KeyWords:ecologicalsecuritypattern;ecologicalsources;ecologicalresistancesurface;ecologicalcorridor随着城市化的快速发展,高强度的人类活动以及不合理的土地利用使得生态系统日益遭受损坏,生态系统退化越来越呈现出大面积㊁成片蔓延的特点[1],既造成了生态系统质量下降㊁水土流失等一系列严峻的生态环境问题,也威胁了生态安全和人类的可持续发展[2]㊂生态安全格局是以一个相对完整的生态系统作为研究区域,针对区域内的生态环境问题,通过识别并保护潜在的生态关键要素,实现生态环境问题的有效控制和持续改善[3],被视为保障区域生态安全和实现可持续发展的重要途径[4 5]㊂当前,生态安全格局构建已成为研究热点㊂生态安全格局的构建方法具有多元化,最为常见的是由俞孔坚提出的 生态源地识别 阻力面构建 廊道提取 的生态安全格局构建方式㊂然而,在生态源地识别和阻力面构建方面仍存在一定的局限性㊂具体表现为:(1)现有研究在生态源地的选取中侧重考虑生境斑块的内在功能属性,对斑块在景观中的空间结构较少关注,忽视了其与周围环境之间的联系[6],个别研究则直接将自然保护区等特定的生态功能区视为生态源地,该方法带有一定的政策性和主观性,缺乏定量分析㊂(2)构建生态阻力面选取的阻力因子大多带有较强的主观性,且同一土地利用类型无差别的赋值方式疏忽了其内部自然属性的差异,也无法体现同一土地利用类型下人类活动有差别的干扰[4]㊂生态源地提取㊁生态阻力面构建作为生态廊道构建的两个重要前提,影响了生态廊道的数量和走向,进而影响了生态安全格局的构建㊂因此,本研究拟针对上述两个问题,进一步优化和完善生态源地识别和生态阻力面构建,从而使生态安全格局的构建更具有合理性㊂厦门市作为经济特区,历经多年高强度的开发建设,围海造地与城市新区发展㊁旧区改造并行,建成区由岛内向岛外逐渐蔓延扩张,导致大量耕地㊁林地㊁湿地㊁滩涂等生态空间转变为城市用地㊂近年来由于气候变化和城市化发展影响叠加,部分区域生态功能退化,生态安全受到威胁㊂为实现高质量发展,厦门市已开展了生态修复项目㊂本研究拟优化和完善生态安全格局构建方法,为厦门市的生态修复工作提供科学依据㊂5822㊀6期㊀㊀㊀李倩瑜㊀等:基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建㊀6822㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀1㊀研究区概况与数据来源1.1㊀研究区概况㊀㊀厦门市(24.23ʎ 24.25ʎN㊁117.53ʎ 118.26ʎE)位于福建省东南沿海,由思明㊁湖里㊁集美㊁海沧㊁同安和翔安6个行政区组成,陆地总面积1700.61km2(图1)㊂北邻泉州,南接漳州,东临台湾海峡,是闽南 金三角 的中心, 一带一路 的海陆交通枢纽㊂属南亚热带海洋性季风气候,年均气温21ħ左右,年均降雨量为1200mm左右,常年气候温暖,雨热同期,雨量充沛㊂地势西北高㊁东南低,以滨海平原㊁山地和丘陵为主㊂图1㊀厦门市区位图Fig.1㊀LocationoftheXiamen1.2㊀数据来源本研究使用的数据包括植被净初级生产力㊁数字高程模型㊁月平均气温㊁月平均降水量㊁土壤质地㊁土地利用数据㊁夜间灯光数据以及归一化植被指数,数据来源详见表1㊂为减小数据误差,保障空间参考的一致性,将坐标统一为GCS_WGS_1984,并将栅格数据重采样为30m的分辨率㊂表1㊀数据来源Table1㊀Datasource数据类型Datatypes数据来源DataaccuracyDatasources数据精度植被净初级生产力NPP资源环境科学与数据中心2001 2020年500m数字高程模型DEM地理空间数据云30m月平均气温Monthlymeantemperature国家地球系统科学数据中心2001 2020年1km月平均降水Monthlymeanprecipitation国家地球系统科学数据中心2001 