邓嘉祺 2013031104分子生物学
班级成绩明细表

2024年度-朱玉贤现代分子生物学第四版

蛋白质翻译后加工的意义
对于蛋白质的成熟、定位和功能发挥具有重要作用。例如,信号肽的去除可以使蛋白质从细胞内分泌 到细胞外或定位到细胞膜上;化学修饰可以调控蛋白质的活性和稳定性,从而影响细胞的生理功能; 剪切可以产生具有不同功能的蛋白质片段,增加蛋白质的多样性。
17
转录与转录后加工的调控
转录的调控主要通过转录 因子与DNA的结合来实 现,可以影响RNA聚合酶 的活性和选择性。
转录和转录后加工的调控 具有协同作用,可以共同 调节基因的表达水平和蛋 白质的功能。
ABCD
转录后加工的调控涉及多 种蛋白质和RNA的相互作 用,可以影响RNA的加工 效率和产物种类。
29
基因工程与基因组学的应用前景
农牧业领域
通过基因工程改良作物和畜禽品种, 提高产量和品质,增强抗逆性;应用 基因组学解析重要农艺性状形成的分 子机制,指导新品种选育。
工业领域
利用基因工程生产工业酶、生物燃料 和生物材料等;应用基因组学优化工 业生产过程和开发新产品。
医学领域
基因工程可用于生产重组蛋白药物、 基因诊断和基因治疗等;基因组学可 用于解析人类疾病的遗传基础,发现 新的治疗靶点和药物。
异常的转录和转录后加工 调控可能导致疾病的发生 ,如癌症、遗传性疾病等 。
18
05
蛋白质翻译与翻译后加工
19
蛋白质翻译的过程与特点
蛋白质翻译的过程
起始、延长和终止三个阶段。起始阶段,核糖体与mRNA结合,形成起始复合物;延长阶段,tRNA携带氨基酸 进入核糖体,进行肽链的延伸;终止阶段,释放完成翻译的蛋白质。
2018 年生命科学学院全日制硕士研究生复试名单 - SCNU
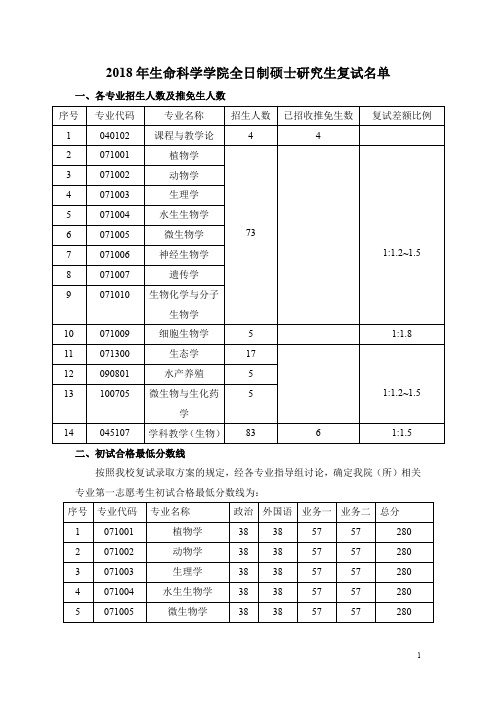
2018年生命科学学院全日制硕士研究生复试名单
一、各专业招生人数及推免生人数
二、初试合格最低分数线
按照我校复试录取方案的规定,经各专业指导组讨论,确定我院(所)相关专业第一志愿考生初试合格最低分数线为:
三、复试名单
(第一志愿)
按照差额比例,确定第一志愿考生复试名单如下(按初试成绩由高到低排列):
(调剂考生)
按照我校调剂办法,经考生申请,我院(所)相关专业指导组审查和筛选,根据专业培养要求,按照1:1.2-1.5的差额比例,确定调剂考生复试名单如下(因调剂生流动性较大,最后实际参加复试的名单以研招网调剂系统上的为准)
生命科学学院
2018年3 月23日。
种子活力或抗老化能力的分子机制研究进展

种质资源(germplasm resources)是生物遗传信息的载体,是现代种业的核心竞争力和农业科技原始创新的物质基础,是保障国家粮食、生态及能源安全的重要战略性资源[1]。
为避免种质资源的丢失,有必要对其安全保存[2]。
种子是种质资源保存的重要载体,但生理成熟后的种子在贮藏过程中质量会不可逆转地逐渐下降,这种变化称为老化或劣变(aging or deterioration)。
种子因老化而面对外界胁迫挑战的脆弱性增加,种子活力降低[3]。
种子老化程度因贮藏条件不当特别是高温高湿条件而不断加重[4~5];在此期间,种子内部会发生一系列有害变化,例如种皮机械耐性下降[6]、细胞膜损伤、蛋白质变性、DNA 损伤和突变、核酸合成系统破坏等[7~9],最终导致种子活力的丧失。
如果种子在贮藏过程中老化或活力下降的问题得不到解决,就会造成种子萌发迟缓、生长势差、抗逆性弱、生物产量和经济产量降低,从而影响农业生产。
近年来,随着分子生物学研究技术的发展,特别种子活力或抗老化能力的分子机制研究进展姜孝成*,周诗琪(湖南师范大学生命科学学院作物不育资源创新与利用湖南省重点实验室,中国湖南长沙410081)摘要:种子代表高等植物生活史中的一个重要时期,也是种质多样性资源保存的重要载体。
种子在贮藏过程中的老化是不可避免的,了解种子活力或抗老化能力的分子机制对于农业生产具有重要意义。
本文对数量性状位点分子标记技术、全基因组关联分析、组学技术和基因工程等在种子活力或抗老化能力的分子机制方面的研究进展进行了综述,以期为通过分子设计育种技术改良种子的活力或抗老化能力提供理论指导。
