第15章 合成生物学
生物信息学智慧树知到课后章节答案2023年下华东理工大学

生物信息学智慧树知到课后章节答案2023年下华东理工大学华东理工大学第一章测试1.随着人类基因组计划的完成,以下哪些基因组计划是近期启动的计划答案:中国十万人基因组计划;G10K;我们所有人计划;英国十万人基因组计划2.统计学是一门独特学科,不是生物信息学研究工具和手段之一。
答案:错3.生物信息学研究任务之一包括SNP的发现和鉴定,对于疾病机理和药物开发靶点发现具有重要意义。
答案:对4.随着越来越多大规模测序项目的完成,其中最重要的科学使命之一就是要通过比较基因组学方法了解物种的起源和进化过程答案:对5.高等生物基因组中含有大量的非编码区,以及可能含有大量的外源病毒序列,只有通过生物信息学方法,解析其中功能和区域,为将来可能通过基因组编辑技术进行疾病机制解析提供基础答案:对第二章测试1.国际核酸数据库由EMBL,DDBJ和GenBank组成,它们在1988年形成国际核酸数据库联合中心,对数据进行答案:三方共享;数据同步更新;数据格式相同2.GenBank对于核酸数据的显示方式有以下几种答案:ASN.1;FASTA;Graph;GBK3.Uniprot KB对于生物数据在不同数据库中的链接、调用和标签转换具有非常重要的作用答案:对4.生物信息学的研究对象中包括各种数据库,比如答案:Uniprot;PDB;GenBank;KEGG5.BLAST是基于局部比对算法,采用渐进式比对方法,对数据分成字段等思路进行的成对比对方法答案:对第三章测试1.全局序列比对用于整体相似性程度较低、在较小区域内有局部相似性的两个序列比对。
答案:错2.以下哪些属于蛋白质打分矩阵?答案:PAM矩阵;遗传密码矩阵;疏水矩阵;BLOSUM矩阵3.传统的基于动态规划的局部性比对性算法采用的是精确的序列比对,虽然有着较好的比较结果,但是算法的时间复杂度较高。
答案:对4.在蛋白质数据库中比对蛋白质序列,需要选择一下那种blast模式?答案:blastp5.多序列比对就是两条以上的序列进行比对,可以用于进化树分析、寻找保守区域等。
重庆医科大学刘先俊《生物化学》第15章.血液的生物化学

当人从海平面上到4500 米高原后两天内, 2,3 BPG的浓度有原来的5mM 增加到8mM,结果对组织 的供氧量又恢复到接近 正常水平(0.38)。
如果BPG仍维持5mM水 平,对组织的供氧量将 减少四分之一,为0.30。
本章总结 血浆蛋白的电泳分类;非蛋白氮的 概念及主要种类。
{ CO2的运输 (6%) 碳酸氢盐形式
{ 化合结合
(约2/3 )
(94%) 氨基甲酸血红蛋白
(约1/4 )
1. 碳酸氢盐形式的运输
2.氨基甲酸血红蛋白的形式运输
HbNH2+CO2
在组织中
(氨基甲酸血红蛋白)
HbNHCOOH
肺
HbNHCOO- + H+
HbH+
+ HbO2
O2
Bohr效应的总结
CO2+H2O = H++HCO3-
结果使得细胞内的pH降低。低pH使Hb对氧的亲和 力降低。
波耳效应提高了氧转运系统的效率。在肺部,CO2 水平低,氧很容易被血红蛋白占有,同时释放出质子;
而在代谢的组织中,CO2水平相对来说比较高,pH较低,
O2容易从氧合血红蛋白中卸载。
(三)运输二氧化碳
Байду номын сангаас
物理溶解
运输O2、CO2、H+ (一)运输O2
与Hb结合占98.5% 氧的运输
物理溶解占1.5%
Hb与O2呈可逆结合
O2分压高(肺)
Hb+O2
HbO2
O2分压低(组织)
(还原、紫蓝色)
(氧化、红色)
Hb氧饱和曲线呈S型,两端斜率小,中间大; Mb氧饱和曲线呈双曲线型
生理意义:
(二)波尔效应(Bohr effect)
分子生物学--第15章 DNA损伤与修复

2017年3月科学杂志论文:癌症发生的原因中运气不 好占66.1%。
目录
DNA复制错误: 1. 碱基错配 互变异构移位(tautomeric shift): 碱基发生了酮式-烯 醇式或氨基-亚氨基异构体互变,造成碱基配对发生改 变,使复制后的子链上出现错误。
目录
目录
2.复制打滑
目录
目录
DNA损伤的后果
DNA 修复机制 短期效应 生理功能紊乱 细胞死亡
基因组不稳定 信号传导异常
异常增生和代谢
基因表达异常
长期效应
衰老 肿瘤 疾病
目录
第二节 DNA损伤的修复
The repair of DNA damage
目录
托 马 斯 ·林 达 尔 ( TomasRobertLindahl ) 1938 年 1 月 28 日出生,瑞 典医学家,专门从事癌症 研究,挪威科学和文学研 究院的成员。发现碱基切 除修复。
目录
(一)突变是进化的分子基础 (二)只有基因型改变的突变 (三)突变是某些疾病的发病基础 (四)突变导致死亡
目录
二、DNA损伤修复障碍与肿瘤等 多种疾病相关
DNA损伤与肿瘤、衰老以及免疫性疾病 等多种疾病的发生有着密切的关联
目录
思考题:
试说明DNA损伤的几种类型及修 复合成主要方式的名称。 试述核苷酸切除修复的过程。
