第九章 突变、修复与重组
高中生物DNA修复与突变知识点总结

高中生物DNA修复与突变知识点总结DNA修复与突变是生物学中重要的概念,对于高中生物学知识的理解至关重要。
本文将通过总结DNA修复与突变的知识点,帮助读者更好地理解此概念。
一、DNA修复的概念与意义DNA修复是一种生物体对DNA中产生的损伤进行修复的过程。
DNA损伤的原因包括外源性因素(如辐射、化学物质)和内源性因素(如代谢产物)。
DNA修复的意义在于保持遗传物质的完整性和稳定性,确保生物正常生长、发育和繁殖。
二、DNA损伤与修复的分类1. 物理性损伤:来自于辐射,如紫外线。
2. 化学性损伤:如环境污染物、化学试剂等。
3. 内源性损伤:细胞内代谢过程中产生的氧自由基等。
4. DNA修复机制:分为直接修复、错配修复、切除修复和重组修复等。
三、DNA损伤与修复的主要类型及机制1. 基因突变:DNA碱基发生改变,包括点突变、插入突变和缺失突变等。
DNA聚合酶的错误复制和DNA损伤引起的突变是最常见的突变机制。
2. 染色体畸变:染色体结构的改变,包括染色体缺失、染色体重排等。
染色体的非同源重组和染色体断裂产生的重排引起染色体畸变。
3. 光反应和化学损伤:紫外线、化学物质等引起DNA光反应和化学反应,导致碱基损伤、碱基交联和DNA链断裂等。
4. 不同的修复机制:直接修复修复DNA中的碱基损伤,错配修复修复DNA复制过程中出现的碱基配对错误,切除修复修复DNA链断裂,重组修复修复DNA双链断裂。
四、重要的DNA修复机制1. 直接损伤修复:包括光修复和酶修复。
光修复是利用光酶修复DNA中的嘌呤二聚体损伤,酶修复则是利用特定的酶修复DNA中的碱基损伤。
2. 错配修复:包括错配识别、错配切除和错配修复等步骤,通过DNA聚合酶和联合酶的作用修复DNA中的错配碱基。
3. 切除修复:包括碱基切除修复、核苷酸切除修复和修复性DNA 重组等不同类型。
切除修复是通过核酸内切酶、外切酶等酶的作用将损坏部分切除,然后通过DNA聚合酶和联合酶的作用进行修复。
突变和修复PPT课件
第三章 突变和修复
Tay-sachs 家族性白痴病(Tay-sachs disease):是因为在其细胞 内缺乏氨基己糖脂酶,不能将神经节苷脂GM2 加工成 为GM3,结果大量的GM2累积在神经细胞中,导致中 枢神经系统退化
脱氨氧化
CMe
T
C
U
13
第二节 突变剂(续)
C→U的错误可由尿嘧啶糖 基酶系统修复
CMe→T的错误,修复系统 的修复效率较低。如果在 新一轮复制前未能完成修 复,则产生突变
14
第三节 DNA的修复
一、复制修复: 1. 尿嘧啶-糖基酶系统: 修复对象:掺入到DNA链的U(能与A配对,PolIII
无法修复) 修复过程:
27
4.SOS修复(续)
SOS反应由Rec A蛋白和Lex A蛋白相互作用引起。
Rec A被(相对分子质量为22700)称为辅蛋白酶(coprotease), 在有单链DNA和ATP存在时,Rec A蛋白被激活而促进LexA自身 的蛋白水解酶活性。
LexA蛋白是许多基因的阻遏物,当它被RecA激活自身的蛋白水 解酶活性后自我分解,使一系列SOS基因得以表达,其中包括:
6
二、突变的分类(续)
自发突变(spontaneous mutatiow) 突变率 是指在单位时间内某种突变发生的概率,即每 代每对核苷的突变概率数或每代每个基因的突 变概率。 突变频率是指在一个细胞群体或个体中,某种突变发 生的数目,即每10万个生物中发生突变体的 数目,或每百万个配子中突变的数目
第九章遗传学习题答案

第九章突变和重组机理一、名词解释1.碱基替换:一个碱基对被另一个碱基对代替。
又分2种:转换和颠换。
2.染色单体转变:减数分裂的4个产物中,有一个产物发生基因转变。
3.显性致死:只有一个致死基因就引起致死效应的,杂合态即有致死效应。