关键词:种子老化;数量性状位点(OTL);全基因组;组学;基因工程中图分类号:Q945;Q756文献标识码:A文章编号:1007-7847(2021)05-0406-11收稿日期:2021-08-12;修回日期:2021-09-28基金项目:国家自然科学基金资助项目(32072125,31671773);湖南省生物发育工程及新产品研发协同创新中心项目(20134486)作者简介:*通信作者:姜孝成(1964─),男,湖南新化人,湖南师范大学教授,博士生导师,主要从事种子安全与分子生物学研究,E-mail:***************.cn 。
红外图像的目标检测研究

红外图像的目标检测研究
余祉祺
【期刊名称】《数字通信世界》
【年(卷),期】2024()1
【摘要】红外搜索与跟踪系统被广泛应用于现代国防中针对远距离目标的检测、跟踪及预警。
但是,该统检测的目标一般为尺寸较小且模糊的点状结构通常难以辨别,一种基于图像中目标与背景局部结构的差异性,采用NRAM模型实现的方法可以对单帧红外图像中的小目标进行有效检测。
【总页数】4页(P34-37)
【作者】余祉祺
【作者单位】香港大学
【正文语种】中文
【中图分类】TN215
【相关文献】
1.基于可见光图像和红外图像决策级融合的目标检测算法
2.基于红外图像与可见光图像特征融合的目标检测算法
3.融合红外特征的可见光图像目标检测算法研究
4.单帧红外图像弱小目标检测研究综述
因版权原因,仅展示原文概要,查看原文内容请购买。
MAPK信号途径与自噬在甘蔗鞭黑粉菌有性配合与菌丝生长过程中的功能研究

作者签名:
日期:
导师签名:
日期:
学位论文提交同意书
本学位论文符合国家和华南农业大学关于研究生学位论文的相关 规定,达到学位授予要求,同意提交。
导师签名:
日期:
学科带头人签名:
日期:
摘要
甘 蔗 鞭 黑 穗 病 ( Sugarcane smut ) 是 一 种 由 孢 堆 黑 粉 菌 属 的 甘 蔗 鞭 黑 粉 菌 (Sporisorium scitamineum)引起的重要病害,对我国乃至世界的糖产业均是一个巨大 的威胁。研究病原菌的分子生物学特性可为明确致病机制、制定防控策略等提供理论 依据,但对于甘蔗鞭黑粉菌迄今人们还知之甚少,亟待加强研究。为给甘蔗鞭黑粉菌 分子生物学特性增加资料,本论文对甘蔗鞭黑粉菌自噬基因 SsATG8 和 MAPK 信号途 径关键基因 SsKPP2 进行了近三年的研究,获得了如下主要结果:
1. We identified the S. scitamineum MAP kinase Kpp2 by using the Candida albicans Cek1 protein sequence, named the coding gene as SsKPP2. Alignment of the SsKpp2 with it’s ortholous proteins demonstrated that there is an invariant TXY motif among the MAP kinases, which has been shown to be phosphorylated upon stimulation through its MAPK signaling cascade.
3. We identified the S. scitamineum ATG8 gene, named SsATG8, by TBLASTN using the S. cerevisiae Atg8 protein sequence as a bait. Alignment of the SsAtg8 protein with its othorlogs in other smut fungi and model organisms showed a high degree of conservation in their amino acid sequence, including the conserved C-terminal glycine residue subject to post-translational modification, which is essential for autophagy induction.