根据DNA结构的改变分为:
碱基脱落 碱基结构破坏 嘧啶二聚体形成
DNA单链或双链断裂
DNA交联
目录
碱基脱落
目录
碱基结构破坏导致碱基之间发生错配
EMS(甲基磺酸乙酯)
目录
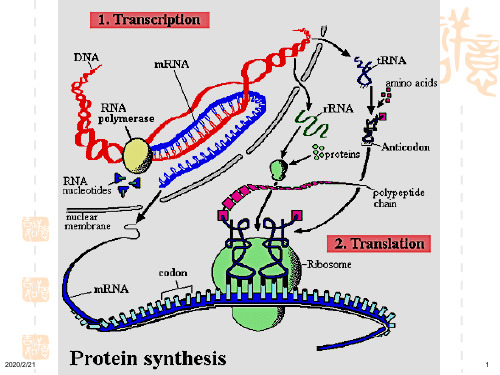
分子生物学第十五章 蛋白质的生物合成

1.遗传密码种类:
• mRNA 分子有4种碱基:A、G、C、U,可组合成64个密 码子,其中61个分别代表20种不同氨基酸
• 遗传密码共有43=64种, 64: UAA、UAG、UGA 终止密码 61: AUG 起始密码 代表Met 60: 代表19种氨基酸
9
遗传密码表 第一碱基
(5/-端)
第二碱基
43
一、起始阶段
(一)原核生物翻译起始复合物的形成
参与的物质: 核糖体50S和30S大小两类亚基 mRNA 起始作用的fMet-tRNAfMet GTP供能 起始因子(IF1、IF2、IF3)
44
起始过程:
• 核糖体大小亚基解离 • 小亚基与mRNA结合:
16sRNA识别mRNA的SD序列
• fMet-tRNAfMet与mRNA起始密码子AUG结合 • 核糖体大小亚基形成起始复合物
tRNAphe Phe-tRNAphe
• 肽链起始和延长的甲硫氨酰-tRNA:
真核生物:起始: Met-tRNAiMet 延长: Met-tRNAMet
原核生物: 起始:fMet-tRNAfMet 延长:Met-tRNAMet
39
40
第三节 肽链的合成过程
41
整个翻译过程可分为 :
• 翻译的起始 • 翻译的延长 • 翻译的终止
tRNA反密码子 第1位碱基
mRNA密码子 第3位碱基
I
U
G AC
U, C, A A, G U, C U G
17
摆 动 配 对
32 1
U
123
18
• ⑤遗传密码的通用性
从原核生物到人类都共用同一套遗传密码,被称为遗传密码的通用性。 密码的通用性进一步证明各种生物进化自同一祖先。
蛋白质的生物合成习题与参考答案

第十五章蛋白质生物合成一、填空题:1.三联体密码子共有64 个,其中终止密码子共有 3 个,分别为UAA 、UAG 、UGA ; 2.密码子的基本特点有四个分别为从5′→3′无间断性、简并性、变偶性、通用性; 3.次黄嘌呤具有广泛的配对能力,它可与U 、 C 、 A 三个碱基配对,因此当它出现在反密码子中时,会使反密码子具有最大限度的阅读能力;4.原核生物核糖体为70 S,其中大亚基为50 S,小亚基为30 S;而真核生物核糖体为80 S,大亚基为60 S,小亚基为40 S;5.原核起始tRNA,可表示为tRNA f甲硫,而起始氨酰tRNA表示为f Met-tRNA f甲硫;真核生物起始tRNA可表示为tRNA I甲硫,-tRNA表示为Met-tRNA f甲硫;6.肽链延伸过程需要进位、转肽、移位三步循环往复,每循环一次肽链延长 1 个氨基酸残基,原核生物中循环的第一步需要EF-Tu 和EF-Ts 延伸因子;第三步需要EF-G 延伸因子;7.原核生物mRNA分子中在距起始密码子上游约10个核苷酸的地方往往有一段富含嘌呤碱基的序列称为Shine-Dalgrano序列,它可与16S-rRNA 3′-端核苷酸序列互补;8.氨酰-tRNA的结构通式可表示为:OtRNA-O-C-CH-RNH2,与氨基酸键联的核苷酸是A腺嘌呤核苷酸;9.氨酰-tRNA合成酶对氨基酸和相应tRNA都具有较高专一性,此酶促反应过程中由ATP 水解提供能量;10.肽链合成的终止阶段, RF1因子和RF2因子能识别终止密码子,以终止肽链延伸,而RF3因子虽不能识别任何终止密码子,但能协助肽链释放;11.蛋白质合成后加工常见的方式有磷酸化、糖基化、脱甲基化、信号肽切除; 12.真核生物细胞合成多肽的起始氨基酸为甲硫氨酸,起始tRNA为tRNA I甲硫,此tRNA 分子中不含T C 序列;这是tRNA家庭中十分特殊的;二、选择题只有一个最佳答案:1.下列有关mRAN的论述,正确的一项是 CA、mRNA是基因表达的最终产物B、mRNA遗传密码的阅读方向是3′→5′C、mRNA遗传密码的阅读方向是5′→3′D、mRNA密码子与tRNA反密码子通过A-T,G-C配对结合E、每分子mRNA有3个终止密码子2.下列反密码子中能与密码子UAC配对的是 DA、AUGB、AUIC、ACUD、GUA3.下列密码子中,终止密码子是 BA、UUAB、UGAC、UGUD、UAU4.下列密码子中,属于起始密码子的是 AA、AUGB、AUUC、AUCD、GAG5.下列有关密码子的叙述,错误的一项是 CA 、密码子阅读是有特定起始位点的B 、密码子阅读无间断性C 、密码子都具有简并性D 、密码子对生物界具有通用性6.密码子变偶性叙述中,不恰当的一项是 AA 、密码子中的第三位碱基专一性较小,所以密码子的专一性完全由前两位决定B 、第三位碱基如果发生了突变如A G 、C U,由于密码子的简并性与变偶性特点,使之仍能翻译出正确的氨基酸来,从而使蛋白质的生物学功能不变C 、次黄嘌呤经常出现在反密码子的第三位,使之具有更广泛的阅读能力,I-U 、I-C 、I-A 从而可减少由于点突变引起的误差D 、几乎有密码子可用U C XY 或U C XY 表示,其意义为密码子专一性主要由头两个碱基决定7.