2020年1km土壤质地Soiltexture世界土壤数据库(HWSD)的中国土壤数据集(v1.1)1km土地利用LanduseGlobeLand3030m夜间灯光Nightlight科罗拉多矿业大学https://eogdata.mines.edu/products/vnl/2020年500m归一化植被指数NDVI地理空间数据云2020年30m㊀㊀NPP:植被净初级生产力Netprimaryproductivity;DEM:数字高程模型Digitalelevationmodel;NDVI:归一化植被指数Normalizeddifferencevegetationindex2㊀研究方法2.1㊀生态源地识别作为生态安全格局构建的第一环节,生态源地能否准确识别至关重要,直接影响了生态廊道构建和生态修复空间识别的结果[7]㊂本研究拟从生态系统结构和功能两个方面识别生态源地,提高生态源地识别的准确性㊂2.1.1㊀生态系统服务重要性评价生态系统服务是指人类从生态系统中所获得的效益,包括人类赖以生存的自然环境条件与效用[8]㊂生态系统服务能力反映了生态环境的状况,根据研究区特点,选择水源涵养㊁水土保持以及生物多样性保护这3个生态系统服务作为厦门市生态系统服务重要性的评价指标,并根据‘生态保护红线划定技术指南“(2015)提供的NPP定量指标法进行评价㊂为避免主观赋值导致研究结果有所偏倚,默认3种服务同等重要,即权重一致[9]㊂运用分位数法(Quantiles)进行分级再等权叠加,得到研究区域的生态系统综合服务能力指数,并将其划分为5个等级㊂2.1.2㊀MSPA分析形态学空间格局分析(MorphologicalSpatialPatternAnalysis,MSPA)是2007年由Vogt等提出的一种基于数学形态学原理对二值化的栅格图像进行分类的方法[10 11]㊂该方法简单高效,可快速地识别景观类型,且不受研究尺度的限制㊂本研究选取受人类干扰较大的耕地㊁建设用地作为背景,选取林地㊁草地㊁灌木地㊁湿地㊁水体等自然生态要素作为前景㊂基于GuidosToolbox软件,采用默认的八领域分析法进行计算,得到厦门市的7类景观类型,即核心区(core)㊁孤岛(islet)㊁边缘区(edge)㊁孔隙(perforation)㊁桥接区(bridge)㊁环道区(loop)以及支线(branch),进而提取生境斑块最大的核心区作为潜在的生态源地[6]㊂2.1.3㊀生态源地提取将生态系统服务中度及以上重要性区域与潜在的生态源地进行叠加分析,提取面积大于1km2的重叠的核心区斑块作为生态源地㊂进而基于Conefor和ConeforInputsforArcGIS10.x插件对其进行景观连通性计算㊂根据输出结果,斑块重要性指数(thedeltavaluesforprobabilityofconnectivity,dPC)的值越大,说明该斑块对维持较高的景观连通性发挥的作用越大㊂2.2㊀生态阻力面构建物种在生态源地之间的迁移和扩散在一定程度上会受到土地覆被状态和人类活动的阻扰[12]㊂作为生态廊道能否准确识别的关键,构建生态阻力面模拟生态要素流动和传递的难易程度,对于生态安全格局的构建至关重要㊂本研究选取土地利用类型㊁高程和坡度这3个影响较大且较常使用的自然因子构建生态综合阻力面,参考相关文献进行分级和赋值[13 14](表2)㊂为弱化主观赋值的影响,以人类居住合成指数(HumanSettlementIndex,HSI)表征人类活动对生态要素流动和传递的干扰,对生态综合阻力面进行修正[15 16](公式1 3)㊂NTLnor=NTL-NTLminNTLmax-NTLmin㊀㊀㊀㊀㊀㊀(1)HSI=1-NDVImax()+NTLnor1-NTLnor()+NDVImaxˑNTLnorˑNDVImax(2)式中,NTL㊁NTLmax㊁NTLmin分别为原始的夜间灯光数据及其最大值和最小值;NTLnor为归一化的夜间灯光数据;NDVImax为归一化植被指数的最大值;HSI为人类居住合成指数㊂Ri=HSIiHSIaˑR(3)式中,Ri为基于人类居住合成指数修正的生态阻力系数;HSIi为栅格i的人类居住合成指数;HSIa为栅格i对7822㊀6期㊀㊀㊀李倩瑜㊀等:基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建㊀应的景观类型a的平均人类居住合成指数;R为栅格i的景观类型的综合生态阻力系数㊂表2㊀生态阻力系数2.3㊀生态廊道提取2.3.1㊀MCR模型生态廊道作为生态要素在生态源地之间流动和传递的重要途径[17],具有维持生态系统运转㊁维护区域生态安全的重要功能[18],也是生态修复中最有可能改善和提高连通性的关键区域[19]㊂生态廊道构建的方法包括最小累积阻力模型(MinimumCumulativeResistance,MCR)[20 21]㊁蚁群算法[22]㊁电路理论[23 