DNA的生物合成
dna生物合成法

dna生物合成法DNA生物合成法是一种基因工程技术,通过人工合成DNA序列,使其具备特定的功能。
它在生物医学、农业和工业领域有着广泛的应用。
本文将从DNA生物合成法的原理、应用以及未来发展等方面进行介绍。
DNA生物合成法基于DNA的化学合成原理,通过化学合成方法合成具有特定序列的DNA。
DNA合成可分为两种主要方法:固相合成和液相合成。
固相合成是将DNA序列逐个碱基单位地合成在固相载体上,然后逐个碱基单位地进行去保护和连接,最终得到完整的DNA序列。
液相合成是将DNA序列逐个碱基单位地合成在液相中,并通过反应条件的调控来实现碱基的合成和连接。
DNA生物合成法在生物医学领域有着重要的应用。
通过人工合成的DNA序列,可以构建特定的基因和基因组,用于研究基因功能、疾病机制以及药物研发。
例如,科学家可以通过合成DNA序列来研究某种基因在细胞生长和分化过程中的作用,从而揭示其调控机制。
此外,DNA生物合成法还可以用于合成人工基因组,用于构建合成生物和人工细胞等研究。
在农业领域,DNA生物合成法也有着广泛的应用。
通过合成DNA 序列,可以改良作物的性状和产量,提高作物的抗病性和适应性。
例如,科学家可以通过合成DNA序列来改良作物的免疫系统,使其对病原体具有更强的抵抗力。
此外,DNA生物合成法还可以用于合成转基因作物,使其具备特定的抗虫性或耐草甘膦等特性。
在工业领域,DNA生物合成法也有着重要的应用。
通过合成DNA 序列,可以构建具有特定功能的酶和代谢途径,用于生物催化合成和生物能源转化等领域。
例如,科学家可以通过合成DNA序列来构建高效的酶催化系统,用于生物催化合成有机化合物。
此外,DNA生物合成法还可以用于合成生物能源,如合成生物柴油和生物氢等。
DNA生物合成法在未来还有着广阔的发展前景。
随着合成生物学和基因工程技术的不断发展,合成DNA序列的合成效率和质量将得到进一步提高。
这将为生物医学、农业和工业领域的研究提供更多的选择和可能性。
DNA的生物合成(精)

一. DNA的复制
复制部位:
真核生物:细胞核
原核生物:细胞质的核质区
(一) 复制的反应
一. DNA的复制
n1d ATP n2d CTP n3d GTP n4d TTP
DNA聚合酶 DNA模板
DNA +(n1+n2+n3+n4)PPi
PPi随即被焦磷酸酶水解,从 而推动聚合反应的进行。
做半保留复制(semiconservative replication)。
(二) 复制的方式 半保留复制
一. DNA的复制
(二) 复制的方式
一. DNA的复制
如何证明半保留复制
1958年,Meselson 证明:用,15NH4Cl唯一氮源
培养大肠杆菌,之后,用14NH4Cl培养,然后进行
CsCl2进行密度梯度离心。由于15NH4Cl密度大于
双螺旋DNA
3′5′ 带切开的3′ 端单链穿越 与另一条连 接封口 Tyr
一.DNA的复制
TopⅠ被解离 (-) (-)
P OH
2个负超螺旋 DNA-酶中间物
O R HN CH C NH R′ CH 2 Tyrosine N O O O 5′ H Oˉ H P O O P Oˉ (b) O O H H DNA链 N H N NH 2 N
② 随后链的合成
引物的合成:随后链的每个冈崎片段都需要合成
RNA引物。也是由引物酶催化。
冈崎片段的合成: DNA聚合酶 Ⅲ (原核细胞 )在引物的 3'末端使DNA链延伸,直至抵达其 下游的另一个冈崎片段的 RNA引物
的5'端。
(五)复制的过程 3.复制叉的推进-复制叉推进的过程
11.DNA的生物合成
The model of DNA-pol III
11/62
DNA pol I
H B
J
小片段 大片段 (Klenow fragment)
5 ‘ →3 ’聚合功能, 3 ' →5 '外切酶活性 5 ' →3 '外切酶活性
12/62
真核细胞中DNA聚合酶
种类: DNA-pol α、β、γ、δ、ε… DNA pol δ:合成领头链