关于核糖体叙述不恰当的一项是 BA 、核糖体是由多种酶缔合而成的能够协调活动共同完成翻译工作的多酶复合体B 、核糖体中的各种酶单独存在解聚体时,同样具有相应的功能C 、在核糖体的大亚基上存在着肽酰基P 位点和氨酰基A 位点D 、在核糖体大亚基上含有肽酰转移酶及能与各种起始因子,延伸因子,释放因子和各种酶相结合的位点8.tRNA 的叙述中,哪一项不恰当 DA 、tRNA 在蛋白质合成中转运活化了的氨基酸B 、起始tRNA 在真核原核生物中仅用于蛋白质合成的起始作用C 、除起始tRNA 外,其余tRNA 是蛋白质合成延伸中起作用,统称为延伸tRNAD 、原核与真核生物中的起始tRNA 均为fMet-tRNA9.tRNA 结构与功能紧密相关,下列叙述哪一项不恰当 DA 、tRNA 的二级结构均为“三叶草形”B 、tRNA3′-末端为受体臂的功能部位,均有CCA 的结构末端C 、T C 环的序列比较保守,它对识别核糖体并与核糖体结合有关D 、D 环也具有保守性,它在被氨酰-tRNA 合成酶识别时,是与酶接触的区域之一10.蛋白质生物合成中多肽的氨基酸排列顺序取决于 CA 、相应tRNA 的专一性B 、相应氨酰tRNA 合成酶的专一性C 、相应mRNA 中核苷酸排列顺序D 、相应tRNA 上的反密码子11.下列有关氨酰- tRNA 合成酶叙述中,哪一项有误 CA 、氨酰-tRNA 合成酶促反应中由ATP 提供能量,推动合成正向进行B 、每种氨基酸活化均需要专一的氨基酰- tRNA 合成酶催化C 、氨酰-tRNA 合成酶活性中心对氨基酸及tRNA 都具有绝对专一性OD 、该类酶促反应终产物中氨基酸的活化形式为R -CH -C -O -ACC -tRNANH 212.原核生物中肽链合的起始过程叙述中,不恰当的一项是 DA 、mRNA 起始密码多数为AUG,少数情况也为GUGB 、起始密码子往往在5′-端第25个核苷酸以后,而不是从mRNA5′-端的第一个苷酸开始的C 、在距起始密码子上游约10个核苷酸的地方往往有一段富含嘌呤的序列,它能与16SrRNA3′-端碱基形成互补D、70S起始复合物的形成过程,是50S大亚基及30S小亚基与mRNA自动组装的13.有关大肠杆菌肽链延伸叙述中,不恰当的一项是 CA、进位是氨酰-tRNA进入大亚基空差的A位点B、进位过程需要延伸因子EFTu及EFTs协助完成C、甲酰甲硫氨酰-tRNA f进入70S核糖体A位同样需要EFTu-EFTs延伸因子作用D、进位过程中消耗能量由GTP水解释放自由能提供14.移位的叙述中哪一项不恰当 CA、移位是指核糖体沿mRNA5′→3′作相对移动,每次移动的距离为一个密码子B、移位反应需要一种蛋白质因子EFG参加,该因子也称移位酶C、EFG是核糖体组成因子D、移位过程需要消耗的能量形式是GTP水解释放的自由能15.蛋白质生物合成的方向是: BA、从C端到N端B、从N端到C端C、定点双向进行D、从C端、N端同时进行16.在蛋白质合成过程中,下列哪些说法是正确的 CA、氨基酸随机地连接到tRNA上去B、新生多肽链的合成都是从C-端向N-端方向延伸的C、通过核糖核蛋白体的收缩,mRNA不断移动D、肽键形成是由肽酰转移酶作用下完成的,此种酶不属于核糖体的组成成分17.70S起始复合物的形成过程的叙述,哪项是正确的 DA、mRNA与30S亚基结合过程需要超始因子IF1B、mRNA与30S亚基结合过程需要超始因子IF2C、mRNA与30S亚基结合过程需要超始因子IF3D、mRNA与30S亚基结合过程需要超始因子IF1、IF2和IF318.mRNA与30S亚基复合物与甲酰甲硫氨酰-tRNA f结合过程中起始因子为 AA、IF1及IF2B、IF2及IF3C、IF1及IF3D、IF1、IF2及IF319.原核细胞中氨基酸掺入多肽链的第一步反应是: DA、甲酰蛋氨酸-tRNA与核蛋白体结合B、核蛋白体30S亚基与50S亚基结合C、mRNA与核蛋白体30S亚基结合D、氨酰tRNA合成酶催化氨基酸活化20.假设翻译时可从任一核苷酸起始读码,人工合成的AACnn为任意整数多聚核苷酸,能够翻译出几种多聚氨基酸 CA、一种B、二种C、三种D、四种21.绝大多数真核生物mRNA5’端有 AA、帽子结构B、PolyAC、起始密码D、终止密码22.能与密码子ACU相识别的反密码子是DA、UGAB、IGAC、AGID、AGU23.原核细胞中新生肽链的N-末端氨基酸是CA、甲硫氨酸B、蛋氨酸C、甲酰甲硫氨酸D、任何氨基酸24.tRNA的作用是 DA、把一个氨基酸连到另一个氨基酸上B、将mRNA连到rRNA上C、增加氨基酸的有效浓度D、把氨基酸带到mRNA的特定位置上25.细胞内编码20种氨基酸的密码子总数为: DA、16B、64C、20D、6126.