华中师范大学核物质科学协同创新中心2019年硕士研究生招生拟录取名单公示

单位名称
姓名
033 核物质科学协同创新中心 段文烽
考生编号
录取专 业码
录取专业
105119112303503 070200 物理学
033 核物质科学协同创新中心 方涛
105119112410697 070200 物理学
033 核物质科学协同创新中心 周文艺 105119112410698 070200 物理学
04 嵌入式系统应用
全日制 非定向 328 69.3 68.56
备注
非定向
366 75.2 74
理
02
粒子物理与原子核物 全日制
非定向
297 67.72 62.73
理
02
粒子物理与原子核物 全日制
非定向
367 83.08 77.27
理
02
粒子物理与原子核物 全日制
非定向
331 69.88 67.67
理
02
粒子物理与原子核物 全日制
非定向
352 81.24 74.74
研究方向
01 理论物理
录取学习 录取类
复试
总分
总成绩
方式 别
成绩
全日制 非定向 308 80.48 69.15
01 理论物理
全日制 非定向 330 74.68 69.47
01 理论物理
全日制 非定向 298 80.6 68
01 理论物理
全日制 非定向 326 71.96 67.9
01 理论物理
全日制 非定向 359 74.64 72.94
02
粒子物理与原子核物 全日制
非定向
391 88.8 82.44
理
02
2019云南大学发展研究院硕士研究生拟录取名单
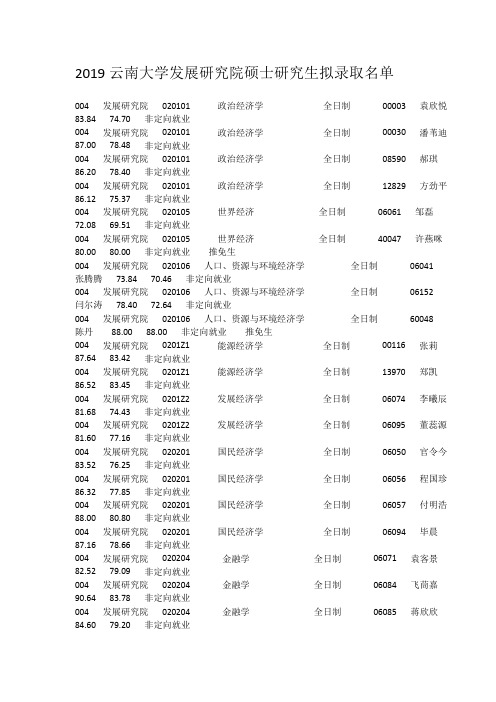
2019云南大学发展研究院硕士研究生拟录取名单004 发展研究院020101 政治经济学全日制00003 袁欣悦83.84 74.70 非定向就业004 发展研究院020101 政治经济学全日制00030 潘苇迪87.00 78.48 非定向就业004 发展研究院020101 政治经济学全日制08590 郝琪86.20 78.40 非定向就业004 发展研究院020101 政治经济学全日制12829 方劲平86.12 75.37 非定向就业004 发展研究院020105 世界经济全日制06061 邹磊72.08 69.51 非定向就业004 发展研究院020105 世界经济全日制40047 许燕咪80.00 80.00 非定向就业推免生004 发展研究院020106 人口、资源与环境经济学全日制06041张腾腾73.84 70.46 非定向就业004 发展研究院020106 人口、资源与环境经济学全日制06152闫尔涛78.40 72.64 非定向就业004 发展研究院020106 人口、资源与环境经济学全日制60048陈丹88.00 88.00 非定向就业推免生004 发展研究院0201Z1 能源经济学全日制00116 张莉87.64 83.42 非定向就业004 发展研究院0201Z1 能源经济学全日制13970 郑凯86.52 83.45 非定向就业004 发展研究院0201Z2 发展经济学全日制06074 李曦辰81.68 74.43 非定向就业004 发展研究院0201Z2 发展经济学全日制06095 董蕊源81.60 77.16 非定向就业004 发展研究院020201 国民经济学全日制06050 官令今83.52 76.25 非定向就业004 发展研究院020201 国民经济学全日制06056 程国珍86.32 77.85 非定向就业004 发展研究院020201 国民经济学全日制06057 付明浩88.00 80.80 非定向就业004 发展研究院020201 国民经济学全日制06094 毕晨87.16 78.66 非定向就业004 发展研究院020204 金融学全日制06071 袁客景82.52 79.09 非定向就业004 发展研究院020204 金融学全日制06084 飞茼嘉90.64 83.78 非定向就业004 发展研究院020204 金融学全日制06085 蒋欣欣84.60 79.20 非定向就业004 发展研究院020204 金融学全日制06105 邓攀86.28 81.43 非定向就业004 发展研究院020204 金融学全日制30049 杨友琴81.00 81.00 非定向就业推免生第20 页,共97 页2019(按照学院代码+专业代码+考生编号后五位排序)学院考生编号学院名称专业代码专业名称培养方式姓名复试成绩综合成绩拟录取类别备注代码后五位004 发展研究院020204 金融学全日制30050 潘韵迪85.00 85.00 非定向就业推免生004 发展研究院020204 金融学全日制30051 鲁子琪85.00 85.00 非定向就业推免生004 发展研究院020205 产业经济学全日制06116 司莹86.72 76.93 非定向就业004 发展研究院020205 产业经济学全日制06117 王琳82.80 78.24 非定向就业004 发展研究院020205 产业经济学全日制06119 唐洁薇89.84 78.90 