UGA(终止密码子):Trp AGA/AGG(Arg):终止密码子 AUA(Ile):Met(起始密码子)
14/62
3’
5’ 5’ 3’
OH
P
DNA-pol
5’
3’ 5’
3’
DNA-pol 的 5´3´聚合作用
15/62
外切酶与内切酶作用图解
内切酶 (限制性内切酶) 5´ 3´外切 5’ 3´5´外切 3’
4. 冈崎片段(Okazaki fragment):
不连续复制的片段
38/62
ori 5. 双向复制 以起始点为中 心,向两个方 向进行复制。
6. 复制子(replicon) 真核生物两个相 邻复制起始点之 间的DNA片段。 ori
ori
ori
39/62
滚环复制
是某些病毒,质粒、线粒体 DNA的特殊复制形式。
性质
Ⅲ 20 100000 有 无 有 复制
10/62
Leading strand synthesis
Lagging strand synthesis
’
form the catalytic core
生物化学重点_第十一章dna的生物合成

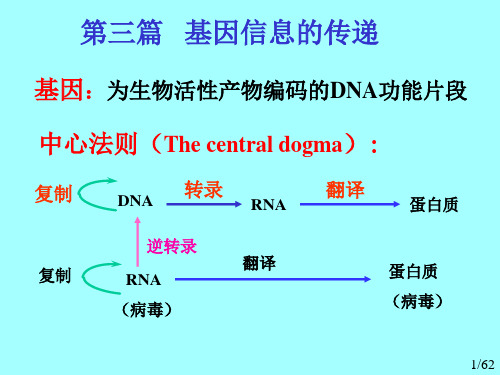
生物化学重点_第十一章D N A的生物合成work Information Technology Company.2020YEAR第十一章 DNA的生物合成一、中心法则:① DNA的自我复制将遗传信息由亲代传递给子代;② 转录:以DNA为模板合成RNA;③ 翻译:mRNA指导蛋白质的生物合成,从而决定生物的表现型。
DNA的复制、转录和翻译过程就构成了遗传学的中心法则。
但在少数RNA病毒中,其遗传信息贮存在RNA中。
因此,在这些生物体中遗传信息的流向是④ RNA通过复制,将遗传信息由亲代传递给子代;⑤ 通过反转录将遗传信息传递给DNA,再由DNA通过转录和翻译传递给蛋白质,二、DNA复制的特点:1.半保留复制:DNA在复制时,以亲代DNA的每一股作模板,合成完全相同的两个双链子代DNA,每个子代DNA中都含有一股亲代DNA链,这种现象称为DNA的半保留复制(semiconservative replication)。
DNA以半保留方式进行复制,是在1958年由M. Meselson 和 F. Stahl 所完成的实验所证明。
2.需要引物(primer):DNA聚合酶必须以一段具有3'端自由羟基(3'-OH)的RNA作为引物,才能开始聚合子代DNA链。
3.半不连续复制:由于DNA聚合酶只能以5'→3'方向聚合子代DNA链,因此两条亲代DNA链作为模板聚合子代DNA链时的方式是不同的。
以3'→5'方向的亲代DNA链作模板的子代链在聚合时基本上是连续进行的,这一条链被称为前导链(leading strand)。
而以5'→3'方向的亲代DNA链为模板的子代链在聚合时则是不连续的,这条链被称为随后链(lagging strand)。
DNA在复制时,由随后链所形成的一些子代DNA短链称为冈崎片段(Okazaki fragment)。
三、DNA复制的条件:1.底物:以四种脱氧核糖核酸(deoxynucleotide triphosphate)为底物,即dATP,dGTP,dCTP,dTTP。
第13章 DNA生物合成(简明生物化学)

Dna A辨认复制启始点,然后引物酶进入(DnaG 蛋白) ,加上解螺旋酶、 DnaB蛋白和DnaC蛋 白等,与DNA的起始复制区域形成引发体。
DNA聚合酶Ⅲ 由其β亚单位辨认引物,新链的 第一个脱氧核苷酸与引物的3-OH形成磷酸二酯键, 开始复制
滚动环式:单向复制,低等生物如质粒 共价闭环双链分子的正链由核酸内切酶在一特
定位点切开,游离出的5’-磷酸基末端固定在细胞膜 上,然后以环状负链为模板,从正链的3’-OH末端 延长形成正链。不需要另外合成引物。
3′ 5′
5′
3′
领头链
5′Leabharlann 5′ 随从链3′ 3′
5′
(二)引发体的生成
复制过程需要引物--短链RNA
拓扑异构酶 单链结合蛋白 解链酶 引物酶及引发体 DNA聚合酶 DNA连接酶 引物