下列关于遗传密码的描述哪一项是错误的CA、密码阅读有方向性,5'-端开始,3'-端终止B、密码第3位即3′-端碱基与反密码子的第1位即5′-端碱基配对具有一定自由度,有时会出现多对一的情况C、一种氨基酸只能有一种密码子D、一种密码子只代表一种氨基酸27.蛋白质合成所需的能量来自CA、ATPB、GTPC、ATP和GTPD、CTP28.下列关于氨基酸密码的描述哪一项是错误的 AA、密码有种属特异性,所以不同生物合成不同的蛋白质B、密码阅读有方向性,5′-端起始,3′-端终止C、一种氨基酸可有一组以上的密码D、一组密码只代表一种氨基酸29.mRNA的5′-ACG-3′密码子相应的反密码子是 CA、5′-UGC-3′B、5′-TGC-3′C、5′-CGU-3′D、5′-CGT-3′30.下列哪一个不是终止密码 BA、UAAB、UACC、UAGD、UGA三、是非题在题后括号内打√或×:1、蛋白质生物合成所需的能量都由ATP直接供给; ×2、反密码子GAA只能辨认密码子UUC; ×3、生物遗传信息的流向,只能由DNA—→RNA而不能由RNA—→DNA; ×4、原核细胞新生肽链N端第一个残基为fMet,真核细胞新生肽链肽链N端第一个氨基酸残基为Met; √5、DNA复制与转录的共同点在于都是以双链DNA为模板,以半保留方式进行,最后形成链状产物; ×6、依赖DNA的RNA聚合酶叫转录酶,依赖于RNA的DNA聚合酶即反转录酶; √7、密码子从5’-端至3’-端读码,而反密码子则从3’-端至5’-端读码; ×8、一般讲,从DNA的三联体密码子中可以推定氨基酸的顺序,相反从氨基酸的顺序也可毫无疑问地推定DNA顺序; ×9、DNA半不连续复制是指复制时一条链的合成方向是5′→3′而另一条链方向是3′→5′; ×10、真核生物蛋白质合成起始氨基酸是N-甲酰甲硫氨酸; ×11、原核细胞的DNA聚合酶一般都不具有核酸外切酶的活性; ×12、在具备转录的条件下,DNA分子中的两条链在体内都可能被转录成RNA; ×13、核糖体是细胞内进行蛋白质生物合成的部位; √14、mRNA与携带有氨基酸的tRNA是通过核糖体结合的; √15、核酸是遗传信息的携带者和传递者; √16、RNA的合成和DNA的合成一样,在起始合成前亦需要有RNA引物参加; ×17、真核生物mRNA多数为多顺反子,而原核生物mRNA多数为单顺反子; ×18、合成RNA时,DNA两条链同时都具有转录作用; ×19、在蛋白质生物过程中mRNA是由3’-端向5’-端进行翻译的; ×20、蛋白质分子中天冬酰胺,谷氨酰胺和羟脯氨酸都是生物合成时直接从模板中译读而来的; ×21、逆转录病毒RNA并不需要插入寄主细胞的染色体也可完成其生命循环; ×四、问答题:1.氨酰-tRNA合成酶在多肽合成中的作用特点和意义;答:氨基酰-tRNA合成酶具有高度的专一性:一是对氨基酸有极高的专一性,每种氨基酸都有一种专一的酶,它仅作用于L-氨基酸,不作用于D-氨基酸,有的氨基酸-tRNA合成酶对氨基酸的专一性虽然不很高,但对tRNA仍具有极高专一性;这种高度专一性会大大减少多肽合成中的差错;2.原核细胞与真核细胞蛋白质合成起始氨基酸起始氨基酰—tRNA及起始复合物的异同点有那些答:为了便于比较列表如下3.原核生物与真核生物mRNA的信息量及起始信号区结构上有何主要差异;答:为了便于比较列表如下:4.简述三种RNA在蛋白质生物合成中的作用;答:1 mRNA:DNA的遗传信息通过转录作用传递给mRNA,mRNA作为蛋白质合成模板,传递遗传信息,指导蛋白质合成;2 tRNA:蛋白质合成中氨基酸运载工具,tRNA的反密码子与mRNA上的密码子相互作用,使分子中的遗传信息转换成蛋白质的氨基酸顺序是遗传信息的转换器;3 rRNA:核糖体的组分,在形成核糖体的结构和功能上起重要作用,它与核糖体中蛋白质以及其它辅助因子一起提供了翻译过程所需的全部酶活性;五、名词解释:1.遗传密码与密码子多肽链中氨基酸的排列次序mRNA分子编码区核苷酸的排列次序对应方式称为遗传密码;而mRNA分子编码区中每三个相邻的核苷酸构成一个密码子;由四种核苷酸构成的密码子共64个,其中有三个不代表任何氨基酸,而是蛋白质合成中的终止密码子;2.起始密码子与终止密码子蛋白质合成中决定起始氨基酸的密码子称为起始密码子,真核与原核生物中的起始密码子为代表甲硫氨酸的密码子AUG和代表缬氨酸的密码子GUG;3.密码的简并性和变偶性一种氨基酸可以具有好几组密码子,其中第三位碱基比前两位碱基具有较小的专一性,即密码子的专一性主要由前两位碱基决定的特性称为变偶性;4.核糖体与多核糖体生物系统中合成蛋白质的部侠,称为核糖体;多聚核糖体:一条mRNA模板链可附着10-100个核糖体,这些核糖体依次结合起始密码子,沿5-3方向读码移动,同时进行肽链合成,这种mRNA与多个核糖体形成的聚合物称为多聚核糖体;5.