4.Muller-5品系的果蝇:X染色体上有显性棒眼基因B和隐性杏色眼基因Wa,还有一些倒位,可以抑制 Muller-5的X染色体与野生型X染色体的重组。
5、基因突变(点突变):指一个基因变成了它的等位基因。
6、自发突变:由外界环境条件自然作用或生物体内的生理生化变化发生的突变。
7、诱发突变:在特设的诱变因素(物理、化学、生物等)诱发下发生的突变。
8、显性致死:杂合态即有致死效应。
9、隐性致死:纯合态时才有致死效应,常见条件致死突变:在某些条件下是成活的,而在另一些条件下是致死的突变。
11、突变率:指生物体(微生物指每一个细胞)在每一世代中,每一基因组或每一细胞发生突变的概率。
12、易变基因:比一般基因容易突变的基因。
13、诱变剂:凡能增加突变率的物质都叫诱变剂。
14、转换:DNA分子中的嘌呤被嘌呤或嘧啶被嘧啶替换。
15、颠换:DNA分子中的嘌呤被嘧啶或嘧啶被嘌呤替换。
16、DNA修饰物:通过化学变化改造DNA分子结构的物质。
其作用与DNA复制无关。
17、染色单体转变:减数分裂的4个产物中,有一个产物发生基因转变。
18、半染色单体转变:减数分裂的4个产物中,有1个或2个产物的一半出现基因转变。
影响DNA 的一条链,分离一定发生在减数分裂后的有丝分裂中,所以叫做减数后分离。
二、选择填空1、紫外线照射主要引起( B )。
A DNA分子的断裂B 胸腺嘧啶二聚体C 尿嘧啶二聚体D DNA分子交连2、辐射剂量增加一倍,突变频率(D )。
A 不变 B增加2倍 C 增加3倍 D 增加1倍3、吖啶类分子扁平、能插入DNA相邻碱基对间,使碱基分开,从而使DNA分子双链歪斜,导致交换时出现(A )。
基因突变
突变的特点:
1. 随机性 2. 稀有性 3. 可回复性:真回复突变和第二点回复 突变
正向突变
his+
回复突变
his-
4. 多方向性 5. 通常为有害性
2007年
57.已知某一腺嘌呤缺陷型的粗糙脉胞菌受到离原 突变25个图距单位的一个抑制基因的作用后回复 为原养型,假如将这一菌株与野生型杂交,那么 后代中腺嘌吟缺陷型所占的百分数为: A.0 B.12.5% C.25% D.50% 解: ade-su- X ade+su+ 亲本型后代:ade-su-(原养型), ade+su+(原养型) 重组型后代:ade-su+(腺嘌呤缺陷型), ade+su-(原 养型), 重组型后代合计应占25%,各占12.5%。(B)
点突变的类型(以 Tyr 的密码子为例) 无义突变 DNA TAC→TAA , TAG ↓ ↓ ↓ ↓ RNA UAC UAA UAG ↓ ↓ ↓ ↓ aa Tyr Och Amb 同义突变 TAC → TAT ↓ ↓ UAC UAU ↓ ↓ Tyr Tyr 错义突变 TAC→ TCC ↓ ↓ UAC UCC ↓ ↓ Tyr Ser
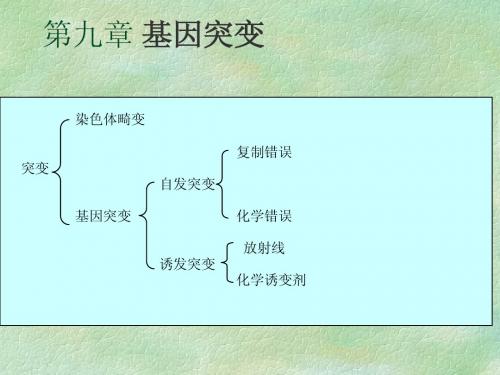
第九章 基因突变
染色体畸变 复制错误 突变 自发突变 基因突变 诱发突变 化学诱变剂 化学错误 放射线
直接修复
修复
切除修复
复制后修复
DNA Pol 3'→5'外切酶活性 光复活酶 一般切除修复——UvrABC 系统 AP 内切酶 特殊切除修复 糖基酶 GO 系统-MutM,MutY,MutT 错配修复-Dam, MutL,MutH,UvrD 重组修复-RecA SOS 修复-RecA,LexA,UvrAB,UmuC,HimA
遗传学期末总结详解演示文稿

第十三页,共59页。
第十一章 免疫遗传
1. 抗原(antigen): 凡是能够刺激机体的免疫系统产生抗体,并且能够和相应的抗体发 生特异性结合反应的物质
2. 新生儿溶血症 • 由胎儿与母体红细胞抗原不相容所引起的。 • 胎盘渗血、胎盘剥离时胎儿红细胞进入母体,母体产生全抗体IgG。IgG抗体通
