DNA的生物合成和损伤修复DNA Biosynthesis and DNA Damage Repair
DNA的损伤和修复

C 碱基的互变异构
由于碱基氢原子位置的可逆性变化,导致基因发生 酮式-烯醇式或氨式-亚氨式间的结构互变。 导致A-C错配或G-T错配。
3 自由基对DNA的氧化损伤
O2.,OH. ,H2O2等
如:造成DNA链上脱氧戊糖C-3或C-5磷脂键断裂。
自由基还可引起碱基损伤或脱落。
二、物理因素引起的DNA损伤
指DNA分子内发生较大片段的交换,也称为重组。
移位的DNA可以在新位点上颠倒方向反置(倒位), 也可以在染色体之间发生交换重组。
二、突变的意义
1. 突变是进化、分化的分子基础
进化过程是突变的不断发生所造成的。
没有突变就没有今天的五彩缤纷的世界。 遗传学家认为:没有突变就不会有遗传学。
大量的突变都属于由遗传过程自然发生的,
叫自发突变或自然突变(spontaneous mutation)。
•
由于基因突变产生的新性状是生物 从未有过的性状,因此它是
• • •
产生新基因的途径, 生物变异的根本来源, 为生物进化提供了原始的材料。
2. 突变导致基因型改变
突变只改变基因型,而无可察觉的表型改变。
多态性 (polymophism):是用来描述个体之间的基因 型差别现象。利用DNA多态性分析技术,可识别个体差
碱基对组成或排列顺序的改变。
时间:DNA复制时期,即细胞(有丝和减数)分裂间期。
基因的精确复制是相对的。一定的条件下基因结构发生变化变成一个新基因(突变基因)。 于是后代的表现型中也就出现祖先了从未有的新性状
基因突变
插入
┯┯┯┯ ATGC TACG ┷┷┷┷
缺失 替换
┯┯┯┯┯ ATAGC TATCG ┷┷┷┷┷ ┯┯┯ AGC TCG ┷┷┷ ┯┯┯┯ ACGC TGCG ┷┷┷┷
分子生物学 6 DNA 损伤、修复和重组

吖啶橙、原黄素、吖黄素等吖啶类染料 嵌合到DNA碱基对之间 base addition /deletion / frameshift mutation
DNA损伤(DNA damage)
自发损伤: 脱氨基/ 脱嘌呤 外源损伤: 1. 氧化损伤 (需氧细胞) 活性氧:超氧化物,过氧化氢和羟自由基(· OH) 8-氧鸟嘌呤,2-氧腺嘌呤,5-甲酰尿嘧啶 2. 烷基化损伤 影响DNA复制和转录时的解旋 多数是间接诱变 3. 加成损伤 嘧啶二聚体 苯并芘(肝脏细胞色素P-450) 双环氧物-G 芳基化试剂 黄曲霉毒素B1(肝致癌剂)
DNA损伤、修复和重组
突变和突变发生
(mutation and mutagenesis) DNA损伤(DNA damage) DNA修复(DNA repair) 重组(recombination)
突变概念
突变(mutation) DNA分子碱基序列的可遗传改变 突变体(mutant) 与野生型(+)相对 突变剂(mutagen) 突变发生(mutagenesis) 自发突变(spontaneous mutation) 诱发突变(induced mutation)
突变类型 1. DNA碱基序列改变的多少 单点突变(point mutation) 碱基替换(base substitution) 转换(transition) A-T G-C 颠换(transversion)A-T T-A 碱基增加(base addition) 碱基删除(base deletion) 多点突变(multiple mutation)
BER
5' 3' UvrABC 3' 5' 3' 5' Pol I (或δ和ε) 5' 3' DNA glycosylase 5' 3' AP内切核酸酶 5' 3' 进一步酶切
DNA损伤与修复

防止突变和癌症发生
DNA损伤可能导致基因突变, 进而引发癌症
DNA修复机制可以及时修复损 伤,防止突变积累
修复机制的缺陷可能导致突变 增加,增加癌症风险
了解DNA损伤与修复有助于预 防和治疗癌症
适应性进化
DNA损伤与修 复是生物进化 的基础,有助 于物种的适应 性和生存能力。
DNA损伤与修 复机制的突变 和变异是生物 进化的重要驱 动力,有助于 物种的多样性
激光辐射:导致DNA结构变化和 突变
添加标题
添加标题
添加标题
添加标题
电离辐射:直接或间接导致DNA 断裂或交联
极端温度:高温导致DNA变性, 低温引起结晶化