冈崎片段
领头链 3′ 5′
随从链 3′
5′
五、 DNA连接酶(ligase)
• 催化两段DNA之间的连接
′
5P
3′ OH
+ 5′ P
γ
P O-
β
O PO Oα-
3′
OH
DNA
ligase +AMP
5′ P
PPi
O 3′ OH
一种是全部轻的14N-14N。为1∶1; 3代:仍有两种分子,但14N-14N增多,为
1∶3; 4代:两者比为1∶7。
DNA半保留复制的证据
细菌 (含15N-DNA)
普通培养基
第一代
普通培养基
普通DNA
普通DNA 重DNA
第二代
重DNA
DNA的生物合成

参与 参与DNA复制的物质 参与 复制的物质
底物(substrate): dATP, dGTP, dCTP, dTTP 底物 聚合酶(polymerase): 依赖DNA的DNA聚合酶,简写 聚合酶 依赖 的 聚合酶, 聚合酶 为 DNA-pol 模板(template) : 解开成单链的 解开成单链的DNA母链 模板 母链 引物(primer): 提供 ′-OH末端使 提供3′ 末端使dNTP可以依次聚合 引物 末端使 可以依次聚合 其他的酶和蛋白质因子
A G G T A C T G C C A C T G G
T C C A T G A C G G T G A C C
C C A C T G G
G G T G A C C
AT GC GC TA AT CG TA GC CG CG AT CG TA GC GC
+
AT GC GC TA AT CG TA GC CG CG AT CG TA GC GC
一、半保留复制的实验依据和意义
半保留复制的概念 半保留复制的概念
DNA生物合成时 , 母链 生物合成时, 母链DNA解开为两 生物合成时 解开为两 股单链,各自作为模板(template)按碱基配对 股单链,各自作为模板 按碱基配对 规律,合成与模板互补的子链。 规律 , 合成与模板互补的子链 。 子代细胞的 DNA,一股单链从亲代完整地接受过来,另 ,一股单链从亲代完整地接受过来, 一股单链则完全从新合成。 一股单链则完全从新合成 。 两个子细胞的 DNA都和亲代 都和亲代DNA碱基序列一致。这种复制 碱基序列一致。 都和亲代 碱基序列一致 半保留复制。 方式称为半保留复制 方式称为半保留复制。
DNA的生物合成

13/16.DNA的生物合成 13.2 原核生物DNA的复制 13.2.1 参与原核DNA复制的酶和蛋白质
1.原核生物的DNA聚合酶 (1)DNA聚合酶Ⅰ:
Klenow片段,含DNA聚合 酶和3´→5´核酸外切酶活性
13/16.DNA的生物合成
13.2 原核生物DNA的复制
13.2.1 参与原核DNA复制的酶和蛋白质
• 基因组能独立进行复制的单位称为复制子,每个复制子都含有控制复制起始
的起点,可能还有终止复制的终点
• 大多数原核生物染色体DNA的复制是双向,形成复制眼,单向复制的特殊形
式,称为滚动环式
• 真核生物染色体DNA是线形双链分子,含有许多复制起点,因此是多复制子。
13/16.DNA的生物合成 13.1 DNA复制的概况 13.1.2 DNA复制的起点和方向
13/16.DNA的生物合成 13.2 原核生物DNA的复制 13.2.1 参与原核DNA复制的酶和蛋白质
1.原核生物的DNA聚合酶
β-滑动夹子 将正在复制的DNA固定在夹子中心,并能随DNA复制沿着模板DNA链滑动 使DNA聚合酶不易从模板脱离,有利于DNA的连续复制
13/16.DNA的生物合成 13.2 原核生物DNA的复制 13.2.1 参与原核DNA复制的酶和蛋白质
2.参与原核生物DNA复制的其他酶和蛋白质 (5)其它蛋白因子
单链结合蛋白(SSB-single-strand binding protein) 稳定已被解开的DNA单链,阻止复性和保护单链不被核酸酶降解。
引发前体 它由多种蛋白质dnaA、dnaB、dnaC、n、n´、n´´ 和i组成。引发前体再与引发