非定向就业004 发展研究院020205 产业经济学全日制06133 张菊招79.08 73.51 非定向就业004 发展研究院020205 产业经济学全日制06134 邵雪88.88 78.75 非定向就业004 发展研究院020205 产业经济学全日制06163 梁德飞84.08 77.55 非定向就业004 发展研究院020205 产业经济学全日制06164 蒲云洋78.88 74.39 非定向就业004 发展研究院020205 产业经济学全日制10052 耿梦佳80.00 80.00 非定向就业推免生004 发展研究院020207 劳动经济学全日制06114 徐婧80.52 73.01 非定向就业004 发展研究院020207 劳动经济学全日制06177 曹雨轩87.80 78.20 非定向就业004 发展研究院020207 劳动经济学全日制06188 李凯80.76 73.94 非定向就业004 发展研究院025100 金融全日制06194 唐伟杰83.60 82.64 非定向就业004 发展研究院025100 金融全日制06196 马义波81.64 72.98 非定向就业004 发展研究院025100 金融全日制06204 李馨79.88 73.23 非定向就业004 发展研究院025100 金融全日制06212 赖星宇77.20 71.68 非定向就业004 发展研究院025100 金融全日制06213 李玲娜81.24 72.70 非定向就业004 发展研究院025100 金融全日制06223 孔雪梅80.56 73.74 非定向就业004 发展研究院025100 金融全日制06228 李奇采83.40 73.92 非定向就业004 发展研究院025100 金融全日制06233 李晨瑜84.72 75.77 非定向就业004 发展研究院025100 金融全日制06234 钱见超74.08 71.27 非定向就业004 发展研究院025100 金融全日制06236 刘庆喜79.92 74.45 非定向就业004 发展研究院025100 金融全日制06241 曹颖77.68 73.19 非定向就业004 发展研究院025100 金融全日制06246 刘亦珩85.48 75.59 非定向就业004 发展研究院025100 金融全日制06256 王洋洋82.76 77.98 非定向就业004 发展研究院025100 金融全日制06259 陈嘉辉78.72 75.05 非定向就业004 发展研究院025100 金融全日制06260 曾子钰81.48 73.39 非定向就业004 发展研究院025100 金融全日制06262 赵文华86.04 75.46 非定向就业004 发展研究院025100 金融全日制06265 莫崇昆80.88 73.15 非定向就业004 发展研究院025100 金融全日制06269 张悦84.96 76.10 非定向就业004 发展研究院025100 金融全日制06274 郝倩倩81.04 74.42 非定向就业004 发展研究院025100 金融全日制06275 刘于波84.76 79.62 非定向就业004 发展研究院025100 金融全日制06280 肖洋72.28 71.27 非定向就业004 发展研究院025100 金融全日制06291 黄美娇83.48 76.11 非定向就业004 发展研究院025100 金融全日制06300 孔德玮76.60 72.88 非定向就业004 发展研究院025100 金融全日制06301 黎东79.68 73.63 非定向就业004 发展研究院025100 金融全日制06302 宋燕云80.7676.94 非定向就业004 发展研究院025100 金融全日制10054 徐鑫鑫85.00 85.00 非定向就业推免生第21 页,共97 页2019(按照学院代码+专业代码+考生编号后五位排序)学院考生编号学院名称专业代码专业名称培养方式姓名复试成绩综合成绩拟录取类别备注代码后五位004 发展研究院025100 金融全日制90053 朱峰琳70.00 70.00 非定向就业推免生004 发展研究院025600 资产评估全日制00245 黄子建88.48 78.95 非定向就业004 发展研究院025600 资产评估全日制00337 肖媛玲82.40 74.96 非定向就业004 发展研究院025600 资产评估全日制01651 石育铭86.40 76.80 非定向就业004 发展研究院025600 资产评估全日制02402 戴玮琪86.80 81.76 非定向就业004 发展研究院025600 资产评估全日制04050 张伦伟84.40 74.44 非定向就业004 发展研究院025600 资产评估全日制04255 张洪铭81.92 73.33 非定向就业004 发展研究院025600 资产评估全日制06155 白林鹭84.48 75.55 非定向就业004 发展研究院025600 资产评估全日制06322 钱锦78.36 74.90 非定向就业004 