BIOENVIRONMENTAL POLYMER SOCIETY
微塑料对双壳贝类毒性效应的研究进展

Advances in Environmental Protection 环境保护前沿, 2022, 12(3), 543-553Published Online June 2022 in Hans. /journal/aephttps:///10.12677/aep.2022.123072微塑料对双壳贝类毒性效应的研究进展李佳,吴为,卫洲,张万*安徽理工大学,安徽淮南收稿日期:2022年5月11日;录用日期:2022年6月15日;发布日期:2022年6月22日摘要微塑料因比表面积大、传输距离长、吸附性强、难降解等特性而广泛存在于水环境系统中,其造成的污染问题已经引起了世界各地学者的广泛关注。
本文以双壳贝类为主要对象,综述了微塑料对水生生物造成影响的途径和海洋中微塑料引起双壳贝类产生的各种毒性效应。
今后,应进一步加强微塑料对双壳贝类毒性效应的研究,建立以双壳贝类作为水环境中微塑料污染的指示生物,形成一套系统的微塑料生物监测方法。
关键词微塑料,水生生物,双壳贝类,毒性效应Research Progress in the Study ofMicroplastics on Toxic Effects on BivalveMollusksJia Li, Wei Wu, Zhou Wei, Wan ZhangAnhui University of Science and Technology, Huainan AnhuiReceived: May 11th, 2022; accepted: Jun. 15th, 2022; published: Jun. 22nd, 2022AbstractMicroplastics are widely present in the water environment system because of their large specific surface area, long transport distance, strong adsorption, and difficulty in degradation. The pollu-tion problems caused by them have attracted extensive attention from scholars all over the world.In this paper, bivalve shellfish as the main object, the influence of microplastics on aquatic organ-*通讯作者。
LetPub-JCR2012-期刊

0239-7528 1851-2372 1224-2780 2146-3123 1860-5397 1876-2883 1757-6180 1369-703X 1934-8630 0791-7945 1996-3599 0324-1130 0525-1931 0144-8617 0008-6223 1897-5593 1664-3828 1634-0744 1842-4090 2044-4753 0920-5861 1933-6918 2045-3701 1478-811X 2228-5806 2211-1247 0008-8846 1895-1066 1733-7178 2081-9900 1868-4904 0008-8994 0272-8842 0862-5468 0173-9913 1584-8663 0352-9568 1612-1872 1752-153X 1359-7345 0275-7540 0098-6445 1385-8947 0009-2347 0255-2701 0360-7275 0263-8762 0009-2509 0930-7516 0009-2460 0009-3122 1451-9372 0009-3068 0251-0790
B POL ACAD SCI-TECH B SOC ARGENT BOT BALK J GEOM APPL BALK MED J BEILSTEIN J ORG CHEM BENEF MICROBES BIOANALYSIS BIOCHEM ENG J BIOINTERPHASES BIOL ENVIRON BUILD SIMUL-CHINA BULG CHEM COMMUN BUNSEKI KAGAKU CARBOHYD POLYM CARBON CARDIOL J CARDIORENAL MED CARNETS GEOL CARPATH J EARTH ENV CATAL SCI TECHNOL CATAL TODAY CELL ADHES MIGR CELL BIOSCI CELL COMMUN SIGNAL CELL J CELL REP CEMENT CONCRETE RES CENT EUR J CHEM CENT EUR J ENERG MAT CENT EUR J GEOSCI CENT EUR NEUROSURG CENTAURUS CERAM INT CERAM-SILIKATY CFI-CERAM FORUM INT CHALCOGENIDE LETT CHEM BIOCHEM ENG Q CHEM BIODIVERS CHEM CENT J CHEM COMMUN CHEM ECOL CHEM ENG COMMUN CHEM ENG J CHEM ENG NEWS CHEM ENG PROCESS CHEM ENG PROG CHEM ENG RES DES CHEM ENG SCI CHEM ENG TECHNOL CHEM ENG-NEW YORK CHEM HETEROCYCL COM+ CHEM IND CHEM ENG Q CHEM IND-LONDON CHEM J CHINESE U
胞外聚合物在环境工程中的研究进展

胞外聚合物在环境工程中的研究进展摘要:胞外聚合物(EPS)具有易于生物降解、高效、无毒、无二次污染等优点,被认为是传统化学聚合物的潜在替代品。
近年来,胞外聚合物在水处理、化工冶炼等方面的应用越来越受到人们的重视。
在这种背景下,关于EPS的文献信息分布广泛,非常稀少。
因此,本文主要分析胞外聚合物在环境工程中的研究。
关键词:聚合物;污水处理;污泥特性;应用前景引言胞外聚合物(EPS)是广泛存在于活性污泥絮体、生物膜和颗粒污泥等微生物聚集体细胞外的聚合物,主要由微生物在一定条件下释放的多糖、蛋白质、核酸、腐殖质等高分子物质聚合而成。
它不仅能形成一个缓冲层为微生物创造稳定的生存环境,还能在细胞缺乏营养物质时,充当底物供细胞存活。