24]㊁小波变换[25]等㊂其中,MCR模型是俞孔坚在Knaapen等提出的模型的基础上改进得到的[5]㊂相较于其他模型,该模型可更好地模拟和量化物种㊁能量和信息在生态源地之间流动的最小成本路径,已被广为采用[26 27]㊂因此,本研究采用MCR模型,基于ArcGIS提取每一个生态源地的中心点为生态源点,以修正后的生态阻力面为成本,通过成本路径工具,模拟每一个生态源点到其他n-1个生态源点的最小成本路径,构建两两之间的潜在生态廊道,共计C2n条㊂计算方法[17]如公式4:MCR=fminði=mj=nDijˑRi()(4)式中,MCR为物种从生态源地扩散到其他生态源地的最小累积阻力值;Dij为物种从生态源地j到景观单元i的空间距离;Ri为景观单元i对应的生态阻力系数,即前文所述的基于人类居住合成指数修正后的生态阻力系数㊂2.3.2㊀重力模型基于重力模型构建生态源地之间的相互作用矩阵,以此量化潜在生态廊道的相对重要性㊂相互作用力越大说明生态源地之间的联系越紧密,生态要素流动和传递越频繁,生态源地之间的生态廊道重要性等级越高,计算方法[28]如公式5:Gij=L2maxlnSi()lnSj()L2ijPiPj(5)式中,Gij为斑块i和斑块j之间的相互作用力;Pi和Pj分别为斑块i和斑块j的阻力值;Si和Sj分别为斑块i和斑块j的面积;Lij为斑块i和斑块j之间潜在生态廊道的累积阻力值;Lmax为研究区所有潜在生态廊道的最大累积阻力值㊂3㊀结果与分析3.1㊀生态源地识别3.1.1㊀生态系统服务重要性评价水源涵养㊁水土保持和生物多样性保护重要性等级大体上呈现出西北高东南低的特征(图2)㊂由这三者8822㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀等权叠加得到的生态系统综合服务重要性等级也呈现出相同的空间分布特征㊂中度及以上重要区主要位于研究区的西部和北部,面积合计为753.25km2,约占研究区总面积的47.71%㊂中部和南部耕地和建成区较为密集,受人类活动影响较大,生态系统综合服务重要性等级较低,主要为不重要区和轻度重要区,面积分别为406.84km2和418.68km2㊂图2㊀生态系统服务重要性评价结果空间分布Fig.2㊀Spatialdistributionofimportanceassessmentresultsofecosystemservices3.1.2㊀MSPA分析基于MSPA分析得到7类生态景观的面积和占比(表3)㊂7类生态景观的总面积约为692.60km2㊂核心区的面积最大,约为635.19km2,占生态景观总面积的91.71%㊂边缘区为核心区的外部边界,是核心区与其外部的非生态景观类型之间的过渡区域;孔隙为核心区与其内部存在的非生态景观类型之间的过渡区域㊂两者分别占生态景观总面积的5.82%和1.45%㊂孤岛零星散布在研究区域中,约占生态景观总面积的0.17%㊂支线㊁桥接区和环道区均具有连通作用,数量越少意味着连通性越差,生态要素流动和传递的阻扰越大,越不利于生物多样性[29]㊂其中,支线作为连通核心区与其他生态景观之间的条带状区域,占生态景观总面积的0.65%,说明生态要素在核心区与其他生态景观之间流动和传递受到较大的阻扰,连通性较差;桥接区是连通各核心区之间的条带状区域,约占生态景观总面积的0.15%;环道区为核心区内物种迁徙的捷径,面积最小,仅为0.33km2,占比为0.05%㊂表4㊀景观类型分类统计由图3可知,核心区在西北部连片聚集,整体性较好,而在其他地区则呈零散分布,破碎化较为严重㊂在9822㊀6期㊀㊀㊀李倩瑜㊀等:基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建㊀研究区西北部的核心区,虽然面积大且连片分布,但其间也具有多个孔隙,孔隙的范围越大,表明非生态景观类型占据的面积越多,或者意味着核心区生态系统退化的范围扩大,从而使生态要素在流动和传递的过程中受到一定程度的阻碍㊂孤岛在一定程度上可作为物种迁徙的踏脚石,既减小生境斑块之间的成本距离,又可间接提高生境斑块之间的连通性[29 