发展研究院025600 资产评估全日制06332 戴睿宸84.96 74.42 非定向就业004 发展研究院025600 资产评估全日制08129 潘胜贵78.12 71.81 非定向就业004 发展研究院025600 资产评估全日制09835 张天奇82.28 74.07 非定向就业004 发展研究院025600 资产评估全日制12513 严鹏86.44 79.46 非定向就业004 发展研究院025600 资产评估全日制12532 高雁泽88.24 77.66 非定向就业004 发展研究院025600 资产评估全日制12541 范志伟82.72 74.37 非定向就业004 发展研究院025600 资产评估全日制13204 袁青绵85.12 77.37 非定向就业004 发展研究院025600 资产评估全日制13243 王皓月82.36 75.66 非定向就业004 发展研究院025600 资产评估全日制13933 王杉81.80 74.48 非定向就业004 发展研究院025600 资产评估全日制19197 周悦84.36 74.78 非定向就业004 发展研究院025600 资产评估全日制20425 曾静逸72.60 69.84 非定向就业004 发展研究院025600 资产评估全日制22930 吴虹霖79.60 74.92 非定向就业004 发展研究院025600 资产评估全日制27799 董雨菲87.68 81.75 非定向就业004 发展研究院025600 资产评估全日制60127 熊清雨77.20 73.48 非定向就业004 发展研究院030302 人口学全日制06345 黄旭东72.60 70.80 非定向就业004 发展研究院030302 人口学全日制06346 李忠骥84.40 76.84 非定向就业004 发展研究院085239 项目管理全日制36279 杨威85.00 73.84 非定向就业004 发展研究院120100 管理科学与工程全日制06368 李鸿浩82.00 74.80 非定向就业004 发展研究院120100 管理科学与工程全日制06381 杨理想83.60 76.64 非定向就业004 发展研究院120100 管理科学与工程全日制20509 刘迁迁79.60 77.80 非定向就业004 发展研究院120100 管理科学与工程全日制40056 李仕灵79.00 79.00 非定向就业推免生004 发展研究院120100 管理科学与工程全日制60055 王月84.00 84.00 非定向就业推免生004 发展研究院120100 管理科学与工程全日制60057 孙绍林83.00 83.00 非定向就业推免生004 发展研究院120204 技术经济及管理全日制03696 张凡82.40 74.00 非定向就业004 发展研究院120204 技术经济及管理全日制20052 陈予涵86.40 78.72 非定向就业004 发展研究院120204 技术经济及管理全日制22960 孙煜88.60 77.44 非定向就业004 发展研究院125600 工程管理非全日制06413 许永太84.90 66.16 非定向就业004 发展研究院125600 工程管理非全日制06416 张逸伦81.06 69.82 非定向就业004 发展研究院125600 工程管理非全日制06424 毕天祥82.20 71.28 定向就业84.56 76.62 定向就业第22 页,共97 页2019(按照学院代码+专业代码+考生编号后五位排序)学院考生编号学院名称专业代码专业名称培养方式姓名复试成绩综合成绩拟录取类别备注代码后五位004 发展研究院125600 工程管理非全日制06434 李良84.98 73.59 非定向就业004 发展研究院125600 工程管理非全日制06437 何芳76.70 65.68 非定向就业004 发展研究院125600 工程管理非全日制06442 牛丁81.26 70.30 非定向就业004 发展研究院125600 工程管理非全日制06443 马杰79.90 64.56 非定向就业004 发展研究院125600 工程管理非全日制06445 李丹83.00 69.60 非定向就业004 发展研究院125600 工程管理非全日制06447 王蕊84.98 70.99 非定向就业004 发展研究院125600 工程管理非全日制06449 江珂86.00 69.20 非定向就业004 发展研究院125600 工程管理非全日制06451 袁利阳81.00 70.00 非定向就业004 发展研究院125600 工程管理非全日制06457 段志钢79.54 69.02 定向就业004 发展研究院125600 工程管理非全日制06459 李秀笑92.00 76.00 定向就业004 发展研究院125600 工程管理非全日制06460 唐倩84.84 73.34 非定向就业004 发展研究院125600 工程管理非全日制06461 陈逵林78.30 64.92 非定向就业004 发展研究院125600 工程管理非全日制06463 