2. 如何区别核外遗传和母性影响;
3. 植物的雄性不育概念及类型
4. 不育系、保持系、恢复系的关系
第二十五页,共59页。
第十三章 核外遗传
1. 核外遗传的概念和特点
细胞质内遗传物质控制的遗传现象和遗传规律。--细胞质遗传
1. 正交和反交的遗传表现不同,F1通常只表现母本的性状;
2. 与父本连续回交母本核基因可被全部置换掉,但母本细胞质基因变化不大;
5. C值悖理:
C值和进化程度之间没有严格的对应关系
6. N值悖理: 基因数目与其生物结构的复杂性不成比例的现象
7. 基因组学:
研究基因组的组成、结构和功能的学科
基因组学按研究内容:
结构基因组学
功能基因组学
蛋白质组学
第九页,共59页。
8. 结构基因组学:
是指通过基因作图、核苷酸序列分析,确定基因的组成、进行基 因定位的学科。
首先是重链发生基因重排,随后是轻链重排。
重链:胚系基因 → D-J连接 → V-DJ连接
7. 等位排斥与同种型排斥
• 等位排斥(allelic exclusion):一条染色体上Ig重链基因的有效重排,抑
制另一条同源染色体重链基因重排。 • 同种型排斥(isotype exclusion):是指两种轻链之间的排斥,轻链有κ链和λ链
基础生化-2009-第九章DNA合成

的两个子代DNA。细菌的旋转酶是Ⅱ型的代表。
(三) 引物酶(primrase)
合成RNA引物,又叫引物合成酶、引发酶。
它以单链DNA为模板,以ATP、GTP、CTP、UTP为原料,
从5→3方向合成出RNA片段,即引物。
(四) 切除引物的酶
原核细胞是DNA聚合酶Ⅰ或RNaseH(水解与DNA杂交的 RNA链)。 真核细胞是RNaseHⅠ
爬行模式
四、复制的机制
(一)θ复制(大肠杆菌为代表) (二)滚环复制 (三)D-环复制 (四)真核细胞的复制
滚环复制(rolling circle replication)是一些简单 低等生物或染色体以外的DNA复制的特殊形式。
3'
3'' 3 5'
5'
5'
五、复制的过程(大肠杆菌为例)
(一) 复制的起始第九章 DNA的生物合成• 第一节 DNA的复制
• 第二节 DNA的损伤修复
• 第三节 反转录
第一节 DNA的复制 一、复制的一般特点
1.模板、前体、Mg2+。 2.DNA的解链 3.半保留复制 4.需要引物
5.链延伸的方向
6.固定的起点(复制起始区有共有特征)
7.复制的方向(双、单)
8.半不连续复制
光修复
UV
光修复酶 (photolyase)
细菌、真菌、植物、很多脊椎动物有该酶,但胎盘类哺乳动 物没有。
2.烷基化碱基的直接修复(P601了解) 烷基转移酶,6-甲基鸟嘌呤甲基转移酶最常见。该酶以“自杀” 方式催化反应。 其活性中心的Cys残基作为甲基受体,但得到甲基就失活了。随
后被选择性降解。
二、 复制的方式——半保留复制
第九章 DNA的生物合成
主要内容
• 概述 • DNA的生物合成 • DNA的损伤与修复
概
述
遗传信息传递的中心法则
反映了从DNARNA蛋白质的遗传信息主 流,揭示了生物体内遗传信息的贮存、传递和 表达的规律。
复制
DNA
转录
RNA
翻译
蛋白质
反转录
复制
RNA (病毒)
翻译
蛋白质 (病毒)
第一节
★ 定义:
DNA的复制
复制是指以亲 代DNA为模板合成 子链DNA的过程。
四、DNA的复制过程
大致分为三阶段: 复制的起始 链的延长 复制的终止
(一)复制的起始
1. DNA解成单链
由特定蛋白质识别复制起始位点(ori)解螺 旋酶、TOPO酶及单链DNA结合蛋白的共同作用下, DNA解链,解旋,形成复制叉 2. 引发体的生成 解旋酶解开双链后引物酶进入形成引发体
3. RNA引物的合成 依赖于单链模板,由引物酶催化按碱基配对 规律合成一小段RNA引物(原核细胞引物长50-100 个碱基,真核约10个碱基)。