若双链DNA中一条链有切口,一端是3´-OH,另一端是5´-磷酸基,连接酶可 催化这两端形成磷酸二酯键,而使切口连接。
生化-第十章DNA的生物合成

3. 大肠杆菌 大肠杆菌DNA聚合酶 Ⅲ——polⅢ 聚合酶 Ⅲ DNA复制酶,1972年发现 复制酶, 复制酶 年发现 是真正起复制作用的酶, (1)pol Ⅲ 是真正起复制作用的酶,由10种 ) 种 亚基组成不对称二聚体 不对称二聚体, 、 、 组成核心酶 亚基组成不对称二聚体,α、ε、θ组成核心酶 (2)功能: )功能: 聚合酶活性; ① 5′→3′聚合酶活性; 聚合酶活性 外切酶活性。 ② 3′→5′外切酶活性。 外切酶活性 该酶在原核细胞中主要负责DNA链的延伸, 链的延伸, 该酶在原核细胞中主要负责 链的延伸 是复制延长中真正起催化作用的酶。 是复制延长中真正起催化作用的酶。
双向复制
复制叉
起点 单向复制 起点
的复制--( 三、原核细胞DNA的复制--( 原核细胞 的复制--(DNA指导下的 指导下的 DNA合成) 合成) 合成 (一)DNA聚合酶 聚合酶 1956年kornberg等首先从大肠杆菌中发现 年 等首先从大肠杆菌中发现DNA 等首先从大肠杆菌中发现 聚合酶。其后在广泛不同的生物中都找到有这 聚合酶。 种酶。 种酶。
加入的dNTP 加入的
亲核攻击
5′
引 物
3′
DNA模板链 模板链 脱氧核糖
底物: 底物: dNTP (dATP dGTP dCTP dTTP); ; 聚合酶( 聚合酶(polymerase, DNA-pol): , 依赖DNA的DNA聚合酶 是1种模板指导的酶 聚合酶,是 种 依赖 的 聚合酶 模板( 解开成单链的DNA母链; 母链; 模板(template): 解开成单链的 母链 引物( 提供3′-OH末端 使dNTP聚合; 末端,使 聚合; 引物(primer): 提供 末端 聚合 其它酶和蛋白质因子
Arthur Kornberg won the 1959 Nobel Prize in Medicine for his discovery of the mechanism in the biological synthesis of deoxyribonucleic acid (before Watson and Crick won theirs!)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
母链DNA
复制过程中形成 的复制叉
子代DNA
目录
• 密度梯度实验
梯度离心结果
含重氮N15-DNA的细菌
培养于普 通培养液
第一代
继续培养于 普通培养液
第二代 ——实验结果支持半保留复制的设想。
目录
• 半保留复制的意义
按半保留复制方式,子代DNA与亲代DNA
的碱基序列一致,即子代保留了亲代的全部遗
传信息,体现了遗传的保守性。 遗传的保守性,是物种稳定性的分子基础, 但不是绝对的。
目录
(二)真核生物的DNA聚合酶
DNA-pol DNA-pol DNA-pol DNA-pol DNA-pol 起始引发,有引物酶活性。 参与低保真度的复制 。 在线粒体DNA复制中起催化作用。 延长子链的主要酶,有解螺旋酶活性。 在复制过程中起校读、修复和填补缺 口的作用。
目录
5' 3'
dCTP
dGTP
dTTP
dATP
dCTP
dATP
dGTP
dTTP
目录
领头链的合成
目录
随从链的合成
目录
目录
目录
目录
目录
目录
(三)复制的终止
• 原核生物基因是环状DNA,双向复制的复制 片段在复制的终止点(ter)处汇合。
ori
0
ori
50
82
32
ter
SV40
ter
E.coli
全称:依赖DNA的DNA聚合酶 (DNAdependent DNA polymerase) 简称:DNA-pol
活性:1. 5'→3' 的聚合酶活性
2. 核酸外切酶活性
目录
目录
• 核酸外切酶活性
5´
AG C T T C A G G A T A
3´
| | | | | | | | | | |
3´
?