同功tRNA、起始tRNA、延伸tRNA用于携带或运送同一种氨基酸的不同tRNA称同功tRNA,能特异识别mRNA上起始密码子的tRNA,称为起始tRNA;在肽链延伸过程中,用于转运氨基酸的tRNA称为延伸tRNA;6.EFTu-EFTs循环,移位,转肽肽键形成EF-Tu与EF-T S称为延伸因子,参与氨基酰-tRNA进位,每完成一次进位需要EF-Ts-EF-Tu循环一周,其过程如下:移位:就是核糖体沿着mRNA从5′向3′-端移动一个密码子的距离:转肽则是位于核糖体大亚基P 位点的肽酰基在转肽酶的作用下,被转移到A位点,氨在酰-tRNA的氨基上形成肽键的过程;7.信号肽几乎所有跨膜运送的蛋白质结构中,多数存在于N-末端的肽片段称为信号肽,其长度一般为15—35个氨基酸残基;它在蛋白质跨膜运送中起重要作用;少数信号肽位于多肽中间某个部位,称为“内含信号肽;”8.移码突变在mRNA分子编码区内插入一个或删除一个碱基,就会使这点以后的读码发生错误,这称为移码;由于移码引起的突变称为移码突变;。
第十五章 蛋白质的合成-答案

一、选择题1.与原核生物核糖体小亚基结合的蛋白质合成抑制剂是(A )A.链霉素B.氯霉素C.利福霉素D.放线菌素E.青霉素3.大肠杆菌合成的所有未修饰的多肽链,其N末端的氨基酸残基应是(C)A.甲硫氨酸B.丝氨酸C.甲酰甲硫氨酸D.甲酰丝氨酸E.谷氨酸4.某5岁儿童突发咽痛,呼吸急促,表现烦躁。
体温测得为38.5T,咽部检查见扁桃体肿大,甚至观察有循环系统衰竭的症状,诊断为咽白喉,白喉病产生的外毒素(白喉毒素)为致病的主要因素。
白喉毒素抑制蛋白质合成的主要机制是(E)A.降解核糖体大亚基的28SrRNA使其失活B.特异性结合40S亚基的A位干扰延长阶段的进位C.抑制肽酰转移酶从而抑制肽链延长D.与40S亚基结合影响翻译准确性E.使eEf-2失活从而阻断肽链延长6.小明因为受伤需要抹点红霉素,红霉素作为一种抗生素其作用原理是(C)A.阻止翻译起始复合物的形成B.抑制氨基酸tRNA与小亚基结合C.抑制肽酰转移酶D.引起读码错误E.抑制EF-G10.多肽链的延长与下列物质无关的是(B)A.肽酰转移酶B.甲酰甲硫氨酰-tRNAC.GTPD.mRNAE.EF-Tu、EF-Ts和EF-G11.肽键形成部位是(B)A.核糖体大亚基P位B.核糖体大亚基A位C.两者都是D.两者都不是E.核糖体大亚基E位13.氨基酰-tRNA合成酶的特点是(E)A.存在于细胞核内B.只对氨基酸的识别有专一性C.只对tRNA的识别有专一性D.催化反应需GTPE.对氨基酸、tRNA的识别都有专一性15.多数氨基酸都有两个以上密码子,下列氨基酸只有一个密码子的是(D)A.苏氨酸、甘氨酸B.脯氨酸、精氨酸C.丝氨酸、亮氨酸D.色氨酸、蛋氨酸E.天冬氨酸、天冬酰胺19.翻译起始复合物的组成(C)A.DNA模板+RNA+RNA聚合酶B.Dna蛋白+开键DNAC.核糖体+甲硫氨酰tRNA+mRNAD.翻译起始因子+核糖体E.核糖体+起始者tRNA20.下列关于核糖体的叙述,正确的是(B)A.是遗传密码的携带者B.由rRNA与蛋白质构成C.由snRNA与hnRNA构成D.由引物、DNA和蛋白质构成E.由tRNA与蛋白质构成21,下列关于密码子的叙述,正确的是(C)A.由DNA链中相邻的三个核苷酸组成B.由tRNA中相邻的三个核苷酸组成C.由mRNA上相邻的三个核苷酸组成D.由rRNA中相邻的三个核苷酸组成E.由多肽链中相邻的三个核苷酸组成22.遗传密码的简并性是指(C)A.密码子的第3位碱基决定编码氨基酸的特异性B.一个密码子可代表多个氨基酸C.多个密码子可代表同一氨基酸D.密码子与反密码子之间不严格配对E.所有生物可使用同一套密码23.下列关于遗传密码的叙述,正确的是(E)A.遗传密码只代表氨基酸B.一种氨基酸只有一个密码子C.一个密码子可代表多种氨基酸D.密码子与反密码子遵守严格的碱基配对原则E.密码子的简并性降低了基因突变的效应24.一个tRNA的反密码子为5,UGC3,,它可识别的密码子是(A )A.5'GCA3'B.5,ACG3'C.5'GCU3'D.5'GGC3'E.5'AUG3,25.氨基酸通过下列哪种化学键与tRNA进行特异结合(B)A.糖苷键B.酯键C.酰胺键D.磷酸酯键E.氢键26.蛋白质生物合成中氨基酸的活化与tRNA的结合需要(B )A.氨基酸tRNA合成酶B.氨基酰tRNA合成酶C.ATP合成酶D.转位酶E.GTP27.参与新生多肽链正确折叠的蛋白质是(A)A.分子伴侣B.G蛋白C.转录因子D.释放因子E.p因子28.原核生物起始tRNA是 (C)A.甲硫氨酰-tRNAB.缬氨酰-tRNAC.甲酰化的甲硫氨酰-tRNAD.氨酰-tRNAE.乙酰化的甲硫氨酰-tRNA29.新生肽链合成的方向是 (A )A.从N端到C端合成B.从C端到N端合成C.没有固定的方向D.先从N端到C端合成小片段,再连接成一条多肽链E.从中间部位向N端和C端两个方向同时进行30.蛋白质磷酸化修饰的潜在修饰位点是(B)A.甘氨酸B.酪氨酸C.苯丙氨酸D.谷氨酸E.赖氨酸34.蛋白质合成终止是 (B)A.核糖体到达mRNA分子的3'末端B.释放因子识别终止密码子进入A位C.释放因子进入P位D.mRNA出现发夹结构,核糖体无法移动E.特异的tRNA进入A位35.氯霉素对细菌蛋白质合成的生物学影响是(E )A.导致未成熟多肽链的释放B.抑制核糖体大小亚基的结合C.抑制tRNA功能D.抑制30S核蛋白体亚基的活化E.抑制50S核蛋白体亚基的肽酰转移酶活性38.若向mRNA的编码区插入一个核苷酸,则会出现(D)A.翻译出的蛋白质的第1个氨基酸残基改变B.翻译出的蛋白质的氨基酸残基序列不变C.插入处上游翻译出来的氨基酸序列完全改变D.插入处下游翻译出来的氨基酸序列完全改变E.仅在插入处一个氨基酸残基改变。