尽管采用活性污泥法处理污水已经超过百年,但是针对作为活性污泥重要组成部分的EPS的机理研究还不透彻。
笔者通过综述近年来EPS在水处理领域的研究进展,讨论了EPS在生物膜和颗粒污泥形成中的作用,以及对污水生物处理的影响机制和EPS调控方面还需解决的问题,以期为今后的相关研究和应用提供参考。
1、EPS在脱氮除磷中的作用强化生物除磷以其经济、可持续等优点被广泛应用。
通常认为,强化生物除磷是由于聚磷菌在厌氧释放磷酸盐,好氧过量吸收磷酸盐,并以聚磷酸盐的形式储存在聚磷菌中,从而达到除磷效果。
然而近年来的研究表明,胞外聚合物(EPS)也在这个过程中起作用。
不同的EPS提取方法检测活性污泥及颗粒污泥EPS中磷含量,发现颗粒污泥EPS中磷含量比活性污泥EPS中磷含量高,污泥中EPS总含量越高,其在磷积累中所占的比例也就越大(EPS中磷占到了颗粒污泥磷积累的45.4%)。
表明磷的积累在强化生物除磷的EPS中是不可忽略的。
EPS中的磷主要包括正磷酸盐(orthophosphate,简称Ortho-P)、焦磷酸盐(pyrophosphate,简称Pyro-P)和聚磷酸盐(polyphosphate,简称Poly-P)。
生物学2007-2009年3区SCI分区及影响因子

JCR2015影响因子(所有期刊从高到低排序)+中科院分区
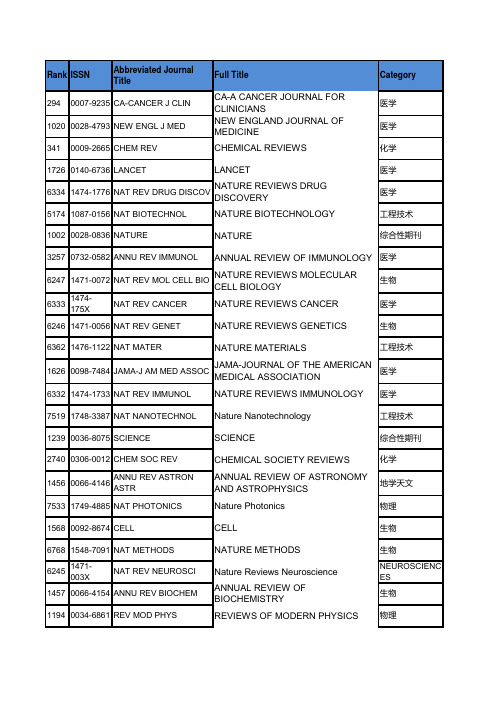
Rank ISSN Abbreviated JournalTitleFull Title Category2940007-9235CA-CANCER J CLIN CA-A CANCER JOURNAL FOR CLINICIAN医学10200028-4793NEW ENGL J MED NEW ENGLAND JOURNAL OF MEDICINE医学3410009-2665CHEM REV CHEMICAL REVIEWS化学17260140-6736LANCET LANCET医学63341474-1776NAT REV DRUG DISCOV NATURE REVIEWS DRUG DISCOVERY医学51741087-0156NAT BIOTECHNOL NATURE BIOTECHNOLOGY工程技术10020028-0836NATURE NATURE综合性期刊32570732-0582ANNU REV IMMUNOL ANNUAL REVIEW OF IMMUNOLOGY医学62471471-0072NAT REV MOL CELL BIO NATURE REVIEWS MOLECULAR CELL B生物63331474-175X NAT REV CANCER NATURE REVIEWS CANCER医学62461471-0056NAT REV GENET NATURE REVIEWS GENETICS生物63621476-1122NAT MATER NATURE MATERIALS工程技术16260098-7484JAMA-J AM MED ASSOC JAMA-JOURNAL OF THE AMERICAN ME医学63321474-1733NAT REV IMMUNOL NATURE REVIEWS IMMUNOLOGY医学75191748-3387NAT NANOTECHNOL Nature Nanotechnology工程技术12390036-8075SCIENCE SCIENCE综合性期刊27400306-0012CHEM SOC REV CHEMICAL SOCIETY REVIEWS化学14560066-4146ANNU REV ASTRON ASTR ANNUAL REVIEW OF ASTRONOMY AND地学天文75331749-4885NAT PHOTONICS Nature Photonics物理15680092-8674CELL CELL生物67681548-7091NAT METHODS NATURE METHODS生物62451471-003X NAT REV NEUROSCI Nature Reviews Neuroscience NEUROSCIENCES 14570066-4154ANNU REV BIOCHEM ANNUAL REVIEW OF BIOCHEMISTRY生物11940034-6861REV MOD PHYS REVIEWS OF MODERN PHYSICS物理48751061-4036NAT GENET NATURE GENETICS生物15060079-6425PROG MATER SCI PROGRESS IN MATERIALS SCIENCE工程技术51051078-8956NAT MED NATURE MEDICINE医学11140031-9333PHYSIOL REV PHYSIOLOGICAL REVIEWS医学15100079-6700PROG POLYM SCI PROGRESS IN POLYMER SCIENCE化学76261755-4330NAT CHEM Nature Chemistry化学62371470-2045LANCET ONCOL LANCET ONCOLOGY医学74001740-1526NAT REV MICROBIOL NATURE REVIEWS MICROBIOLOGY生物66151535-6108CANCER CELL CANCER CELL医学67091543-5008ANNU REV PLANT BIOL ANNUAL REVIEW OF PLANT BIOLOGY生物63191473-3099LANCET INFECT DIS LANCET INFECTIOUS DISEASES医学150001-4842ACCOUNTS CHEM RES ACCOUNTS OF CHEMICAL RESEARCH化学80331934-5909CELL STEM CELL Cell Stem Cell生物57941364-6613TRENDS COGN SCI TRENDS IN COGNITIVE SCIENCES医学63351474-4422LANCET NEUROL LANCET NEUROLOGY医学14660066-4308ANNU REV PSYCHOL ANNUAL REVIEW OF PSYCHOLOGY医学50621074-7613IMMUNITY IMMUNITY医学19000163-769X ENDOCR REV ENDOCRINE REVIEWS医学580001-8732ADV PHYS ADVANCES IN PHYSICS物理17250140-525X BEHAV BRAIN