(4)在RNA引物上合成DNA
DNA复制的精确性(高保真复制)
DNA复制必须具有高度精确性,在大肠杆菌的细 胞DNA复制中其错误率约为1/109~1/1010,即每109~ 1010个核苷酸才出现一个错误,也就是大肠杆菌染色 体DNA复制1000~10000次才出现一个核苷酸的错误。 这么高的精确性的保证主要与下列因素有关: 1、碱基的配对规律:摸板链与新生链之间的碱 基配对保证碱基配错几率约为1/104~1/105。 2、DNA聚合酶的3’→5’外切酶活性的校对功能, 使碱基的错配几率又降低100~1000倍。 3、DNA的损伤修复系统。
(1)DNA聚合酶 即依赖于DNA的DNA聚合
突变类型与修复要点
这种突变不能引起氨基酸序列的改变。
9
碱基置换造成多肽链中一个氨基酸的改变, 但该氨基酸不能影响多肽链的正常功能, 因此不改变微生物的遗传性状。
10
移码突变是在DNA分子上的密码子中添加 或丢失一个或几个碱基,其结果造成从改 变的碱基开始所有其后的密码子碱基都往 后移动,使密码子杂乱而重新编组,使多 肽链上的氨基酸序列发生很大的改变,并 出现明显的遗传性状变异。
错义突变通常在第一或第二个碱基发生变 化时,容易引起氨基酸种类改变。
决定某氨基酸的密码子被终止密码子 (UAG)代替,因而mRNA翻译多肽链 过程中出现句号,使翻译工作中途停止, 难以完成一条完整的多肽链的合成,这种 肽链是没有活性的。
8
指DNA分子上的遗传密码由于置换而成为 新的密码子,但是这种新密码子构成的氨 基酸与原有密码子所构成的氨基酸相同。
最典型的是营养缺陷型,它从野生型基因突变形成。其特点是 由于突变而失去合成某种代谢物质的能力,如氨基酸、维生素 等,当环境中缺乏这种物质它就不能生长繁殖。反之,只有给 它补充了这种物质,才能具有正常的生命活动。
还有糖类分解发酵突变株、色素形成突变株及有益代谢产物生 产能力突变株。
突变体形成的三个步骤:
缺陷型
15
形态突变型 生化突变型 条件致死突变型 致死突变型 抗性突变型
形态突变是一种可见突变,包括:
菌落——形状大小、颜色、表面结构、孢子 数量和颜色;
细胞形态——鞭毛、荚膜、菌体形状、大小、 孢子形态和大小;
细胞结构——细胞膜透性等; 噬菌体——噬菌斑形状、大小及清晰度等。
(suppressor tRNA)。 抑制tRNA不是细胞对无义突变应答的产物,而是自发
DNA的损伤修复及突变PPT课件
着色性干皮病(xeroderma pigmentosis,XP) 是一种切除修复有缺陷的遗传性疾病。
在研究其发病机制时,发现一些相关的基 因,称为 XPA、XPB、XPC等。这些基因的表达产物起辨认 和切除损伤DNA作用的。
XP病人是由于XP基因有缺陷,不能修复紫外 线照射引起的DNA损伤,因此易发生皮肤癌。
•19
➢ DNA链断裂 脱氧核糖破坏或磷酸二酯键断开而导致DNA链断裂。 一条链断裂称单链断裂(single strand broken); DNA双链在同一处或相近处断裂称为双链断裂(double strand broken )。
•20
➢交联(binding) 同一条DNA链上或两条DNA链上的碱基间以共价
烷化剂的种类很多,常见的有甲磺酸乙酯(EMS)、 亚硝基胍(NG)和芥子气等。
•27
EMS能使鸟嘌呤的 N位置上有乙基,成为7一 乙基鸟嘌呤。与胸腺嘧啶配对,故能使G-C转换成 A-T。
烷化剂的另一作用是脱嘌呤。例如烷基在鸟嘌 呤N位上活化糖苷键引起断裂,使嘌呤从DNA链上 脱掉,产生缺口。复制时,与缺口对应的位点上可 能配上任一碱基,从而引起转换或颠换;而且去嘌 呤后的DNA容易发生断裂,引起缺失或其他突变。
•46
• 管理基因( caretaker genes) : 执行DNA的损伤修复, 维持基因组的完整性。如着色性干皮病的修复基因 XPA→XPF。