第15章 蛋白质的生物合成
tRNA反密码子 第1位碱基
I
U G AC
mRNA密码子 第3位碱基
U, C, A A, G U, C U G
2020/2/21
21
Robert W. Holley 1922~1993
Har Gobind Khorana
1922~
Marshall W. Nirenberg
1927~2010
The Nobel Prize in Physiology or Medicine 1968
• 原核细胞中数个结构基因常串联为一个转录 单位,转录生成的mRNA可编码几种功能相关
的蛋白质,为多顺反子(polycistron) 。
2020/2/21
5
原核生物的多顺反子
5 PPP
3
真核生物的单顺反子
5 mG - PPP
蛋白质
3
蛋白质
非编码序列
核蛋白体结合位点
编码序列
起始密码子
终止密码子
"for their interpretation of the genetic code and
2020/2/21
its function in protein synthesis"
22
二、核糖体是多肽链合成的装置
2020/2/21
23
2020/2/21
24
2020/2/21
27
原核、真核生物核蛋白体的组成
4. 通用性(universal)
▪ 蛋白质生物合成的整套密码,从原核生 物到人类都通用。
▪ 已发现少数例外,如动物细胞的线粒体、 植物细胞的叶绿体。
▪ 密码的通用性进一步证明各种生物进化 自同一祖先。
生物化学第15章 蛋白质的生物合成

P397
(2)核糖体的功能 ❖ 详见蛋白质合成过程;
15. 1. 3 tRNA
P398
❖ 每一种组成蛋白质的氨基酸(20种),至少有一种 tRNA 负 责 转 运 , 大 多 数 氨 基 酸 具 有 几 种 用 来 转 运 的 tRNA;
❖ 一个细胞中常含有50或更多的不同tRNA分子; ❖ 书写时,将所运氨基酸写在tRNA的右上角,如
❖ 主要参与物: (1)EF-T:延长因子,有EF-Tu、EF-Ts 、EF-G; (2)能量:GTP; (3)新来的氨酰- tRNA;
核糖体上有接受肽基的P位和接受氨酰基的A位
1. P位:fMet-tRNAfMet在P位; 2. A位:新来的AA- tRNA进A位并成肽; 3. fMet-tRNAfMet移到A位成肽; 4 .肽基移到P位,A位空,又接受新来的氨酰- tRNA;
❖ 真核细胞蛋白质合成机制与大肠杆菌有许多相似之处;
蛋白质合成过程有许多机制保证蛋白质合成的 忠实性
15. 1. 2 核糖体 ❖ 核糖体是蛋白质合成位
❖ 核糖体存在于细胞质中或细胞内膜上: 1. 原核细胞中:核糖体或以游离形式分布在细胞质基质内,
或附着在细胞质膜内侧,或与mRNA 结合形成串状的多 核糖体; 2. 真核细胞中:核糖体既可游离存在,也可与细胞内质网 (真核细胞重要细胞器,属于细胞膜系统)结合,形成
GTP提供,约占生物合成反应总耗能量的90%;
蛋白质的生物合成快速而复杂
❖ 以E.coil为例,蛋白质占细胞干重50%左右。每个细胞 中约有3000种不同的蛋白质分子,每种蛋白质又有无数 分子,而大肠杆菌细胞的分裂周期不过20min,可见蛋 白质生物合成的速度之快;
❖ 蛋白质生物合成过程十分复杂,几乎涉及到细胞内所有 种类的RNA和几十种蛋白质因子;
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
15.3 合成生物学的研究方向
15.3.1 创建新的基因调控模块和线路
各种蛋白质、DNA、RNA的相互作用形成复杂 的表达调控网络。通过构建非天然的基因调控模 块设计构建细胞生命活动的分子网络。
用途:调节基因表达和蛋白质功能。
基因线路
1) 基因拨动开关
诱导物B
e.g. E. coli
里的意志力就可以操作电脑,能发送电子邮件。 未来在芯片的帮助下,人们可以直接用大脑
控制手臂和腿的假肢。现在就已经出现了可以部 分控制的假肢。
合成生物学工程化三原则:
标准化 抽象化 复杂系统去偶合
Drew Endy (MIT)
Endy created the comic book Adventures in Synthetic Biology to help explain his work to students and other scientists.