SCI BEHAVIORAL AND BRAIN SCIENCES医学76141754-5692ENERG ENVIRON SCI Energy & Environmental Science化学74791745-2473NAT PHYS Nature Physics物理29820370-1573PHYS REP PHYSICS REPORTS-REVIEW SECTION 物理65501529-2908NAT IMMUNOL NATURE IMMUNOLOGY医学61991465-7392NAT CELL BIOL NATURE CELL BIOLOGY生物84372159-8274CANCER DISCOV Cancer discovery ONCOLOGY 18180147-006X ANNU REV NEUROSCI ANNUAL REVIEW OF NEUROSCIENCE医学60511433-8351LIVING REV RELATIV Living Reviews in Relativity物理28850360-1285PROG ENERG COMBUST PROGRESS IN ENERGY AND COMBUST工程技术医学68241553-4006ANNU REV PATHOL-MECH Annual Review of Pathology-Mechanisms o14640066-4278ANNU REV PHYSIOL ANNUAL REVIEW OF PHYSIOLOGY医学32580732-183X J CLIN ONCOL JOURNAL OF CLINICAL ONCOLOGY医学29200362-1642ANNU REV PHARMACOL ANNUAL REVIEW OF PHARMACOLOGY 医学1520003-4819ANN INTERN MED ANNALS OF INTERNAL MEDICINE医学地学天文39930935-4956ASTRON ASTROPHYS REV ASTRONOMY AND ASTROPHYSICS REV71031614-4961LIVING REV SOL PHYS LIVING REVIEWS IN SOLAR PHYSICS ASTRONOMY & AS 67881550-4131CELL METAB Cell Metabolism生物39970935-9648ADV MATER ADVANCED MATERIALS工程技术76391756-1833BMJ-BRIT MED J BMJ-British Medical Journal MEDICINE, GENER 36650893-8512CLIN MICROBIOL REV CLINICAL MICROBIOLOGY REVIEWS医学1830003-9926ARCH INTERN MED ARCHIVES OF INTERNAL MEDICINE医学10940031-6997PHARMACOL REV PHARMACOLOGICAL REVIEWS医学680002-5100ALDRICHIM ACTA ALDRICHIMICA ACTA化学11890034-4885REP PROG PHYS REPORTS ON PROGRESS IN PHYSICS物理生物26600301-5556ADV ANAT EMBRYOL CEL ADVANCES IN ANATOMY EMBRYOLOGY化学14630066-426X ANNU REV PHYS CHEM ANNUAL REVIEW OF PHYSICAL CHEMIS5210016-5085GASTROENTEROLOGY GASTROENTEROLOGY医学51251081-0706ANNU REV CELL DEV BI ANNUAL REVIEW OF CELL AND DEVELO生物医学32960735-1097J AM COLL CARDIOL JOURNAL OF THE AMERICAN COLLEGE20440169-5347TRENDS ECOL EVOL TRENDS IN ECOLOGY & EVOLUTION生物71041614-6832ADV ENERGY MATER Advanced Energy Materials CHEMISTRY, PHYS 52791097-6256NAT NEUROSCI NATURE NEUROSCIENCE医学化学59481389-5567J PHOTOCH PHOTOBIO C JOURNAL OF PHOTOCHEMISTRY AND P81491946-6234SCI TRANSL MED Science Translational Medicine CELL BIOLOGY医学64501520-765X EUR HEART J SUPPL EUROPEAN HEART JOURNAL SUPPLEM14600066-4197ANNU REV GENET ANNUAL REVIEW OF GENETICS生物39160927-796X MAT SCI ENG R MATERIALS SCIENCE & ENGINEERING 工程技术80491936-122X ANNU REV BIOPHYS Annual Review of Biophysics生物76891759-4758NAT REV NEUROL Nature Reviews Neurology医学21640195-668X EUR HEART J EUROPEAN HEART JOURNAL医学37130896-6273NEURON NEURON医学20380169-409X ADV DRUG DELIVER REV ADVANCED DRUG DELIVERY REVIEWS医学75131748-0132NANO TODAY Nano Today工程技术86088755-1209REV GEOPHYS REVIEWS OF GEOPHYSICS地学81551947-5454ANNU REV CONDEN MA P Annual Review of Condensed Matter Physi PHYSICS, CONDEN 19620167-5729SURF SCI REP SURFACE SCIENCE REPORTS化学11550033-2909PSYCHOL BULL PSYCHOLOGICAL BULLETIN医学5530017-5749GUT GUT医学51931088-9051GENOME RES GENOME RESEARCH生物生物52311092-2172MICROBIOL MOL BIOL