陈永炜73.60 65.64 非定向就业004 发展研究院125600 工程管理非全日制06464 唐春艳77.50 66.20 非定向就业004 发展研究院125600 工程管理非全日制06470 王赐秋89.20 75.68 非定向就业004 发展研究院125600 工程管理非全日制06482 陈俊潇90.20 71.28 定向就业004 发展研究院125600 工程管理非全日制06486 杨紫彦82.50 66.00 非定向就业84.90 70.16 定向就业004 发展研究院125600 工程管理非全日制06489 杨霄85.90 70.96 非定向就业004 发展研究院125600 工程管理非全日制06507 陈舒静88.40 68.56 非定向就业004 发展研究院125600 工程管理非全日制06513 张梁87.20 71.88 非定向就业004 发展研究院125600 工程管理非全日制06517 杨凡83.10 69.84 非定向就业004 发展研究院125600 工程管理非全日制06522 许四宇83.32 72.33 非定向就业004 发展研究院125600 工程管理非全日制06528 王世全87.16 72.26 非定向就业004 发展研究院125600 工程管理非全日制06536 李子甘82.44 73.18 非定向就业004 发展研究院125600 工程管理非全日制06539 陈敏瑞78.44 68.78 非定向就业004 发展研究院125600 工程管理非全日制06545 王翔75.70 64.88 非定向就业004 发展研究院125600 工程管理非全日制06547 夏宇83.90 71.56 非定向就业004 发展研究院125600 工程管理非全日制06557 吴儒封79.70 67.28 定向就业004 发展研究院125600 工程管理非全日制06563 宁明84.20 73.68 非定向就业004 发展研究院125600 工程管理非全日制06567 周禹辰78.00 64.80 非定向就业004 发展研究院125600 工程管理非全日制06572 刘瑞格85.20 76.88 非定向就业004 发展研究院125600 工程管理非全日制06577 黄芸77.90 66.96 非定向就业004 发展研究院125600 工程管理非全日制06587 张菲菲81.30 69.92 非定向就业004 发展研究院125600 工程管理非全日制06588 李凌84.80 66.12 非定向就业004 发展研究院125600 工程管理非全日制06589 陈琳86.10 76.64 非定向就业004 发展研究院125600 工程管理非全日制06593 冯光勤80.42 70.77 非定向就业004 发展研究院125600 工程管理非全日制06598 朱鹏举76.30 66.32 定向就业004 发展研究院125600 工程管理非全日制06607 谢依霖88.72 77.69 非定向就业第23 页,共97 页2019(按照学院代码+专业代码+考生编号后五位排序)学院考生编号学院名称专业代码专业名称培养方式姓名复试成绩综合成绩拟录取类别备注代码后五位004 发展研究院125600 工程管理非全日制06614 薛峰78.80 64.72 非定向就业004 发展研究院125600 工程管理非全日制06626 文兴宇81.30 64.52 非定向就业004 发展研究院125600 工程管理非全日制06637 徐梦蕾86.70 68.08 非定向就业004 发展研究院125600 工程管理非全日制06639 蒋洪祥88.20 67.88 非定向就业004 发展研究院125600 工程管理非全日制06647 赵世杰86.00 72.60 非定向就业004 发展研究院125600 工程管理非全日制06685 穆胜才82.00 69.40 非定向就业004 发展研究院125600 工程管理非全日制06691 张志阳80.00 69.40 非定向就业004 发展研究院125600 工程管理非全日制06700 晏平76.20 66.08 非定向就业004 发展研究院125600 工程管理非全日制06702 骆雪姣77.40 64.76 非定向就业004 发展研究院125600 工程管理非全日制06705 刘金璨71.00 64.60 非定向就业004 发展研究院125600 工程管理非全日制06708 徐作华85.10 71.24 非定向就业004 发展研究院125600 工程管理非全日制06709 龙银辉86.30 70.52 非定向就业。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
DNA切除修复途径托马斯·林达尔,彼得·卡伦和理查德·伍德碱基切除的DNA切除修复途径主要是为修复内源性DNA损伤和核苷酸切除,为了修复紫外线造成的DNA损伤,重建了用纯化的哺乳动物蛋白和这些修复机制的细节不断出现。
类似的数据变得可用关于错配修复作为校正复制错误的回应。