30]㊂例如,位于同安区南部的核心区,其右侧间隔分布着多个孤岛,呈弧形状处于该核心区与其他两个较小的核心区之间,可使生态要素在流动和传递的过程中能够短暂栖息㊂图3㊀基于形态学空间格局分析的景观类型空间分布Fig.3㊀Spatialdistributionofthelandscapetypesbasedonmorphologicalspatialpatternanalysis3.1.3㊀生态源地提取基于生态系统服务重要性和MSPA的分析结果,共有14个面积大于1km2的核心区斑块与中度及以上生态系统综合服务重要性区域重叠,面积合计为558.64km2,约占核心区总面积的87.95%㊂表明多数核心区拥有较好的生态系统服务能力,可为生物生存提供较好的栖息条件㊂因此,选取这14个重叠的核心区斑块作为生态源地(图4)㊂生态源地主要位于研究区西部㊁北部和东北部地区,涵盖国家级和省级森林公园㊁水源保护区等重要区域㊂中部和南部地区以建设用地和耕地为主,人类活动强度较大,生态源地数量较少且较为分散㊂图4㊀生态源地空间分布Fig.4㊀Spatialdistributionofecologicalsources根据景观连通性分析结果(表5)可知,14个生态源地的斑块重要性(dPC)差距较大,仅有3个生态源地0922㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀的斑块重要性大于1,分别为10号㊁7号和6号生态源地,其余的11个生态源地的斑块重要性均小于1㊂其中,10号生态源地面积最大,约占14个生态源地总面积的82.71%,其斑块重要性也最大,说明该生态源地对于维持较高的景观连通性发挥了重要作用㊂表5㊀景观连通性计算结果3.2㊀生态阻力面构建由土地利用类型㊁高程和坡度这三个自然环境因子所构建的生态综合阻力面(图5),空间分布上呈现出明显的南北异质性㊂东南部人类活动密集的区域阻力系数明显高于西北部,但高阻力系数零星分布在西北部㊂主要原因在于这些地区单因子阻力系数为 两高 或 三高 ,从而使得其综合阻力系数相对于周边地区呈现出较高的趋势㊂经HSI修正后的生态综合阻力面,各土地利用类型内部的阻力系数发生了显著变化㊂高阻力值由西北部转移至中部和南部,且高阻力区域具备一定的规模㊂尤其是湖里区和思明区,因开发建设早城市化水平较高,且受区域面积的限制,人类活动区域高度聚集,对生态要素流动和传递的干扰明显增强㊂图5㊀生态阻力面空间分布Fig.5㊀Spatialdistributionoftheecologicalresistancesurface3.3㊀生态廊道构建通过MCR模型提取研究区潜在的生态廊道㊂剔除重复路径和经过生态源地内部的无效路径,最终得到21条生态廊道,总长度为159.40km㊂其中,生态源地14在地理位置上与其他生态源地存在海域的阻隔,彼此之间未能构筑起生态廊道㊂因本研究的生态阻力系数设置比较大,由此计算得到的各生态源地之间的相互作用力也较大(表6)㊂参考相关文献[6],以10为临界值对生态廊道的重要性进行等级划分,大于10的视为关键生态廊道,小于10的则为一般生态廊道,共得到关键生态廊道9条,长度为83.11km,一般生态廊道1922㊀6期㊀㊀㊀李倩瑜㊀等:基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建㊀12条,长度为76.29km(图6)㊂表6㊀基于重力模型的生态源地相互作用矩阵Table6㊀EcologicalsourcesinteractionmatrixbasedonGravityModel源地Source1234567891011121314166.5019.3143.582.6212.180.843.563.481.161.190.720.60-279.2988.0814.1617.582.274.985.302.841.470.910.77-31365.278.8391.452.1111.6410.462.922.501.471.22-45.063.325.292.053.574.560.850.560.49-541.540.9014.7211.081.511.831.020.82-61.030.731.1736.820.400.280.26-76.0150.251.503.541.701.24-85.291.872.811.381.05-92.060.730.500.45-101.911.130.94-1119.187.34-1231.01-13-图6㊀生态廊道空间分布Fig.6㊀Spatialdistributionofecologicalcorridors结合景观连通性与重力模型分析结果(表5与表6)可知,对景观连通性具有重大贡献的10号和6号生态源地,两者之间距离较远,但相互作用力较大,表明其间的生态廊道是生态要素流动和传递可能性较大的路径