由于在生物合成抗疟疾药物的突出成就, Keas ling 被美国“发现”杂志评选为2006 年度最有影 响的科学家。
该项目已经获得比尔- 梅林达盖茨基临床实验等后续工作。
3. 代谢途径的快速进化 基因突变
改造代谢途径
生产目标化合物
Church 对20种番茄红素合成有关的基因进行突变; 将突变的90个DNA片段,转入大肠杆菌; 3天内产生了150亿基因突变体; 从中筛选到使番茄红素产量提高5倍的基因。
酵母的乙醇代谢工程 酿酒酵母是工业上生产乙醇的优良菌株,与 细菌相比具有较高的乙醇耐受力,对纤维素水解 液中的抑制物有较高的抗性。 缺点 酿酒酵母缺乏木糖转化为木酮糖所需的酶, 因而不能利用木糖,但它能利用木酮糖。对其菌 种改造涉及木糖跨膜运输、吸收利用、磷酸戊糖 途径、糖酵解及胞内氧化还原状态的维持等多个 方面。
的简单问题。
抽象化:将生物功能单元划分为不同层次。 DNA、RNA、蛋白质、代谢物
相互作用
系统
(2)合成生物学的组成工具
器件 device
生物部件 part
系统
system
模块 module
将这些器件逐级设计构建组合成具有特定功 能的生物系统。
标准生物部件
具有特定生物学功能的基因编码元件
启动子、调控因子、核糖体结合位点、编码
细菌QS系统作用
细菌根据特定信号分子的浓度可以监测周围
环境中自身或其它细菌的数量变化,当信号达到
一定的浓度阈值时,能启动菌体中相关基因的表
达来适应环境中的变化。
枯草芽胞杆菌利用QS系统对细胞的发育进行调控
当营养丰富、菌体稀少时向感受态方向发展;
营养贫乏菌体密度高时向芽胞方向发展。
15.4 展望
酵母的木糖代谢工程
运动发酵单胞菌的乙醇代谢工程
大肠杆菌的乙醇代谢工程
EMP
大肠杆菌的乙醇代谢工程
主要优势
大肠杆菌能够利用非常广泛的碳源,其中包 括六碳糖(葡萄糖,果糖)和五碳糖(木糖,阿拉伯 糖 ) 以及糖酸等物质,这一特性使得大肠杆菌能 利用木质纤维素降解产生的各种糖类,同时又由 于大肠杆菌遗传背景清楚,因此在原核微生物乙 醇代谢工程以及木质纤维素的高效利用中具有重 要的研究价值。
菌,使之能将血红素转化为光敏感的藻胆青素
PCB。
PCB PCB
PCB
Black
ompC promoter
lac Z
15.3.2 生命体代谢途径的重新构建
微生物载体生产外源蛋白,目前人类利用
E. coli生产1000多种人类蛋白。
代谢途径改造----调节核心组件优化途径
不同的生物学途径提取出来
优化整合到宿主细胞 合成目标化学物质
LOGO
第15章
合成生物学
内 容
1. 合成生物学的发展历史及概念 2. 研究方式和工具 3. 合成生物学的研究方向
4. 展 望
15.1 合成生物学的发展史及概念
(1)合成生物学的发展史 1978年 Skallka在对限制性内切核酸酶的评论中 第一次预言了合成生物学的诞生。 1980年 Hobom引入了合成生物学的的名词来描述 基因重组技术。 DNA合成测序技术的发展和工程学在生物体系的应 用,为合成生物学奠定基础。
(高达82Kb), 细菌仍保持了良好的生存状态。
改造后菌株的电穿孔效率、基因表达都有改变。
电穿孔
Endy小组用12 kb 人工合成的DNA取代野生T7
基因组中的11 kb 的非必须DNA 构建新的生命体。
最小基因组优点 选择性的保留所需的代谢途径和功能;
成为合成基因网络理想的容器;
为插入模块提高最简单无干扰的环境。
标准化 从可更的换部件库,快速构建多组分体系, 包括建立生物学功能、试验的检测条件及系统做 出等通用、便捷的标准。
不同部件间要进行标准化来实现“即插即用”
的性能。
2003 MIT成立了标准生物部件登记处,数据
库收集了3200个标准化生物学部件。
复杂系统去偶合 将一个复杂的问题分解成若干可操作的独立
胞”,一种称为丝状支原体的细菌。
(2) 合成生物学
合成生物学学是生物科学在二十一世纪刚刚
出现的一个分支学科。
目的在于设计和创造新的生物组件和体系,
对现有的生物体系进行重新设计。从基本的生物
组件构建复杂的人工生命体系,对整个生命过程
进行重新设计、改造、构建。
合成生物包含的内容
基因合成
构建人工生命体