• 看门基因( gatekeeper genes) : 控制细胞信号传导, 调控细胞的增殖、分化和凋亡。如p53、patched基 因和ras等。皮肤癌的发生与看门基因突变关系密 切。
移码突变: 由于插入或缺失突变引起DNA的阅读框(ORF)
发生改变,从而产生不同蛋白质的过程。
基因突变及修复课件
将待分离突变株的原 始菌株以合适的稀释 度涂布到野生型菌株 和突变株均能生长的 平板(含完全培养基), 培养形成单菌落
通过一消毒“印章”将 平板菌落分别原位移 位到含完全培养基的 平板(c)和某一营养物 质缺乏的平板(d)
培养后对照两个平板上形成的单菌落,在完全培养基的平板 (c)上生长而营养物质缺乏的平板(d)上不生长的菌落即所需分 离的突变型,挑选菌落在完全培养基上分离纯化
基因突变及修复
一、基因突变的类型及其分离 二、基因突变的分子基础 三、DNA损伤的修复
基因突变及修复
二、基因突变的分子基础
突变
自发突变
环境因素的影响,DNA复制过程的偶 然错误等而导致,一般频率较低,通 常为10-10~10-6
诱 变 某些物理、化学因素对生物体的DNA
进行直接作用,突变以较高的频率产生
2、诱发突变(induced mutation) 常 用 诱 变 剂
(1) 碱基类似物:5-溴尿嘧啶(胸腺嘧啶类似物)和2-氨基嘌呤(嘌 呤类似物)
(2) 插入染料:插入DNA分子的碱基对,使其分开,导致DNA 在复制过程中滑动→移码突变,如溴化乙锭、吖啶橙等染料
(3) 直接与DNA碱基起化学反应的诱变剂:如亚硝酸、羟胺和烷 化剂等
基本培养基+ 少量组氨酸 +少量诱变剂
原理:(his-)菌株在不含 组氨酸的培养基中不能 生长,或只有极少数的 自发回复突变子生长, 如果回复突变率因某种 化学诱变剂(或待测物) 的作用而增加,那么这 种化学药物可判断为具 有致癌性
基因突变及修复
二、基因突变的分子基础
3、诱变剂与致癌物质——Ames试验
基因突变及修复
一、基因突变的类型及其分离
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
A· TG· transition C
2. 脱氨剂 (Deamination agents)
除去碱基上的氨基,改变其配对性质
3. 烷化剂 (Alkylating )
使DNA中的碱基发生烷化作用,引起特异性错配,或脱嘌呤。 包括芥子气,甲磺酸已酯(EMS),亚硝基胍(NG)等。如EMS,
自发损伤(spontaneous lesions)
自然产生的DNA损伤引起突变
1. 脱嘌呤:最为常见,DNA分子中碱基和脱氧核糖间的糖 苷键受到破坏,从而引起鸟嘌呤(G)或腺嘌呤(A)从DNA分子 上脱落下来。
常引起缺失或插入突变
2. 氧化性损伤:个体自然产生的氧化基,氧化物如超氧 自由基,氢氧自由基及过氧化基等,能对DNA造成氧化性损 伤,引起突变。
填补上。它可以修复一系列损伤,包括环丁烷二聚
体、6-4光产物和链间交联等引起的DNA变形 (major distortion)。尽管这些损伤也可以通过 途径进行修复,但NER是它们的主要修复手段。其 他能够引起DNA产生显著变形的损伤也可以通过该
途径进行修复。但是,这种机制不能修复DNA上的
错配碱基,以及仅造成DNA微小变形的碱性类似物 和甲基化碱基。
-5个核苷酸,5’切割点距损伤位点8个核苷酸。
UvrD与5’端的切点结合。UvrD是一种解旋酶(又称DNA
helicase II),它解开两个切点之间的DNA双螺旋,
导致一段短的带有损伤的ssDNA和UvrC被释放出来,
此时UvrB仍结合于另一条单链DNA分子上,可能是防
止单链被降解,也可能是指导DNA聚合酶I与缺口的3’ OH结合,合成一段新的核苷酸片断填补缺口,同时释