R MICROBIOLOGY AND MOLECULAR BIOL83072047-7538LIGHT-SCI APPL Light-Science & Applications OPTICS 76811758-678X NAT CLIM CHANGE Nature climate change ENVIRONMENTAL 57581359-4184MOL PSYCHIATR MOLECULAR PSYCHIATRY医学1820003-990X ARCH GEN PSYCHIAT ARCHIVES OF GENERAL PSYCHIATRY医学3590009-7322CIRCULATION CIRCULATION医学67721549-1676PLOS MED PLOS MEDICINE MEDICINE, GENER 49041063-5157SYST BIOL SYSTEMATIC BIOLOGY生物81041941-1405ANNU REV MAR SCI Annual Review of Marine Science地学73441723-8617WORLD PSYCHIATRY World Psychiatry医学64881523-9829ANNU REV BIOMED ENG ANNUAL REVIEW OF BIOMEDICAL ENG工程技术76901759-4774NAT REV CLIN ONCOL Nature Reviews Clinical Oncology医学58401369-7021MATER TODAY Materials Today工程技术52771097-2765MOL CELL MOLECULAR CELL生物26720302-2838EUR UROL EUROPEAN UROLOGY医学14580066-4170ANNU REV ENTOMOL ANNUAL REVIEW OF ENTOMOLOGY生物65641530-6984NANO LETT NANO LETTERS工程技术19380166-2236TRENDS NEUROSCI TRENDS IN NEUROSCIENCES医学67381545-9993NAT STRUCT MOL BIOL NATURE STRUCTURAL & MOLECULAR BBIOCHEMISTRY & 76941759-5029NAT REV ENDOCRINOL Nature Reviews Endocrinology医学19350166-0616STUD MYCOL STUDIES IN MYCOLOGY生物20150168-6445FEMS MICROBIOL REV FEMS MICROBIOLOGY REVIEWS生物医学7260021-9738J CLIN INVEST JOURNAL OF CLINICAL INVESTIGATION84752168-6106JAMA INTERN MED JAMA Internal Medicine MEDICINE, GENER医学50501073-449X AM J RESP CRIT CARE AMERICAN JOURNAL OF RESPIRATORY68091552-4450NAT CHEM BIOL Nature Chemical Biology生物57731360-1385TRENDS PLANT SCI TRENDS IN PLANT SCIENCE生物14610066-4219ANNU REV MED ANNUAL REVIEW OF MEDICINE医学80481936-0851ACS NANO ACS Nano工程技术医学67671548-5943ANNU REV CLIN PSYCHO ANNUAL REVIEW OF CLINICAL PSYCHO76951759-5045NAT REV GASTRO HEPAT Nature Reviews Gastroenterology & Hepat医学10000027-8874JNCI-J NATL CANCER I JNCI-Journal of the National Cancer Institu ONCOLOGY医学7570022-1007J EXP MED JOURNAL OF EXPERIMENTAL MEDICINE44551001-0602CELL RES CELL RESEARCH生物68181552-5260ALZHEIMERS DEMENT Alzheimers & Dementia医学79881931-3128CELL HOST MICROBE Cell Host & Microbe生物1050002-953X AM J PSYCHIAT AMERICAN JOURNAL OF PSYCHIATRY医学3970010-8545COORDIN CHEM REV COORDINATION CHEMISTRY REVIEWS化学14620066-4227ANNU REV MICROBIOL ANNUAL REVIEW OF MICROBIOLOGY生物770002-7863J AM CHEM SOC JOURNAL OF THE AMERICAN CHEMICA化学84802168-622X JAMA PSYCHIAT JAMA Psychiatry PSYCHIATRY 43000962-8924TRENDS CELL BIOL TRENDS IN CELL BIOLOGY生物19870167-7799TRENDS BIOTECHNOL TRENDS IN BIOTECHNOLOGY工程技术76871759-0876WIRES COMPUT MOL SCI W IREs Computational Molecular Science CHEMISTRY, MUL 65801531-7331ANNU REV MATER RES ANNUAL REVIEW OF MATERIALS RESE工程技术1150003-0007B AM METEOROL SOC BULLETIN OF THE AMERICAN METEOR地学71151616-301X ADV FUNCT MATER ADVANCED FUNCTIONAL MATERIALS工程技术68331554-8627AUTOPHAGY Autophagy生物75871752-0894NAT GEOSCI Nature Geoscience地学医学19310165-6147TRENDS PHARMACOL SCI TRENDS IN PHARMACOLOGICAL SCIEN15560091-6749J ALLERGY CLIN IMMUN JOURNAL OF ALLERGY