目录
前导链是连续合成,而后续链是不连续合成。 DNA的合成始于RNA引物的合成,因此前导 链只有一次RNA合成,滞后链的冈崎片断每 个都有RNA引物的合成。RNA引物随后由 DNA片段替换,相邻的DNA片段连接形成一 条完整的DNA链。 DNA聚合酶的校对功能和细胞的错误修复功 能维持DNA分子的准确性和忠实性。
(250kD)
功能 是原核生物复制延长中真正起催化作用的酶。
目录
目录
α亚基具有5′→3′方向合成DNA的催化活性 ε亚基具有3′→5′外切酶活性,起校对功能, 可提高聚合酶Ⅲ复制DNA的保真性 亚基可能起组建的作用 β亚基犹如夹子,功能是识别引物、夹住DNA 分子向前滑动 亚基起着促使核心酶二聚化的作用 其余的亚基构成 -复合物,主要功能是帮助 亚基夹住DNA(也称夹子装置器),并有 增强核心酶活性的作用
目录
5’ 3’
ori
ori
ori
ori
3’ 5’
5’ 3’
3’
5’
5’
3’
3’
5’
复制子
目录
(2)起始点和方向 ①原核生物和真核生物复制都是在DNA分子特定的 位置起始的,如大肠杆菌起始点有固定的重复的保守顺
序,可被酶识别。
②方向:大多是双向、对称复制, 也有单向或不对称 的。 ③速度: 原核生物复制叉移动快(约105bp/min); 真核生物复制叉移动慢 (约5×102~5×103bp/min)
目录
目录
目录
引发体和引物
Dna B、 Dna C Dna A 5 3 DNA拓扑异构酶 SSB 5 3
含有解螺旋酶、DnaC蛋白、引物酶和DNA 复制起始区域的复合结构称为引发体。
目录
3
5 3
引物
5
引物是由引物酶催化合成的短链RNA分子。
目录
目录
DNA复制的调控是在起始阶段进行的, 一旦复制起始,它就会继续下去直到整个复制子 完成复制。 复制起点是以一条链为模板起始DNA 合成的一段序列。有时,两条链的复制起点并不 在同一点上(不对称复制,如D环复制)。
目录
5’
O
3’
3’
O P OO-
HO
ATP
5’
DNA连接酶
ADP 5’ 3’ 3’
O O P OO-
5’
目录
目录
•功能
• DNA 连接酶在复制中起最后接合缺口的
作用。
• 在 DNA修复、重组及剪接中也起缝合缺 口作用。 • 也是基因工程的重要工具酶之一。
目录
一、原核生物的DNA生物合成
(一)复制的起始
目录
• 解螺旋酶(helicase) ——利用ATP供能,作用于氢键,使DNA双链 解开成为两条单链 • 引物酶(primase) ——复制起始时催化生成RNA引物的酶 • 单链DNA结合蛋白(single stranded DNA binding protein, SSB) ——在复制中维持模板处于单链状态并保护单 链的完整
目录
复制起始点、复制子与复制叉(动 画演示)
目录
三、复制的半不连续性
3
5 3
领头链 (leading strand)
解链方向
随从链 (lagging strand)
5
目录
• 顺着解链方向生成的子链,复制是连续进行的,
这股链称为领头链(前导链)。
• 另一股链因为复制的方向与解链方向相反,不能 顺着解链方向连续延长,这股不连续复制的链称 为随从链(后随链、滞后链)。复制中的不连续 片段称为岡崎片段(okazaki fragment)。
引物(primer): 提供3’-OH末端使dNTP可以依次聚合 其他的酶和蛋白质因子
目录
一、复制的化学反应