放出UvrB,最后一个磷酸二酯键由DNA连接酶催化形
成。
核苷酸的切除修复
DNA解螺旋酶
核苷酸切除修复(2)
uvrB、uvrA 蛋白结合 uvrD 切除
•
非条件型突变: 在任何条件下均可表现的突变 条件型突变: 突变的表现 = 突变基因型 + 诱导条件
根据突变的方式:
碱基替换(base substitution),DNA分子中一个碱 基被另一个碱基替代 又称点突变 插入(insertion),DNA序列中插入一个或多个碱基 缺失(deletion),DNA序列上缺失一个或多个碱基 重复(duplication),一段碱基序列发生一次重复 易位(translocation),一段碱基序列从原来的位 置移出,并插入到基因组的另一位置 倒位(inversion),一段碱基序列发生倒转,但仍 保留在原来的位置上
二、烷基的转移
一些酶可将烷基从核苷酸转移到自身的多肽链上,
例如,在人类细胞中发现有一种O6-甲基鸟嘌呤甲基
转移酶,能直接将DNA链鸟嘌呤O 6 位上的甲基移到 蛋白质的半胱氨酸残基上而修复损伤的DNA。
三、核苷酸切除修复
核苷酸切除修复需要移去一段包括损伤在内的核 苷酸序列,然后再通过DNA合成把余下的单链缺口
4 通读(reading through):碱基替换使终止密码子突
变为编码氨基酸的密码子,转录出的mRNA在翻译时不能适时 终止。
按突变发生的原因分类 • 自发突变(spontaneous mutation):在自然状况 下发生的突变。
• 诱发突变(induced mutation):外界因素诱发 引起的突变。
改变了野生型性状的突变,使性状由 野生型变为突变型。
回复突变(reverse mutation):
使突变型性状恢复到野生型性状的 突变。 大多数回复突变都发生在另一位点。因此,这样的突变 并未恢复野生型的序列,只是其表型被抑制了。第二点 突变可以发生在同一基因上,也可以发生在不同的基因 上,前者称为基因内抑制,后者称为基因间抑制。
使G的N位置带有已基(已烷化),成为7-已基鸟嘌呤,不与C配
对而与T配对,使G-C转换成A-T。
4.嵌入剂(intercalating agents) 吖啶橙(acridine orange)、原黄素( proflavine) 和溴化乙啶(ethidium bromide)
吖啶类化合物是一种平面三环分子, 其大小和形状与一个碱基对相似, 插入DNA分子中两个相邻的碱基之 间,使得原来相邻的碱基对分开一 定的距离,致使DNA在复制时增加
1. 基因内抑制
氨基酸上所带的电荷影响蛋白质构象
氨基酸侧链的大小影响构象
2.基因间抑制
如:无义突变的表型 可被tRNA基因的突变 所抑制
基因间抑制常发生在 tRNA或tRNA功能相 关基因上。
错义抑制突变( Missense suppressor )
GGA CCU
Gly
AGA UCU
Arg Gly
碱基替换:
根据碱基变化的不同,可以分为:
转换(transition):嘌呤与嘌呤之间,或嘧啶 与嘧啶之间的替换 颠换(transversion):嘌呤与嘧啶之间的替换。
缺失、插入 缺 失 和 插 入 常 导 致 移 码 突 变 ( frameshift mutations),结果产生截短的或异常的多肽链。
CCU
UCU
Gly
Gly
基因间移码抑制突变
基因内和基因间的错义(无义)、移码抑制突变均 由相应的错义(无义)、移码突变抑制
突变热点(hot spot),
突变可以发生在基因组中的任一位点。但是在基因组中,也 存在一些位点,这些位点发生突变的几率比随机分布所估计的
要高出许多,可能是预期的10倍或100倍,这些位点被称为突
第九章 突变、修复与重组
第一节 突变
第二节 修复 第三节 重组
本章要点:
• 突变的概念、类型 • DNA修复的主要方式 • 了解重组的分子基础