AND CLINICAL I医学82642041-1723NAT COMMUN Nature communications Biological Science Disciplines 20200168-8278J HEPATOL JOURNAL OF HEPATOLOGY医学化学60491433-7851ANGEW CHEM INT EDIT ANGEWANDTE CHEMIE-INTERNATIONA19030163-8998ANNU REV NUCL PART S ANNUAL REVIEW OF NUCLEAR AND PA物理43630968-0004TRENDS BIOCHEM SCI TRENDS IN BIOCHEMICAL SCIENCES生物14690067-0049ASTROPHYS J SUPPL S ASTROPHYSICAL JOURNAL SUPPLEME地学天文物理14590066-4189ANNU REV FLUID MECH ANNUAL REVIEW OF FLUID MECHANICS24880270-9139HEPATOLOGY HEPATOLOGY医学3600009-7330CIRC RES CIRCULATION RESEARCH医学940002-9297AM J HUM GENET AMERICAN JOURNAL OF HUMAN GENE生物74601744-4292MOL SYST BIOL Molecular Systems Biology生物61981465-6906GENOME BIOL GENOME BIOLOGY BIOTECHNOLOGY 36240890-9369GENE DEV GENES & DEVELOPMENT生物370001-6322ACTA NEUROPATHOL ACTA NEUROPATHOLOGICA医学930002-9270AM J GASTROENTEROL AMERICAN JOURNAL OF GASTROENTE医学61551461-023X ECOL LETT ECOLOGY LETTERS环境科学67111543-592X ANNU REV ECOL EVOL S ANNUAL REVIEW OF ECOLOGY EVOLU生物2530006-4971BLOOD BLOOD医学84252157-1724KIDNEY INT SUPPL Kidney International Supplements UROLOGY & NEPHROLOGY 23760261-4189EMBO J EMBO JOURNAL生物35670887-6924LEUKEMIA LEUKEMIA医学62881471-4906TRENDS IMMUNOL TRENDS IN IMMUNOLOGY医学医学1580003-4967ANN RHEUM DIS ANNALS OF THE RHEUMATIC DISEASES85252211-2855NANO ENERGY Nano Energy CHEMISTRY, PHYS 2400006-3223BIOL PSYCHIAT BIOLOGICAL PSYCHIATRY医学16200098-2997MOL ASPECTS MED MOLECULAR ASPECTS OF MEDICINE医学57341355-4786HUM REPROD UPDATE HUMAN REPRODUCTION UPDATE医学16720105-2896IMMUNOL REV IMMUNOLOGICAL REVIEWS医学79641884-4049NPG ASIA MATER NPG Asia Materials MATERIALS SCIEN 81361943-8206ADV OPT PHOTONICS Advances in Optics and Photonics OPTICS24090265-0568NAT PROD REP NATURAL PRODUCT REPORTS化学85472214-109X LANCET GLOB HEALTH Lancet Global Health PUBLIC, ENVIRON 19010163-7827PROG LIPID RES PROGRESS IN LIPID RESEARCH生物28830360-0564ADV CATAL ADVANCES IN CATALYSIS化学26360301-0082PROG NEUROBIOL PROGRESS IN NEUROBIOLOGY医学29480364-5134ANN NEUROL ANNALS OF NEUROLOGY医学20260168-9525TRENDS GENET TRENDS IN GENETICS生物76911759-4790NAT REV RHEUMATOL Nature Reviews Rheumatology医学7130021-9525J CELL BIOLJOURNAL OF CELL BIOLOGY 生物18980163-7258PHARMACOL THERAPEUT P HARMACOLOGY & THERAPEUTICS 医学66051534-5807DEV CELL DEVELOPMENTAL CELL 生物9990027-8424P NATL ACAD SCI USA PROCEEDINGS OF THE NATIONAL ACA 综合性期刊76111754-2189NAT PROTOC Nature Protocols 生物61911464-7931BIOL REV BIOLOGICAL REVIEWS 生物85402213-2600LANCET RESP MED Lancet Respiratory Medicine RESPIRATORY SYSTEM 14650066-4286ANNU REV PHYTOPATHOL ANNUAL REVIEW OF PHYTOPATHOLOG 生物62141467-5463BRIEF BIOINFORM BRIEFINGS IN BIOINFORMATICS 生物33610742-3098J PINEAL RES JOURNAL OF PINEAL RESEARCH 医学42810960-9822CURR BIOL CURRENT BIOLOGY 生物84242157-1422CSH PERSPECT MED Cold Spring Harbor perspectives in medicin MEDICINE, RESEA 62891471-4914TRENDS MOL MED TRENDS IN MOLECULAR MEDICINE 医学47001043-2760TRENDS