,相对重要性较高㊂7号生态源地对景观连通性的贡献程度也较高,与9号生态源地之间的相互作用力最大,其次为8号生态源地,与南北方向的生态源地之间的相互作用力总体偏弱,表明7号生态源地对景观连通性的贡献主要在于连通东西方向的生态源地,特别是与之距离较远的9号生态源地㊂由此本研究认为,生态源地之间距离越近并不意味着彼此之间的联系更紧密,远距离的生态源地之间也存在频繁的生态要素流动和传递㊂2号㊁3号和4号生态源地面积均较小,斑块重要性(dPC)也较差,特别是4号生态源地,其dPC值几乎为0,但三者之间的相互作用力较强,表明对景观连通性贡献度不高的生态源地之间生态要素的流动和传递也具有较大的可能性,其生态源地和生态廊道也具有重要的生态保护和修复意义㊂综合上述分析,本研究认为以往研究根据斑块重要性(dPC)的大小筛选生态源地,可能导致部分具有重要生态功能的斑块被排除在外,从而影响生态廊道的提取㊂因此,本研究保留斑块重要性较差的生态源地,可使生态安全格局更具有完整性㊂3.4㊀生态安全格局构建与分区管控基于上述分析可知,研究区共有生态源地14个,面积约占研究区总面积的35.39%,呈现出西北多东南少的分布特征,基本涵盖了多个国家级或省级森林公园㊁水源保护区等㊂其中,西北部连片的生态源地为研究区筑起了天然的生态保护屏障,有利于维护研究区的生态安全㊂研究区的生态廊道共21条,总长为159.40km㊂关键生态廊道和一般生态廊道纵横交错,将各个生态源地串联起来,是研究区生态安全网络至关重要的组成2922㊀生㊀态㊀学㊀报㊀㊀㊀44卷㊀图7㊀生态安全格局分区管控Fig.7㊀Ecologicalsecuritypatternzoningcontrol部分,保障了水源涵养㊁水体保持㊁生物多样性保护等生态功能的发挥㊂叠加MSPA分析结果发现,在不属于生态源地的核心区中,有15个处于生态廊道与生态源地之间㊁生态廊道与生态廊道之间的交汇处,是生态要素流动和传递的重要节点;有23个处于生态廊道的沿线,可与孤岛共同发挥踏脚石的作用㊂基于对维护区域生态安全的重要性程度,对生态安全格局进行分区管控(图7)㊂将生态源地划为一级管控区;将关键生态廊道及其沿线的一般核心区和孤岛列为二级管控区,关键生态廊道长度为83.11km,一般核心区面积为11.29km2,孤岛面积为0.16km2;将一般生态廊道及其沿线的一般核心区和孤岛列为三级管控区,一般生态廊道长度为76.29km,一般核心区面积为10.98km2,孤岛面积为0.06km2㊂4㊀讨论本研究基于生态源地识别一阻力面构建一生态廊道构建的基本框架,构建了厦门市的陆域生态安全格局㊂‘厦门市国土空间生态修复三年行动计划(2020 2022年)“(以下简称‘计划“)中的生态修复项目分布图也是在生态安全格局构建的基础上进行识别的㊂何子张等对‘计划“所采用的生态安全格局构建方法和结果进行了介绍[31]㊂通过对比可知本研究与‘计划“所识别的生态源地较为一致,但也存在些许不同:(1)‘计划“所识别的生态源地面积明显大于本研究的识别结果,特别是西北部的生态源地,‘计划“的识别结果范围更广,斑块整体性更好㊂主要原因在于‘计划“所采用的基础数据为 三调 数据,数据更加真实可信㊂此外,‘计划“将耕地纳为生态源地,而本研究则将耕地视为背景未进行分析㊂前景分析的目的在于识别出适宜生物栖息和活动的区域,多数学者在进行前景分析时也大多选择生态功能较好且受人类影响较小的自然景观[32 33],耕地受人类活动影响较大,一般不作考虑㊂(2)‘计划“兼顾了陆域和海域,本研究因数据获取有限,且海域与陆域评价方法不同,因此未考虑海域范围㊂(3)‘计划“将风景名胜区㊁自然保护区㊁生态红线区等直接识别为生态源地,而根据本研究的分析,鼓浪屿不属于生态系统服务中度及以上重要区域,因此未列为生态源地㊂在生态阻力面构建过程中,‘计划“所构建的生态阻力面同一土地利用类型的生态阻力系数较为均等㊂本研究为进一步识别同一土地利用类型各生态阻力系数的差异,以HSI修正生态阻力面㊂相比单一使用夜间灯光数据进行修正,HSI综合考虑了夜间灯光数据和NDVI数据,可弥补夜间灯光指数过度饱和的缺陷,更能精细地刻画人类活动干扰强度的空间分布特征,修正效果更好[34 35]㊂因生态源地和生态阻力面有所差异,从而导致本研究和‘计划“基于MCR模型构建的生态廊道也有所不同,但生态廊道的走向大致相同,且本研究所识别的生态廊道既包含了‘计划“基于MCR模型所构建的潜在生态廊道,也包括部分山脊廊道,说明本研究所构建的生态廊道具有一定的合理性㊂本研究虽然构建了生态廊道,但因各生态要素对生态廊道的宽度要求不同,目前尚未形成统一的划定标准㊂因此,本研究未对生态廊道的宽度做进一步分析,今后将针对这一问题进行深入探究㊂厦门市作为一个滨海城市,海域也占据一定的面积,本研究仅考虑陆域生态安全格局,具有一定的局限性,今后将统筹考虑陆域和海域,更加全面地构建全域的生态安全格局㊂5㊀结论本研究采用生态系统服务重要性评价和MSPA分析识别生态源地,兼顾了生态功能和结构,定量分析的3922㊀6期㊀㊀㊀李倩瑜㊀等:基于形态学空间格局分析和最小累积阻力模型的城市生态安全格局构建㊀。