化学因素导致的损伤
烷化剂:导致DNA加合物的形 成
碱基类似物:与DNA碱基配对 错误
甲醛:导致DNA的交联和突变
紫外线:引发嘧啶二聚体的形 成
揭示生物进化历程: DNA损伤与修复的研究 有助于揭示生物的进化历 程,为生物多样性的研究 提供有力支持。
探索生物进化方向:通 过研究DNA损伤与修复 的机制,可以预测生物 未来的进化方向,为生 物多样性的保护提供科 学依据。
05
DNA损伤与修复的挑战和未来发展方向
损伤和修复机制的深入研究
挑战:DNA损伤的类型和复杂 性
的复杂性
挑战:寻找更 有效的修复方
法
未来发展方向: 利用基因编辑 技术治疗遗传
性疾病
未来发展方向: 开发新型药物
和治疗方法
感谢观看
汇报人:XX
阐述如何利用 DNA损伤与修复 机制开发新型抗 癌药物
介绍一些成功的 基于DNA损伤与 修复机制的抗癌 药物案例
分析未来在抗癌 药物研发中DNA 损伤与修复领域 的发展趋势和挑 战
DNA损伤和修复

Chapter 5 DNA damage and repair1.主要内容1)诱变2) DNA损伤3) DNA修复4.教学要求:1)熟悉DNA损伤的原因、类型2)掌握DNA修复的方式5.1Mutagenesis5.2DNA damage5.3DNA Repair5.1 Mutagenesis (诱变)•…5.1.1The reasons of mutation•5.1.2Types of mutations5.1.3Mutagens (诱变剂)5.1.4mutagenesis (诱变)5.1Mutagenesis (诱变)Mutation (突变)=Permanent, heritable alteratioins the base sequence of a DNA molecule(是DNA碱基序列水平上永久性的、可遗传的变化)5.1.1The reasons of mutationSpontaneous errors(自发性错误):in DNA replication or meiotic recombination Mutagen (诱变物): A consequence of the damaging effects of physical or chemical mutagens on DNAMutagen= mutation causing agent• Essentially all mutagens are carcinogens(致癌物)Most carcinogens are mutagens5.1.2Types of mutations• Multisite(多位点):-Cause gross(严重的)chromosome abnormalities(畸形);-Involve large regions of DNA ;-Arise during meiosis(在减数分裂期发生)・Point mutations (点突变)-Involve only one or a few nucleotides ;-Arise during DNA replication-Require two errors :1)An error during DNA replication;2)Failure to correct that errorTypes of point mutationsInversions: ACBDEFDuplications ABCDEEFDeletions: ABCD-FInsertions: ABCDSEFSubstitutions: ATCDEFTypes of point mutationsThe phenotypic effect of point mutation (点突变的表型效应)•Missense mutation(错义突变):碱基序列的改变引起了产物氨基酸序列的改变。
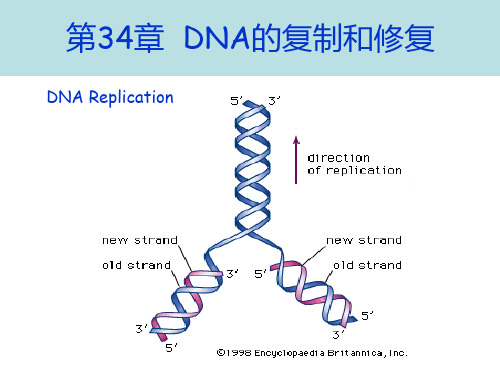
第34章 DNA的复制和修复(2013-2-28)
复制的类型和方式—滚环复制
E. coli phage (噬菌体): ΦX174
• “Template “rolls”, extrudes leading strand • Okazaki frags made on leading strand as it emerges.