小鼠个体DNA修复蛋白基因的缺失提供了关于这些因素在体内和最近三维几种复酶结构中的角色的信息来解释它们作用的详细模式。
地址帝国癌症研究基金会,克莱尔馆实验室,布兰奇巷,南米姆斯,赫特福德郡EN63LD,英国当前观点遗传学与发展1997,7:158-169电子标识:0959-437X-007-00158©目前生物有限公司ISSN0959-437X缩写AAAF N-乙酰乙酰氨基芴BER 碱基切除修复CAF1 染色质组装因子1CS 科凯恩氏综合征8-oxo-G 8-羟基HNPCC 遗传性非息肉性大肠癌NER 核苷酸切除修复PCNA 增殖细胞核抗原UDG 尿嘧啶-DNA糖基UGI UDG抑制剂UV 紫外线XP 着色性干皮介绍DNA分子的完整性病变由三种因素,主要有內源和外缘DNA损伤剂和DNA复制错误。
切除修复三个主要的策略,彼此明显不同,被细胞用于纠正这样的改变。
由于生命的起源,细胞的基因组的主要威胁显然已经存在;高效的DNA修复过程,因此发展比较早,并以相同的潜能继续担任整个进化过程。
事实上,编码基因修复酶每条通路的特点被发现在几个不相关的细菌最近测序的小基因组。
哺乳动物细胞不具有DNA 修复过程,与那些存在的微生物根本不同,尽管反应在多细胞生物的广泛DNA损伤包括p53基因表达上和触发细胞凋亡。
细胞或有机体似乎没有针对损害电离辐射和环境化学诱变剂,但要纠正他们使用的这种病变,不同的成就可以造成一些特定的DNA修复机制,修复功能主要用于校正内源性DNA损伤和遭受紫外线(UV)光DNA损伤。
微生物具有诱导某些修复功能,以抵消诱变和有毒物质的影响的能力:这反映它们的存在在敌对环境的适应和这种诱导通常是不存在于哺乳动物细胞中。
此观点是三个不同的切除修复途径的现有知识的描述,在过去一年中重要的发现。
碱基切除修复D NA和胞嘧啶和5-甲基胞嘧啶残基脱氨的自发水解脱嘌呤,多个反应与羟基自由基生成的副产物的正常氧化代谢和形成,因为细胞内暴露于反应性小的代谢物和辅酶共价关于DNA复合物,生成多种需要精确和快速修复DNA的损伤。
用于校正这种DNA损伤的主要策略是碱基切除修复(BER),其中改变的DNA碱基被游离形式DNA糖基化酶切除,所得脱碱基位点是由AP核酸内切酶、DNA聚合酶校正和DNA连接酶的一致行动。
在1995年初两组[1,2]通过求解尿嘧啶DNA糖基化酶(UDG)提供了一种DNA糖基化酶的第一结构,这是需要从DNA除去脱氨基胞嘧啶残基的高分辨率结构。
所述酶具有DNA结合槽和相邻的口袋紧密配合一个从受损的DNA翻转出的脱氧尿苷残基。
现在相同的两个组已经扩展这项工作UDG的8kDa特异性蛋白抑制剂的结构,称为UGI化酶(UDG抑制剂),它是由枯草芽孢杆菌感染噬菌体PBS1宿主后立即诱导的。
PBS1在其基因组中尿嘧啶代替胸腺嘧啶,这使得它必须关掉宿主UDG功能,PBS1基因组的大量富含AU的DNA序列表示中,未校正的胞嘧啶脱氨在进化过程中已经发生与生存能力兼容最大程度。
有趣的是,UGI蛋白结合非常紧密地与酶的DNA结合槽通过一系列带有极性残基的相互作用。
因而UGI蛋白模仿DNA底物,并采用DNA结合残基与酶的相互作用[1,2]。
抑制剂完全阻止尿嘧啶的接近。
与在UDG 高度保守的残基的相互作用UGI可从许多来源解释,包括细菌,单纯疱疹,和哺乳动物细胞,但不是其它DNA糖基化酶抑制UDG的现象。
UDG的脱氧尿苷结合的结构的精确的学问已经允许构建改变的酶由特异性位点诱变。
通过这种方式; UDG衍生物已经作出将从正常的碱基中切除胞嘧啶或胸腺嘧啶,未损坏的DNA以缓慢的速度[3“]在大肠杆菌中是细胞毒性改变酶的表达;一个致突变作用被认为在表达低水平,因为据推测正常的嘧啶被连续切除。
第二个UDG已在大肠杆菌和哺乳动物细胞[ 4 ]“发现。
这种酶,表达在一个较低的水平,著名的UNG基因产物,可能作为一个内生水解去除DNA损伤的主要酶的备份。
类似的情况,两个DNA糖基化酶识别同一基板,已知的大肠杆菌3 -甲基腺嘌呤DNA糖苷酶——标签和碱性祛痘产品-一个为DNA糖基化酶切除氧化嘧啶残基对,核酸内切酶III 和核酸内切酶VII 1。
在哺乳动物细胞中,小UDG还具有可切除胸腺嘧啶残基在GT碱基的功能。
原来酶被加里纳利和Jiricny发现[ 4 ],由于这个发现,可能有修复脱氨基5-甲基胞嘧啶残基的功能。
大肠杆菌的诱导alkA的基因产物- 和哺乳动物3-甲基腺嘌呤DNA 糖基- 共享了能够切除几种不同类型受损的DNA碱基,除了3-甲基腺嘌呤。
作为酶的良好底物的统一的功能似乎涉及在损坏的基础上的正电荷的存在。
UDG的“策略”,其中有一个贴身合适的缺口,只能容纳一个脱氧尿苷残留,但不正常或损坏的脱氧核苷残基,显然是不适用的。
两组[ 5,6 ]现在已经解决了AIK蛋白质的三维结构。
酶采用一个大的疏水裂富电子芳香族氨基酸残基,而不是缺口容纳翻转损坏残留,作用于多种病变的AIK,然而与UDG酶的周转次数相比相当低。
几种DNA糖基化酶的高分辨率结构,连同突变分析和新兴的数据在酶-底物复合物的三维结构,提供这类修复酶的详细机理的信息。