第一节:突变(mutation) 一、突变的主要类型
突变是DNA碱基序列水平上的永久的、可遗传的改变。 广义的突变包括染色体畸变和基因突变 狭义的突变通常指基因突变 根据突变发生的细胞类型、对基因功能的影响、突变的方 式、发生的原因等可对突变进行不同的分类
变热点,发生在热点上的突变常常是相同的。 大多数突变热点是DNA分子中的5-甲基胞嘧啶位点。
5mC处的突变率明显高于其它碱基处
增变基因(mutator gene): 突变后可使整个基因组中的突变率明显上升 的基因。
增变基 因大多 与DNA 复制或 修复有 关。
第二节 DNA的修复
一系列物理或化学因素可以对DNA造成化学损伤。这
修复过程需要多种酶的一系列作用,其中包括UvrA,
UvrB, UvrC 和 UvrD。由2个UvrA亚基和1个UvrB亚基
构成的复合体,非特异性地结合在DNA分子上,并沿
DNA分子滑动,对DNA进行扫描,其中UvrA负责检测螺 旋中的扭曲,一旦抵达扭曲部位,UvrA就会退出复合 体。UvrB募集UvrC, UvrC具有核酸内切酶活性,它切 断损伤位点两侧的磷酸二酯键,3’切割点距损伤位点4
(1)物理诱变剂 • 紫外线 • X-射线和γ-射线
紫外线:波长为100~400 nm的电磁辐射,非电离辐射, 直接作用于DNA。 嘧啶二聚体 胸腺嘧啶 X-射线和γ-射线:电离辐射 • 直接作用于DNA,对DNA产生损伤 产生多 种突变,并且常常造成DNA重排,例如缺失、 倒转和移位。 • 作用于水分子以及其他分子产生离子和自由基, 自由基对DNA分子产生广泛损伤。
一条子代DNA分子上的C ∶ G被 T∶ A所取代。细胞在 进化过程中,形成了多种修复机制,以保证在损伤阻 遏复制或者产生突变之前就识别并修复损伤。
一、光 复 活( Photoreactivation )
在可见光存在的情况下,DNA光解酶(DNA photolyase) 把环丁烷嘧啶二聚体分解为单体。DNA光解酶,又称光 复活酶(photoreactivating enzyme)。光解酶在暗 中结合到环丁烷二聚体上,吸收300~350 nm的光后被 激活,裂解二聚体后与DNA分子脱离。从光复活修复过 程可以看出,光解酶不是将嘧啶二聚体替换掉,而是 将两个嘧啶环之间的非正常化学键切开,恢复到原来 的形式。由于这种修复作用只在可见光下才可发生, 所以这种修复机制称为光复活。
自发突变的分子基础
• DNA复制错误
• 自发损伤
• 转座因子
DNA复制错误
• 遗传物质是DNA,DNA复制是半保留复制,如果发生 错误,引起碱基替换(base substitution), 造成 DNA遗传信息的改变。从而导致基因突变。
正常情况下,A-T配对,氨基态的腺嘌呤(A)只与胸 腺嘧啶(T)配对,但有时可转变成稀有的亚氨基形式, 可以与胞嘧啶配对,形成A-C, 再经一次复制,DNA分 子中的A-T对变成了G-C对.
根据突变发生的细胞类型:
• 生殖细胞突变(Germline mutation) • 体细胞突变(Somatic mutation)
可遗传给后代
仅表现于当代,癌症
根据对基因功能的影响
• 功能失活突变(loss of function mutation):
使基因功能失活或显著下降的突变。 • 功能获得突变(gain of function mutation): 使基因功能增强或不受抑制的突变。 根据突变的表达类型 •
(2)化学诱变剂
1
碱基类似物:
与碱基结构类似,可替代正常碱基掺入DNA 分子,引起碱基替换。 如5-溴尿嘧啶(BU)是胸腺嘧啶T的类似物,可 掺入DNA分子中。 酮式BU与A配对,而烯醇式与G配对 容易引起G-C对与A-T对的互相转换。。
碱基类似物还有5-溴脱氧尿苷(BrdU),5-尿嘧啶,5-氯尿 嘧啶及其脱氧核苷,2-氨基嘌呤(2-AP)等
正常人 红细胞