复制的类型和方式—多复制叉复制
半保留复制非常普遍
• 单链DNA首先复制合成双链的复制型(单链DNA复 制时,通常先宣传双链复制型,再进行半保留复制)
思考题
1. 设计实验证明DNA是半保留复制
-2012大连理工大学生物化学
2. 猪流感病毒HRN1是反转录病毒,试想出实验方案以阻止病毒在细
胞内复制而不影响细胞内DNA正常复制。 一个同学想研究DNA复制过程用放射性磷来标记底物,他标记的磷是
意义:按半保留复制方式,子代DNA与亲代DNA的碱基 序列一致,子代保留了亲代的全部遗传信息,体现了遗传 的保守性。是物种稳定的分子基础,但是相对的。
possible copying mechanism OF DNA
DNA复制方式有三种可能性,即全保留、半保留和分散式。
弥散式
2. DNA半保留复制的证明(两个)
思考题
3. DNA是以半保留方式进行复制的,如果放射性全标记的双 链DNA分子在无放射性标记的溶液中经两次复制,那么所产 生的4个DNA分子其放射性状况如何?
A、两个分子含有放射性; B、全部含有放射性; C、双链中各有一半含有放射性; D、所有分子的两条链都没有放射性。
华南理工大学2006年生物化学
链的复制起点不在同一点上,一条链先复制,另一条 链保持单链而被取代:当一条链复制到一定程度时才 暴露出另一条链的复制起点,另一条链才开始复制, (单向复制,全连续复制,没有冈崎片段)
第二节 DNA的损伤和修复

Mismatch Repair
5′ ′
3′ ′
(UvrD)
5′ ′
3′ ′
(UvrD)
Mismatch Repair
or X
3. 修复过程
Dam甲基化酶可使GATC序列甲基化。 甲基化酶可使 序列甲基化。 甲基化酶可使 序列甲基化 复制后,新形成的DNA双链 双链GATC序列是半甲基化的,既模板 序列是半甲基化的, 复制后,新形成的 双链 序列是半甲基化的 链甲基化,新合成的链未甲基化。 链甲基化,新合成的链未甲基化。 MutS二聚体可识别新合成链的错配部位。 MutL与MutS结合, 二聚体可识别新合成链的错配部位 结合, 二聚体可识别新合成链的错配部位。 与 结合 复合物可沿DNA链移动,直至 链移动, 序列。 形成的 MutL-MutS复合物可沿 复合物可沿 链移动 直至GATC序列。 序列 由于MutS与错配碱基和 与错配碱基和GATC同时结合,因此 同时结合, 由于 与错配碱基和 同时结合 因此DNA形成一个 形成一个 突环。 序列5′ 开新合成的单链。 突环。MutH在GATC序列 ′端切开新合成的单链。 在 序列 端切开新合成的单链 核酸外切酶 Ⅰ/Ⅹ(3 ′ →5′)或Ⅶ/RecJ( 5′ → 3 ′)切去包括 Ⅹ ′ ( ′ 错配碱基的片断。此过程中, 解螺旋酶Ⅱ 错配碱基的片断。此过程中,由解螺旋酶Ⅱ(UvrD)和SSB ) 帮助解开双链。 帮助解开双链。 DNA pol III 重新合成切除的片断。 重新合成切除的片断。 DNA连接酶连接 DNA。 连接酶连接 连接酶 。
2. SOS反应诱导的修复系统 反应诱导的修复系统 避免差错的修复 (error free repair) ) 错配修复、直接修复、 错配修复、直接修复、 切除修复、 切除修复、重组修复
DNA损伤反应与DNA的修复(一)
DNA损伤反应与DNA的修复(一)DNA损伤的后果很严重,所以生物进化出了各种修复手段,针对不同类型的损伤。
真核生物的DNA修复主要有4种类型:核苷酸切除修复(NER)、碱基切除修复(BER)、错配修复(MMR)和双链断裂修复(DSBR)。
真核生物的四种DNA修复类型。