一个优秀的研究模式[ 7 ]是建立一个新型的螺旋发夹螺旋基序和[4FE-4S]簇环结构与DNA结合的结构在大肠杆菌核酸内切酶III,酶切除胸腺嘧啶乙二醇,胞嘧啶水合物,和环嘧啶DNA碎片。
初始步骤扩大这种研究真核细胞的酶参与最近的酒精酵母和裂殖酵母同源的核酸内切酶III [8,9]表达和克隆。
同样,对大肠杆菌FPG酶切除诱变DNA损伤为8-羟基鸟嘌呤的同系物(8-oxo-g)已被确认在酵母和酿酒酵母的基因已被克隆和表达[10,11]。
对应人类和小鼠cDNA的克隆是在所有实验室的进步,应该允许提高在哺乳动物系统中活性氧的诱变效应得理解。
尝试让小鼠缺乏不同种的酶,在BER是活跃导致早期胚胎致死表型的几例;在过去的一年中,这样的数据已报告AP核酸内切酶,DNA聚合酶β,和XRCC1蛋白。
这种情况说明在胚胎发育过程中的内源性基因修复的相对重要性,并提供了一种解释,显然没有特定缺陷和误码率形式的人类易发癌症疾病。
纯合子突变小鼠细胞在 3 -甲基腺嘌呤DNA糖苷酶的缺陷已经被孙氏集团所[ 12 ]”构建。
这些细胞正常生长,但就像是简单的烷化剂如对甲基敏感的大肠杆菌突变体标签和生物碱,包括专门生产3-甲基腺嘌呤DNA代理。
这种烷基化病变的细胞毒作用可能是通过在模板的小槽中的甲基基团的存在下,通过抑制核酸复制或转录所造成的。
令人惊讶的是,3 -甲基腺嘌呤DNA糖苷酶缺陷细胞对丝裂霉素C过敏。
不知道DNA修复酶是否可以抵消DNA 交联生产的抗癌药物,或是否一个有毒的丝裂霉素C的单加成物去除突变小鼠细胞缺陷。
误码率的过程通常包括一个正常的残基与脱氧核糖核酸的替换。
这个过程现在已经用纯化的重组人蛋白[ 13 ]重组了。
用这种方式,尿嘧啶鸟嘌呤碱基可以有效地纠正了胞嘧啶鸟嘌呤配对。
该反应需要UDG,AP核酸内切酶,DNA聚合酶β,和连接提供最有效的DNA连接酶III / XRCC1基因的异源二聚体的功能。
小鼠纤维细胞细胞系的构建与纯合性缺失的DNA聚合酶[ 3 [ 14 ]提供了很好的证据表明酶的功能特别是误码率。
因此,聚合酶β缺陷细胞正常生长,但简单的烷化剂和细胞提取浓缩物显示用尿嘧啶的DNA底物执行BER的受损能力。
这些缺陷可以被修复,用野生型聚合酶β小转基因稳定转染。
聚合酶β的作用是双重的:修复反应的酶催化的一个核苷酸的间隙填充DNA反应中间物,但小的氨基末端基本域的聚合酶β也是通过13消除过程°[ 15 ]切除的5’-端脱氧核糖磷酸残切脱碱基位点。
除了这种主要形式的误码率,与修复过程的后期阶段不同的第二个次要途径已经确定在哺乳动物细胞中[ 16 ]。
一个明显的反应特征是修补几个核苷酸长的发生,而不只是一个单一的核苷酸,并在复制因子PCNA依赖(图1)。
此外,反应显示对5’核酸酶脱氧和杨核酸酶IV / FEN1有绝对依赖性,有效改变碱基位点的电离辐射是不适合采用脱氧核糖聚合酶活性骨消除β修复产生的修正(一klungland,T林达尔,未发表的数据)。
PCNA反应的影响可以引起DNA缺口由高分子量的DNA聚合酶填充,如δ或ε,结合抗原,或用DNase IV /FEN1 [ 17 ]°在5’端末悬垂结构切除受损的残留。
图1基础切除修复模型,类似于建议在[ 66 ]。
该通路可能被作为分支,不同斑块的大小产生不同的DNA聚合酶进行时。
改变基渣是由DNA糖基化酶和脱碱基位点释放然后被AP核酸内切酶。
通常,DNA聚合酶~填补一个核苷酸和脱氧核糖磷酸残基游离切除。
DNA 连接酶III / XRCC1的异源二聚体进而完成反应。
XRCC1蛋白对DNA聚合酶和DNA连接酶~ 1不同的结合位点。
在另一种路径依赖复制因子的BER过程完成,DNA聚合酶添加几个与亲本DNA链置换核苷酸残基。
“过剩”是由结构特异性核酸酶的DNA酶IV / FEN1修复切除的DNA连接完成。
核苷酸切除修复真核生物NER反应机理的核心核苷酸切除修复(NER)在真核生物中可以起不同效率的作用,在各种各样的DNA加合物中,是最有效的螺旋扭曲病变。
人类的主要功能是去除由于暴露于阳光造成的紫外线诱发的DNA光化细胞。
继1995真核生物NER的纯化组分的重建,阐明了其反应机理的重要细节。
至少,NER双切口反应需要的因素XPA,XPC,RPA,TFIIH,XPG,和ercc1-xpf在哺乳动物细胞中,或在酵母同源因素[ 18,19 ]。
XPA和RPA 彼此结合,优先结合到损伤的DNA。
其中包括其亚基TFIIH XPB和XPD的DNA解旋酶[ 20 ]。
XPG和ERCC1 - XPF因素结构特异性DNA核酸酶和负责裂解在3’和5’侧分别病变,释放伤害一个核苷酸长24-32的残留(图2)。
编码的核心因素XPF基因克隆最近[ 21,22 ]°。
XPF 形成紧密的复杂与ERCC1蛋白,直接与相应的RAD1 RAD 10复合在酵母中。
无论是RAD 1-RAD 10和ERCC1 - XPF配合物具有结构特异性核酸酶活性和贴近单链和双链DNA的单链的边界时,有一个极性5至3远离边境。
这些核酸酶需要ATP和Mg2+,在模型表面不需要其他的结构特异性裂解活性蛋白因子。