ENDOCRIN MET TRENDS IN ENDOCRINOLOGY AND MET 医学47361046-6673J AM SOC NEPHROL JOURNAL OF THE AMERICAN SOCIETY 医学67321545-7885PLOS BIOL PLoS Biology BIOCHEMISTRY &46501040-4651PLANT CELL PLANT CELL 生物47171044-579X SEMIN CANCER BIOL SEMINARS IN CANCER BIOLOGY 医学3220008-5472CANCER RES CANCER RESEARCH 医学83162050-084X ELIFE eLife BIOLOGY 84112155-5435ACS CATAL ACS catalysis CHEMISTRY, PHYSICAL 75601751-7362ISME J ISME Journal 环境科学82692041-6520CHEM SCI Chemical science CHEMISTRY, MULT 2610006-8950BRAIN BRAIN 医学11620033-3190PSYCHOTHER PSYCHOSOM PSYCHOTHERAPY AND PSYCHOSOMAT 医学43460966-842X TRENDS MICROBIOL TRENDS IN MICROBIOLOGY 生物85462213-8587LANCET DIABETES ENDO Lancet Diabetes & Endocrinology ENDOCRINOLOGY 76931759-5002NAT REV CARDIOL Nature Reviews Cardiology 医学26130300-5771INT J EPIDEMIOL INTERNATIONAL JOURNAL OF EPIDEMI 医学58301368-7646DRUG RESIST UPDATE DRUG RESISTANCE UPDATES 医学27240305-1048NUCLEIC ACIDS RES NUCLEIC ACIDS RESEARCH 生物33250737-4038MOL BIOL EVOL MOLECULAR BIOLOGY AND EVOLUTION 生物62311469-221X EMBO REP EMBO REPORTS 生物84392160-3308PHYS REV X Physical Review X PHYSICS, MULTIDISCIPLINAR 32930734-9750BIOTECHNOL ADV BIOTECHNOLOGY ADVANCES 工程技术65391527-8204ANNU REV GENOM HUM G A NNUAL REVIEW OF GENOMICS AND H 生物48421058-4838CLIN INFECT DIS CLINICAL INFECTIOUS DISEASES 医学80501936-1327ANNU REV ANAL CHEM Annual Review of Analytical Chemistry CHEMISTRY, ANAL 18530149-7634NEUROSCI BIOBEHAV R NEUROSCIENCE AND BIOBEHAVIORAL 医学49101063-6706IEEE T FUZZY SYST IEEE TRANSACTIONS ON FUZZY SYSTE 工程技术56991350-9462PROG RETIN EYE RES PROGRESS IN RETINAL AND EYE RESE 医学50971078-0432CLIN CANCER RES CLINICAL CANCER RESEARCH 医学81271943-0264CSH PERSPECT BIOL Cold Spring Harbor perspectives in biology CELL BIOLOGY 81541947-5438ANNU REV CHEM BIOMOL Annual review of chemical and biomolecula CHEMISTRY, APPL 47411047-3211CEREB CORTEX CEREBRAL CORTEX 医学76581757-4676EMBO MOL MED EMBO Molecular Medicine 医学15250084-6597ANNU REV EARTH PL SC ANNUAL REVIEW OF EARTH AND PLAN地学天文85372212-6864PHYS DARK UNIVERSE Physics of the Dark Universe ASTRONOMY & ASTROPHYSI 15260085-2538KIDNEY INT KIDNEY INTERNATIONAL医学17610142-9612BIOMATERIALS BIOMATERIALS工程技术76961759-5061NAT REV NEPHROL Nature Reviews Nephrology医学51771087-0792SLEEP MED REV SLEEP MEDICINE REVIEWS医学41100950-6608INT MATER REV INTERNATIONAL MATERIALS REVIEWS工程技术化学18720161-4940CATAL REV CATALYSIS REVIEWS-SCIENCE AND EN41800955-0674CURR OPIN CELL BIOL CURRENT OPINION IN CELL BIOLOGY生物41120950-9232ONCOGENE ONCOGENE医学31840586-7614SCHIZOPHRENIA BULL SCHIZOPHRENIA BULLETIN医学21910198-6325MED RES REV MEDICINAL RESEARCH REVIEWS医学18490149-5992DIABETES CARE DIABETES CARE医学37140896-8411J AUTOIMMUN JOURNAL OF AUTOIMMUNITY医学71011613-6810SMALL SMALL工程技术21950199-9885ANNU REV NUTR ANNUAL REVIEW OF NUTRITION医学85242211-1247CELL REP Cell reports CELL BIOLOGY37210897-4756CHEM MATER CHEMISTRY OF MATERIALS工程技术1570003-4932ANN