Prog Neurobiol. 2011NER可切除大片段的DNA损伤,BER可修复个别碱基的损伤,MMR用于修复碱基的错配,而DSBR又包括两种机制:非同源末端连接(NHEJ)和同源重组(HR)。
NHEJ直接连接断端而不需要模板,HR使用完整的姐妹染色单体作为修复模板。
多数DNA修复用的蛋白在平时不会大量表达。
例如很多修复途径需要核酸内切酶,但内切酶同时也可以降解核酸,造成额外的损伤,所以只有在需要时才会大量合成。
其它与修复相关蛋白的表达以及活性也同样需要一个调控机制。
另一方面,修复会涉及DNA合成、染色质重塑等过程,需要足够的核苷酸、还原力、甲基和乙酰基供体等,所以这个调控机制还要负责启动代谢重编程,以提供代谢方面的支持。
DNA损伤修复需要代谢支持。
Front Oncol. 2018这种调控机制由DNA的损伤触发,引起细胞内的一系列相关反应,统称为DNA损伤反应(DNA damage response,DDR)。
DDR识别损伤,然后启动级联反应,根据损伤程度决定细胞命运。
如果损伤严重到难以修复,细胞会进入凋亡甚至坏死;如果损伤轻微并可修复,细胞继续存活并启动修复程序。
DNA损伤反应调控细胞命运。
Cancer Biol Ther. 2015DDR从两个方面对需要修复的细胞进行调整:一方面通过激活DNA损伤检查点阻止细胞进入有丝分裂期,直至修复完成;另一方面则激活并协调各种修复途径,以及诱导代谢重编程。
一般认为,DDR中起主导作用的是三个PI3K相关激酶(PIKK)家族的激酶:DNA依赖性蛋白激酶(DNA-PK)、毛细血管扩张性共济失调症突变激酶(ATM),以及ATM和Rad3相关激酶(ATR)。
DNA损伤与修复
位点,特异的核酸内切酶能识别并切割这样的部 位,造成链断裂。 或损伤碱基被另一种特异的DNA-糖基化酶除去, 形成无嘌呤嘧啶位点(apurinic-apyrimidinic site, AP site),或称无碱基位点(abasic site),这些位点 在内切酶等的作用下可形成链断裂。
第十五章
DNA损伤与修复
DNA Damage and Repair
目录
各种体内外因素所导致的DNA组成与结构的 变化称为DNA损伤(DNA damage)
DNA损伤的后果: 其一,DNA的结构发生永久性改变,即 突变 其二,导致DNA失去作为复制和/或转录 的模板的功能
目录
第一节 DNA损伤
目录
二、DNA损伤有多种类型
碱基损伤与糖基破坏 碱基之间发生错配 DNA单链或双链断裂 DNA链的共价交联
目录
碱基损伤与糖基破坏:
化学毒物可通过对碱基的某些基团进行修饰而改 变碱基的性质。
由于碱基损伤或糖基破坏,在DNA链上可能形成 一些不稳定点,最终可导致DNA链的断裂。
目录
目录
缺失或插入都可导致框移突变 。 框移突变是指三联体密码的阅读方式改变,造
成蛋白质氨基酸排列顺序发生改变。
目录
缺失引起框移突变:
正常 5’ ……G C A G U A C A U G U C ……
丙
缬
组缬
缺失C 5’ ……G A G U A C A U G U C ……
谷
酪
蛋
丝
目录
目录
SOS修复中LexA-RecA操纵子的作用机制
第4章 DNA损伤、修复及重组
• TT 、 CC 、 CT 之间都可 形成二聚体。
• 紫外线引起的DNA损伤最易形成胸腺嘧啶二聚 体(TT)。
(2)电离辐射造成的DNA损伤 • 碱基的变化:细胞中的H2O经辐射分解后产生•OH, 使碱基氧化修饰,形成过氧化物,导致碱基的破 坏和脱落。 • 脱氧核糖的变化:•OH可以使脱氧核糖分解,引 起DNA单链或双链断裂。 • DNA的交联,包括DNA链间交联和DNA与蛋白质 的交联。