湿地景观变化与生态系统功能评价
湿地景观变化与生态系统功能评价湿地是一种特殊而重要的生态系统,它具有独特的生物多样性和生态功能。
然而,由于人类活动的不断扩张,湿地景观正在发生着巨大的变化。
这种变化对湿地的生态系统功能产生了深远的影响。
本文将探讨湿地景观变化的原因、影响以及生态系统功能的评价。
湿地景观变化的原因可以归结为自然因素和人为因素两个方面。
自然因素包括气候变化、地质活动等,这些因素的变化会直接导致湿地的水位、植被和土壤的改变,进而影响生态系统功能。
人为因素主要包括农业、城市化和工业化等人类活动对湿地的直接影响。
农业的发展导致了湿地的干燥和水资源的污染,城市化和工业化则导致了湿地的填充和破坏。
这些人为因素对湿地景观造成了巨大的变化,进而对湿地的生态系统功能产生了严重的影响。
湿地景观变化对生态系统功能的影响主要体现在以下几个方面。
首先,湿地景观的改变会直接影响湿地的水循环功能。
湿地作为自然的蓄水池,可以调节水位和水质,保持水量平衡和水质稳定。
然而,湿地景观的变化导致了湿地的干燥和水资源的流失,使得水循环功能受到破坏。
其次,湿地景观改变对生物多样性的影响非常大。
湿地是许多珍稀动植物的家园,但由于湿地景观的变化,许多物种失去了适宜的栖息环境,导致生物多样性的丧失。
此外,湿地景观的改变也会影响湿地的碳储存和气候调节功能。
湿地是天然的碳汇,能够吸收大量的二氧化碳,并且具有减缓气候变化的能力。
然而,湿地景观的改变会导致湿地的碳储存减少,进而影响气候调节功能。
为了评价湿地的生态系统功能,可以综合考虑湿地景观的物理结构和生态过程。
物理结构主要包括湿地的土壤类型、水文条件、植被覆盖等,而生态过程则包括湿地的生产力、物质循环和能量流动等。
通过对湿地结构和过程的综合分析,可以评估湿地的水资源调节、生物多样性维护、碳存储和气候调节等功能的状况和潜力。
评价的结果可以为湿地保护与恢复提供科学依据和决策支持。
综上所述,湿地景观变化对生态系统功能产生了深远的影响。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
近10年来厦门市湿地动态变化分析林巧莺,林广发,杨陈照,游丽平(福建师范大学地理科学学院,福州35007)摘要:通过对厦门市1993年TM遥感影像和2003年ETM+遥感影像的解译分析统计,有效地提取了1993年、2003年厦门市各湿地类型的面积统计数据、以及1993年~2003年各湿地类型的转移矩阵,并进行了主要驱动力分析。
结果表明:1993年~2003年近10年来,由于经济发展、人口增长以及农业生产比较利益的驱动和农业内部结构调整的影响,厦门市养殖区面积变化最大,增加了3990.17公顷,主要通过占用非湿地地类、滩涂、盐田和水库水塘来增加的;红树林的大幅度减少,则主要是由于非湿地地类、养殖区的侵占、以及遭受破坏变为滩涂;养殖区、水库水塘、滩涂、盐田等许多湿地类型也大量被非湿地地类侵占。
关键词:湿地;转移矩阵;厦门市1 引言湿地作为“地球之肾”,是由水陆相互作用而形成的自然综合体,是自然界最富生物多样性的生态景观和人类最重要的生存环境之一[1][2],具有保持水源、蓄洪防旱、调节气候、保护海岸和保持生物多样性等功能。
对于拥有丰富湿地资源的沿海花园式城市的厦门市来说,保护和合理利用湿地资源不仅是厦门市生态环境保护的需要,也是厦门市可持续发展、建设海湾型园林城市的需要。
因此,及时准确地掌握厦门市湿地的时空动态变化,对于厦门市湿地的合理开发和保护具有重要的意义。
目前,土地利用/土地覆盖变化(LUCC)研究是全球环境变化研究的重要组成部分,对于区域土地利用/覆盖的变化及其驱动因子的研究也是目前研究的热点之一[3]-[12]。
本研究综合运用“3S”技术对厦门市1993年的TM影像和2003年的ETM+影像进行分析,从而得到近10年来厦门市湿地的动态变化信息,并对此进行主要驱动力分析。