SURG ANNALS OF SURGERY医学13180040-6376THORAX THORAX医学10150028-3878NEUROLOGY NEUROLOGY医学62101467-2960FISH FISH FISH AND FISHERIES农林科学56481342-937X GONDWANA RES GONDWANA RESEARCH地学56981350-9047CELL DEATH DIFFER CELL DEATH AND DIFFERENTIATION生物85532223-7755GEOCHEM PERSPECT Geochemical Perspectives GEOCHEMISTRY & 4220012-1797DIABETES DIABETES医学57201354-1013GLOBAL CHANGE BIOL GLOBAL CHANGE BIOLOGY环境科学77831838-7640THERANOSTICS THERANOSTICS MEDICINE, RESEA 61781463-9262GREEN CHEM GREEN CHEMISTRY化学78381863-8880LASER PHOTONICS REV Laser & Photonics Reviews物理62171467-7881OBES REV Obesity Reviews医学2760007-1250BRIT J PSYCHIAT BRITISH JOURNAL OF PSYCHIATRY医学74121741-7007BMC BIOL BMC BIOLOGY生物环境科学15570091-6765ENVIRON HEALTH PERSP ENVIRONMENTAL HEALTH PERSPECTIV11570033-295X PSYCHOL REV PSYCHOLOGICAL REVIEW医学69511568-9972AUTOIMMUN REV AUTOIMMUNITY REVIEWS医学3680009-9147CLIN CHEM CLINICAL CHEMISTRY医学医学3730009-9236CLIN PHARMACOL THER CLINICAL PHARMACOLOGY & THERAPE67011542-3565CLIN GASTROENTEROL H Clinical Gastroenterology and Hepatology医学4390012-8252EARTH-SCI REV EARTH-SCIENCE REVIEWS地学58361369-5266CURR OPIN PLANT BIOL CURRENT OPINION IN PLANT BIOLOGY生物医学27100304-419X BBA-REV CANCER BIOCHIMICA ET BIOPHYSICA ACTA-REV生物11730033-5835Q REV BIOPHYS QUARTERLY REVIEWS OF BIOPHYSICS化学560001-8686ADV COLLOID INTERFAC ADVANCES IN COLLOID AND INTERFAC1990004-3591ARTHRITIS RHEUM-US ARTHRITIS & RHEUMATISM-ARTHRITIS医学78321863-2297SEMIN IMMUNOPATHOL Seminars in Immunopathology医学46731040-9238CRIT REV BIOCHEM MOL CRITICAL REVIEWS IN BIOCHEMISTRY 生物25510277-7037MASS SPECTROM REV MASS SPECTROMETRY REVIEWS物理20130168-3659J CONTROL RELEASE JOURNAL OF CONTROLLED RELEASE医学10210028-646X NEW PHYTOL NEW PHYTOLOGIST生物78421864-5631CHEMSUSCHEM ChemSusChem化学37510903-1936EUR RESPIR J EUROPEAN RESPIRATORY JOURNAL医学27360305-7372CANCER TREAT REV CANCER TREATMENT REVIEWS医学48921062-7995PROG PHOTOVOLTAICS PROGRESS IN PHOTOVOLTAICS工程技术生物42350959-437X CURR OPIN GENET DEV CURRENT OPINION IN GENETICS & DEV68281553-7366PLOS PATHOG PLoS Pathogens生物68291553-7390PLOS GENET PLoS Genetics生物11050031-9007PHYS REV LETT PHYSICAL REVIEW LETTERS物理65491529-1839OPTOMETRY OPTOMETRY OPHTHALMOLOGY4270012-3692CHEST CHEST医学41370952-7915CURR OPIN IMMUNOL CURRENT OPINION IN IMMUNOLOGY医学69851571-0645PHYS LIFE REV Physics of Life Reviews生物81631948-7185J PHYS CHEM LETT The journal of physical chemistry letters CHEMISTRY, PHYS 83212050-7488J MATER CHEM A Journal of Materials Chemistry A CHEMISTRY, PHYS环境科学66791540-9295FRONT ECOL ENVIRON FRONTIERS IN ECOLOGY AND THE ENV环境科学38970926-3373APPL CATAL B-ENVIRON APPLIED CATALYSIS B-ENVIRONMENTA8310022-3417J PATHOL JOURNAL OF PATHOLOGY医学1840003-9942ARCH NEUROL-CHICAGO ARCHIVES OF NEUROLOGY医学64781523-0864ANTIOXID REDOX SIGN ANTIOXIDANTS & REDOX SIGNALING生物82562040-3364NANOSCALE Nanoscale CHEMISTRY, MUL80181933-0219MUCOSAL IMMUNOL Mucosal