(dNMP)n + dNTP → (dNMP)n+1 + PPi
目录
• 聚合反应的特点
除了底物和酶外,DNA 新链生成需引
物和模板;
新链的延长只可沿5’→ 3’方向进行 。
目录
二、DNA聚合酶
目录
细菌复制终止区含有多个约22bp的终止子 (terminator)位点,E. coli 有7个终止子位点。
目录
新合成的两条 染色体DNA分子相互 连在一起,被拓扑 异构酶IV分离
目录
DNA复制的分子机制的基本特点
复制的结果是半保留复制,复制的过程是 半不连续复制。 复制以复制单位进行,起始于特定的位点 (复制起点)。 复制可以是单向,也可以是双向,后者更 为常见。 复制时,DNA的两条链都从5′端向3′端延 伸。
子代DNA
目录
第一节
复制的基本规律
Basic Rules of DNA Replication
目录
一、半保留复制的实验依据和意义
•半保留复制的概念
DNA 生物合成时,母链 DNA 解开为两 股单链,各自作为模板(template)按碱基配对 规律,合成与模板互补的子链。子代细胞的 DNA,一股单链从亲代完整地接受过来,另 一股单链则完全从新合成 。两个子细胞的 DNA都和亲代DNA碱基序列一致。这种复制 方式称为半保留复制。
5´
T C G AA G T C C T A G C G A C
• 3'→5'外切酶活性 能辨认错配的碱基对,并将其水解。 • 5'→3' 外切酶活性 能切除突变的 DNA片段。
目录
目录
目录
目录
目录
目录
目录
功能:
① 5′→3′聚合酶活性; 酶与模板结合后构象改变,识别碱基,正 确配对后才发挥聚合作用 ② 3′→ 5′外切酶活性; 主要是对新生DNA链进行校对,防止“错 配” ③ 5′→3′外切酶活性。 主要是对DNA损伤的修复,以及在DNA复 制时RNA引物的切除及其空隙的填补
在一个完整的细胞周期中,每一个复 制起点只使用一次,完成一次复制过程。 多数生物的复制起点,都是富含A.T。
目录
(二)复制的延长
复制的延长指在 DNA-pol 催化下, dNTP 以 dNMP的方式逐个加入引物或延长中的子链上, 其化学本质是磷酸二酯键的不断生成。
目录
3' 5'
DNA-pol
OH 3'
可见,DNA polⅠ是1个多功能酶
目录
N端
DNA-pol Ⅰ 木瓜蛋白酶
C端
小片段
323个氨基酸 5'→3'核酸外切酶活性
大片段/Klenow 片段
604个氨基酸 DNA聚合酶活性 3'→5'核酸外切酶活性 • Klenow片段是实验室合成DNA,进行 分子生物学研究中常用的工具酶。
目录
DNA-pol Ⅲ
需要解决两个问题: 1. DNA解开成单链,提供模板。
2. 合成引物,提供3-OH末端。
目录
原核生物E.coli为例
复制由起始点 oriC(origin)开始双向复制
E.coli的复制起始点oriC(245bp的DNA组分) 序列分析有如下特点 3组串连重复序列(13bp),每个顺序都由GATC开始 2组反向重复序列(9bp) 4个dnaA蛋白结合部位
目录
二、双向复制
原核生物复制时,DNA从起始点(origin)向 两个方向解链,形成两个延伸方向相反的复制 叉,称为双向复制。
复制中的放射自显影图象
目录
Cairns复制模型—θ型复制
目录
目录
ori
ter
A
B
C
A. 环状双链DNA及复制起始点 B. 复制中的两个复制叉
C. 复制接近终止点(termination, ter)