2 研究区与研究方法2.1 研究区概况厦门市位于117º53′~118º25′E,24º23′~24º55′N之间,地处我国东南沿海——福建省东南部,濒临台湾海峡,属南亚热带季风型海洋性气候,是一座环境优良的滨海城市,享有全国首批“国家环境保护模范城市”与“国际花园城市”等盛誉,也是中国首批沿海开放城市之一。
全市辖思明、湖里、集美、海沧、同安和翔安6个区,土地面积1569.3km2(不含滩涂面积),海域面积约300多km2(图1)。
作者简介:林巧莺(1980—),女,福建福州市人,在读硕士研究生,研究方向:GIS与遥感应用E-mail:mylqy_2000@图1 研究区区域及其位置2.2 技术路线本次研究是以厦门市1993年TM遥感影像和2003年ETM+遥感影像为基本资料,以目视解译为基础,利用GIS软件进行统计分析、空间分析,得到1993年和2003年厦门市各湿地类型面积统计数据和1993年~2003年湿地类型转移矩阵表。
技术流程图如图2:图2 厦门市湿地动态变化研究技术流程图2.3研究方法在统计各湿地类型的转移矩阵(包括湿地与非湿地类型间的转移)时,是利用公式1进行赋值,然后分类统计分析得到转移矩阵数据的。
公式1:P=P1*10n+P2 (属性字段的类型为数值型方可利用该公式进行运算赋值)其中,P1 、P2分别为第1、2期的厦门市土地利用类型的属性字段,P为1、2两期湿地类型变化的属性字段。
n为P2字段的最大位数。
在本研究中,由于湿地类型和非湿地类型共十类,所以n取2。
利用公式1进行赋值后,P字段的数值就为三位数或四位数,如608表示1993年养殖区(属性字段值为6)到2003年时转变为了盐田(属性字段值为8)。
3 厦门市湿地的动态变化3.1 湿地分类体系的建立根据科学性、系统性和实用性的原则,参照《全国湿地资源调查与监测技术规程》的有关要求及福建省湿地类型划分标准,结合厦门市滨海城市的特点,重点突出厦门市海岸附近湿地类型,拟订分类体系。
具体分为:浅海水域(以6m等深线为外边界)、沙滩、河口水域、滩涂、红树林、养殖、水库水塘、盐田、河流九大类。
表1 湿地类型及其定义编码 湿地类型 定义1 浅海水域 低潮时水深不超过6米的永久水域,植被盖度<30%,包括海湾、海峡(根据《全国湿地资源调查与监测技术规程》)2 沙滩 潮间植被盖度<30%,底质以砂、砾石为主(福建省湿地类型划分标准――潮间砂石海滩)3 河口水域 从潮差为零界线至淡水舌锋缘之间的永久性水域4 滩涂 淤泥质潮间带,还包括潮上带和潮下带可供开发利用的部分。
5 红树林 以红树植物群落为主的潮间沼泽(福建省湿地类型划分标准)6 养殖区 指近海区域的水产养殖与滩涂养殖7 水塘、水库 主要包括池塘、湖泊和水库8 盐田 借助海水蒸发来制盐的沿海湿地9 河流 主要包括永久性河流和季节性或间歇性河流3.2 各湿地类型的面积统计及其转移矩阵用ArcGIS分别对1993年和2003年影象的解译结果进行拓扑检查,并统计各湿地类型的面积,得到1993年~2003年厦门市各湿地类型面积及其增减情况(表2)。
在统计1993~2003年各湿地类型的转移矩阵(包括湿地与非湿地类型间的转移)时,首先将1993年和2003年影像解译结果分别与厦门市行政区划图(包括海域部分)进行叠置分析,利用公式1进行分类统计分析后,得到1993年~2003年厦门市各湿地类型转移矩阵数据(表3)。
表2 1993年—2003年厦门市各湿地类型面积及其增减情况(单位:公顷)浅海水域沙滩 河口水域 滩涂 红树林养殖区水 库水 塘 盐田 河流 总和1993年 17266.02 69.84 265.28 8947.43 50.33 5778.12 3552.42 1674.75 709.05 38313.24 2003年 15846.41 48.03 284.18 8796.22 17.02 9768.30 3297.80 713.48 630.16 39401.60 1993-2003年的增减 -1419.61-21.81 18.90 -151.22-33.313990.17 -254.62-961.27 -78.891088.36占1993年的百分比(%) -8.22-31.23 7.12 -1.69-66.1869.06 -7.17-57.40 -11.132.84表3 1993—2003年厦门市湿地类型转移矩阵表(单位:公顷)2003年1993年浅海水域 