• 以胸腺嘧啶二聚体为例,含有二聚体的DNA仍可 进行复制,但复制到二聚体时要暂停一下,然后 越过此处障碍,在二聚体的后面以未知的机制开 始复制,这种起始复制可能不需引发。 • 这样在合成的子链上留下一个大缺口,而其互补 链则复制成完整的双链。然后由完整双链中的母 链与带缺口的子链发生重组。
• DNA合成时,复制叉遇到嘧 啶二聚体,会跳跃过损伤部 位,在下游约1000个核苷酸 处重新开始。复制的子链形 成一个缺口。 • recA蛋白可以识别并结合于 此,并同时识别同源双链区。 • 在recA蛋白的作用下,同源 双链发生重组交换,无损伤 的母链断裂修补缺损的空隙。 • 母链的缺损部位以互补链为 模板,进行合成修复。
(四)烷基转移修复
• 烷化剂所引起的最常见的DNA损伤时使鸟嘌呤O6 甲基化。在大肠杆菌中存在 O6-甲基鸟嘌呤转移酶, 可修复甲基化的碱基。将O6甲基鸟嘌呤的甲基转 移至该酶的一个半胱氨酸上,酶自身失活。
二、切除修复
• 切除修复是指在一系列酶的作用下,将DNA分子 中受损伤部分切除,然后以另一条完整的互补链 为模板,重新合成切去的部分,是DNA恢复正常 结构的过程。
5´ 3´
T
G
GATC
GATC
3´ 5´
CH3
dNTPs
十三章-DNA的生物合成-复制
4
第二节 参与DNA复制的酶
• DNA的复制是一个十分复杂而精确的 过程,涉及多种酶和蛋白因子,如:
DNA聚合酶
DNA连接酶
引物酶
解旋酶 拓扑异构酶 单链结合蛋白等
5
一、DNA聚合酶
• 此酶最早在大肠杆菌中发现,以后陆续在细菌、植物
和哺乳动物中找到。
• 这类酶的共同性质是:
• [1]以脱氧核苷三磷酸(dNTP)为底物合成DNA;
39
40
第四节、DNA的复制过程
• DNA复制的起始 • DNA链的延伸 • DNA复制的终止
41
一、复制的起始
DNA复制的起始包括: 对起点的识别 模板DNA超螺旋及双螺旋的解除 引物的合成等
统称为引发(priming)
42
• 1、辨认起点:复制是从DNA分子上的特定部 位开始的,这一部位叫做复制起始点 (originof replication)常用ori或O表示。 关键序列:两组短的重复序列
聚合酶II 主要作用为修复损伤,该酶的最适
模板是双链DNA而中间有空隙(gap)的单链 DNA部分,而且该单链空隙部分不长于100个 核苷酸。对于较长的单链DNA模板区该酶的 聚合活性很低。无5´→3´外切酶活力,有 3´→5´外切酶活力,但活力低。
16
3、大肠杆菌 DNA Polymerase III • 聚合酶III: • 由10种亚基(αβγδδ′εθτχψ)组 成不对称异源二聚体。 • α、ε、θ亚基组成全酶的核心酶 • γ2、δ、δ’、χ、ψ组成γ-复合物
9
II、3'→5'外切酶活性──校对作用
• 3’-5’外切酶活性与聚合酶紧密结合,当 聚合出现错配碱基时,聚合反应立即停 止,生长链的3’末端核苷酸落入3’-5’外 切酶位点,错配核苷酸被迅速除去,聚 合反应继续进行。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
7.Multi-enzymes and proteins participate in DNA replication. ⑴. Topoisomerases regulate the type and level of supercoiling of dsDNA. ⑵. Helicases unwind the dsDNA. ⑶. SSBs bind and stabilize the single DNA strand. ⑷. Primase synthesizes the RNA primer.