用合成生物学方法创造的微生物进入发酵罐培养生长
2. 青蒿酸合成线路的设计构建
疟疾 典型的疟疾多呈周期性发作,表现为间歇性 寒热发作。发作时先有明显的寒战,全身发抖, 面色苍白,接着体温迅速上升,达40℃或更高, 面色潮红,全身大汗淋漓,大汗后体温降至正常 或正常以下。 经过一段间歇期后,又开始重复上述间歇性 定时寒战、高热发作。
1. 生物质能和乙醇发酵微生物
E. Coli 的乙醇代谢重组菌:
具有五碳糖和六碳糖代谢酶系 混合酸发酵 乙醇耐受能力低
绿色植物和海洋藻类合成的有机物(生物质)约 2200亿吨,相当于人类当前每年全部能耗的10倍。
可用于发酵生产乙醇的部分微生物及其主要底物
酵母或细菌 酵母 酿酒酵母 (S. cervisiae) 卡尔斯伯酵母 (S. carlsbergensi) 鲁氏酵母 (S. riuxii)〔嗜高渗透压) 粟酒裂殖酵母 (S. pombe) 胞壁克鲁维酵母 (K. fragilis) 乳酸克鲁维酵母 (ctis) 嗜单宁管囊酵母 (P. tannophilus) 休哈塔假丝酵母 (C. shehatae) 假热带假丝酵母 (C. pseudotropicalis) 热带假丝酵母(C. tropicalis) 树干毕赤酵母 (P. stipitis) 细菌 运动发酵单胞菌 (Zymomonas mobilis) 热纤维梭菌 (C. Thermocellum) (嗜热) 热硫化氢梭菌 (C. Thermohydrosulfricum) (嗜热) 布氏热厌氧菌(Thermoanaerobium brickii) (嗜热) 乙酞乙基热厌氧杆菌 (Thermobacterium acertoethylic us ) 可发酵的主要底物 葡萄糖、果糖、半乳塘、麦芽糖、麦芽三糖和 木酮糖 葡萄糖、果糖、半乳塘、麦芽糖、麦芽三糖和 木酮糖 葡萄糖、果糖、麦芽糖、蔗糖 葡萄糖、木糖 葡萄糖、半乳糖、乳糖 葡萄糖、半乳糖、乳糖 葡萄糖、木糖 葡萄糖、木糖 葡萄糖、半乳糖、乳糖 葡萄糖、木糖、木酮糖 葡萄糖、木糖 葡萄糖、果糖和蔗糖 葡萄糖、纤维二糖和纤维素 葡萄糖、木糖、蔗糖、纤维二糖淀粉 葡萄糖、蔗糖、纤维二搪 葡萄塘、蔗糖、纤维二糖
基于现有的 天然生物组件, 设计构建有新功 能的生物体系。
Personalized Medicine Physical enhancement: 荷尔蒙,义肢的使用 Enhancing Evolution: 使人聪明的药 Bionic man 生化人
Better, stronger, faster
理想的细胞底盘应具备的条件 ① 长期培养中保持基因稳定 ② 能够在低营养培养基中生长以降低成本
③ 同时协调多基因的表达
④ 能够通过调整合成路径抑制与生产无关的合成路径
15.3.4 构建多细胞体系 多细胞体系是建立在群体细胞效应的研究基 础上,多细胞涉及细胞间的通信体系。
群体效应:微生物通过自身产生的一种化学 信号来感受群体的浓度,从而表现出某种特殊的 行为。
1.人工构建合成生命体 2002年 Wimmer小组脊髓灰质炎病毒的合成
Venter 合成噬菌体基因组和生殖道支原体基因组
不同物种间基因组的移植 将蕈状支原体基因组移植到山羊支原体中。
丝状支原体
酵母载体插入到 细菌基因组中
分离
移植
甲基化
转化酵母
基因组改造
2. 最小基因组的构建
Blattnerj小组删除大肠杆菌基因组的15%
4. 利用合成生物学生产新能源
Kaslling利用13个可逆的酶促反应组合起来创
建一条非天然的催化路径。
淀粉 + 水
H2
15.3.3 最小基因组与合成生物学
合成生物学最终目标:
合成独立的可遗传的人工生命体
人工生命的基本要素
具有膜系统 能进行新陈代谢
具有自己的基因
研究最简化生命的两种方法
1. 从下而上:从核苷酸合成新生命体。 2. 从上而下:从基因组中剔除非必要基因组。
每年5亿人感染,100万死亡。目前最有效的
是青蒿素,生产周期长、成本昂贵。 中药青篙中提取的有过氧基团的倍半萜内酯 药物。
Keasling利用合成生物学,将大肠杆菌改造成
青蒿酸工厂。将甲羟戊酸合成途径转入大肠杆菌
中,改造获的E. coli 青蒿酸的产量300mg/L。