河流 沙滩 河 口水 域 滩涂红树林 养殖区水 库水 塘 盐田非湿地地类浅海水域 15503.90 0.00 3.09 0.03 1577.59 0.00 50.02 0.08 0.00 131.31沙滩 8.35 0.00 36.51 0.00 0.00 0.00 0.00 0.00 0.00 24.99 河口水域 0.57 0.07 0.00 199.00 27.60 0.00 33.30 0.00 0.62 4.11滩涂 213.12 1.17 0.20 60.14 6983.90 8.70 1099.14 48.72 17.03 515.31 红树林 1.34 0.00 0.00 0.00 10.57 6.77 15.47 0.00 0.00 16.18 养殖区 0.01 11.39 0.00 15.61 102.77 1.26 4567.85 48.13 12.92 1018.19 水库水塘 0.00 5.97 0.00 0.00 4.88 0.00 354.84 2180.59 0.00 1006.13 盐田 0.00 0.09 0.00 0.81 11.29 0.00 910.61 7.35 639.08 105.51河流 0.00 408.01 0.00 1.41 0.00 0.00 24.63 2.13 0.00 272.87 非湿地地类 119.13 203.45 8.23 7.18 77.62 0.28 2712.43 1010.79 43.82 148919.213.3 湿地动态变化结果分析从表2可看出,1993年~2003年间,厦门市养殖区面积变化最大,增加了3990.17公顷,其次为浅海水域,减少了1419.61公顷,盐田、水库水塘、滩涂则分别减少了961.27公顷、254.62公顷、151.22公顷。
其中,变化幅度最大的也是养殖区,其增加的面积占了1993年养殖区面积的69.06%,其次是红树林快速消失,1993年的红树林有66.18%已经遭砍伐,盐田和沙滩的减少幅度也较大,分别占1993年的57.40%、31.23%。
从表3湿地类型转移矩阵表可知,1993年~2003年间,厦门市养殖区主要通过占用非湿地地类、滩涂、盐田和水库水塘来增加的,分别占用了2712.43公顷、1099.14公顷、910.61公顷、354.84公顷。
红树林的大幅度减少,则主要是由于非湿地地类、养殖区的侵占、以及遭受破坏变为滩涂,红树林转换为非湿地地类、养殖区、滩涂的面积分别为16.18公顷、15.47公顷、10.57公顷。
盐田的减少,主要是因为大部分盐田被改造成了养殖区,盐田转换为养殖区的面积为910.61公顷,超过了1993年~2003年间未发生变化的盐田的面积(639.08公顷),一部分盐田也被非湿地地类侵占,盐田转换为非湿地地类的面积为105.51公顷。
水库水塘的减少,主要是由于非湿地地类的侵占,一部分也被改造成养殖区,水库水塘转换为非湿地地类、养殖区的面积分别为1006.13公顷、354.84公顷。
滩涂的减少则主要是由于养殖区、非湿地地类的围垦而造成的,滩涂转换为养殖区、非湿地地类的面积分别为1099.14公顷、515.13公顷。
4 主要驱动力分析4.1 经济发展和人口增长1993年~2003年,厦门市经济发展迅猛,国内生产总值从1993年的1323163万元发展为2003年的7596934万元,增长了将近5倍,其中,第二产业的生产总值从1993年的590942万元增长到2003年的4445117万元,增长了6.5倍多。
人均GDP从1993年的10945元增加到2003年的53586元。
在这期间,厦门市人口增长也很快,1993年年末总人口为117.49万人,到了2003年,年末总人口增长到141.76万人,增长了20.65%[13][14]。
经济的发展、人口的增长,必然导致了如建设用地等非湿地地类的迅猛发展。
1993年~2003年期间,就有1018.19公顷的养殖区、1006.13公顷的水库水塘、515.31公顷的滩涂被非湿地地类侵占。
红树林遭受人为的破坏也十分严重,绝大部分的红树林被转换为非湿地地类、养殖区、滩涂。
4.2 农业生产比较利益的驱动和农业内部结构调整由于养殖业与其他农业生产活动的经济比较利益的存在以及农业内部结构的大调整,目前水产业已成为厦门市农业经济的五大支柱产业之一,并且已初步形成由水产养殖、海洋捕捞、水产品加工、流通服务组成的产业体系,成为厦门市农业生产的重要产业。