6. DNA replication needs priming. ⑴. DNA polymerases cannot initiate DNA replication by starting a new DNA chain. They can only add nucleotides to the 3’ end of an existent piece of RNA or DNA under the direction of the template. The existent piece of RNA or DNA are called primer.
What was the result ?
There was one DNA band. It had a lower density than 15N DNA because its position was above on that of 15N DNA.
It was 15N/14N hybrid DNA.
2. The point at which separation of the strands and synthesis of new DNA takes place is known as the replication fork. The replication fork is Y-shaped. Two arms (V) are separated strands which act as the template and DNA synthesis is actively taking place. The body (I) is the parental DNA.
These short fragments are called Okazaki fragments. They are joined by DNA ligase and form the lagging strand.
5. Origins contain short AT-rich repeat sequences. ⑴. Prokaryotic and eukaryotic origins have common features: a. They consist of multiple unique short repeat sequences. b. These sequences are recognition and binding sites of multi-subunit initiation factors.
3.DNA replication is usually bidirectional. ⑴.Replicon: Any piece which replicates as a single unit is called a replicon. All bacterial chromosomes and many phage and virus DNA molecules are circular and comprise single replicons.
After another generation growing in the 14NH4Cl medium the bacterial DNA density was determined.
There were two DNA bands.
One half of the DNA was 14N DNA, and another half was hybrid DNA. In succeeding generations the ratio of 14N DNA to hybrid DNA increased gradually.
⑵. The leading strand and all Okazaki fragments are primed by synthesis of a short piece of RNA ( an RNA primer ), which is then elongated with DNA by DNA polymerase. ⑶. There are also DNA priming or nucleotide priming.
⑸. DNA polymerases elongate DNA chains. ⑹. DNA ligase joins Okazaki fragments.
8. DNA replication is of high fidelity. ⑴. There are two types of replication errors. a. base ( nucleotide ) substitution. b. nucleotide insertion or deletion. ⑵. There are two types of error controls a. presynthetic error control. b. proofreading control
The hybrid DNA became less and less..
summary
DNA replicates in a semiconservative manner. When the two parental strands separate, each serves as the template for making a new , complementary strand.
4. DNA replication is semidiscontinuous. ⑴.The mechanism of DNA replication allows only for synthesis in a 5’3’ direction. ⑵. The two strands of DNA are antiparallel.
⑶.Terminus: In a circular replicon there is a single termination site roughly 180° opposite the unique origin.
Summary In a circular replicon replication begins from the fixed origin and forms two replication forks. The two replication forks proceed bidirectionally away from the origin and the strands are copied as they separate until the terminus is reached.
The density of DNA was increased by labeling it with 15N, a heavy isotope of nitrogen.
This was done by growing E coli 15 generations in a medium that contained 15NH4Cl as its only nitrogen source.
2. 3.
⑴. Watson and Crick predicted that DNA might semiconservatively replicate.
4.
5.
the hypotபைடு நூலகம்esis ___________ p ___________ d ___________ p ___________ p
c. These sequences are usually AT- rich. ⑵. E coli ‘s origin is called oriC. It is 254bp long and contains three 13-bp direct repeats and four 9-bp inverted repeats.
⑶. mismatch repair.
____________ d ____________ p
⑵. In 1958 Meselson and Stahl demonstrated the semiconservative nature of DNA replication in E coli.
The experiment:
CsCl equilibrium gradient density ultracentrifugation of 15N labeled E coli DNA.
Question How is the parental strand that runs 5’3’ past the replication fork copied ? The answer is semidiscontinuous replication.
At each replication fork one strand ( the leading strand ), whose template runs 3’5’ past the replication fork, is synthesized as one continuous piece, while the other strand ( the lagging strand ), whose template runs 5’3’ past the replication fork is made discontinuously as short fragments in the reverse direction.