Immunology医学75341749-5016SOC COGN AFFECT NEUR Social Cognitive and Affective Neuroscienc医学42930962-4929ACTA NUMER ACTA NUMERICA MATHEMATICS80611936-8798JACC-CARDIOVASC INTE JACC. Cardiovascular interventions CARDIAC & CARDI 52851098-3600GENET MED GENETICS IN MEDICINE医学1190003-0090B AM MUS NAT HIST BULLETIN OF THE AMERICAN MUSEUM环境科学84892190-5991J CACHEXIA SARCOPENI Journal of Cachexia Sarcopenia and Muscl MEDICINE, GENER 84762168-6149JAMA NEUROL JAMA Neurology CLINICAL NEUROLOGY医学36210890-8567J AM ACAD CHILD PSY JOURNAL OF THE AMERICAN ACADEMY74131741-7015BMC MED BMC Medicine医学76241755-263X CONSERV LETT Conservation Letters BIODIVERSITY CONSERVATIO物理15070079-6565PROG NUCL MAG RES SP PROGRESS IN NUCLEAR MAGNETIC RE19850167-7659CANCER METAST REV CANCER AND METASTASIS REVIEWS医学医学7860022-202X J INVEST DERMATOL JOURNAL OF INVESTIGATIVE DERMATO28540342-4642INTENS CARE MED INTENSIVE CARE MEDICINE医学42370959-440X CURR OPIN STRUC BIOL CURRENT OPINION IN STRUCTURAL BI生物80601936-878X JACC-CARDIOVASC IMAG JACC. Cardiovascular imaging CARDIAC & CARD33330738-8551CRIT REV BIOTECHNOL CRITICAL REVIEWS IN BIOTECHNOLOG工程技术84792168-6203JAMA PEDIATR JAMA Pediatrics PEDIATRICS42210958-1669CURR OPIN BIOTECH CURRENT OPINION IN BIOTECHNOLOG工程技术74501743-8977PART FIBRE TOXICOL Particle and fibre toxicology TOXICOLOGY生物42960962-8436PHILOS T R SOC B PHILOSOPHICAL TRANSACTIONS OF THNEUROPSYCHOPHARMACOLOGY医学36560893-133X NEUROPSYCHOPHARMACO38530923-7534ANN ONCOL ANNALS OF ONCOLOGY医学15500091-3022FRONT NEUROENDOCRIN FRONTIERS IN NEUROENDOCRINOLOG医学17840144-235X INT REV PHYS CHEM INTERNATIONAL REVIEWS IN PHYSICA化学82261998-0124NANO RES Nano Research化学化学14450065-3055ADV ORGANOMET CHEM ADVANCES IN ORGANOMETALLIC CHEM4410012-9615ECOL MONOGR ECOLOGICAL MONOGRAPHS环境科学17300140-7791PLANT CELL ENVIRON PLANT CELL AND ENVIRONMENT生物7120021-9517J CATAL JOURNAL OF CATALYSIS工程技术11830033-8419RADIOLOGY RADIOLOGY医学11250032-0889PLANT PHYSIOL PLANT PHYSIOLOGY生物50551073-8584NEUROSCIENTIST NEUROSCIENTIST医学57701359-7345CHEM COMMUN CHEMICAL COMMUNICATIONS化学35080884-0431J BONE MINER RES JOURNAL OF BONE AND MINERAL RES医学生物58211367-5931CURR OPIN CHEM BIOL CURRENT OPINION IN CHEMICAL BIOLO8170022-3050J NEUROL NEUROSUR PS JOURNAL OF NEUROLOGY NEUROSUR医学COMPUTER SCIEN 68301553-877X IEEE COMMUN SURV TUT IEEE COMMUNICATIONS SURVEYS AND83942150-7511MBIO mBio MICROBIOLOGY 72461674-2788J MOL CELL BIOL Journal of molecular cell biology CELL BIOLOGY 890002-9165AM J CLIN NUTR AMERICAN JOURNAL OF CLINICAL NUT医学52751096-7176METAB ENG METABOLIC ENGINEERING工程技术14380065-2660ADV GENET ADVANCES IN GENETICS生物51221080-6040EMERG INFECT DIS EMERGING INFECTIOUS DISEASES医学81421944-8244ACS APPL MATER INTER ACS Applied Materials & Interfaces工程技术57651359-6446DRUG DISCOV TODAY DRUG DISCOVERY TODAY医学4230012-186X DIABETOLOGIA DIABETOLOGIA医学21520193-936X EPIDEMIOL REV EPIDEMIOLOGIC REVIEWS医学医学47911053-2498J HEART LUNG TRANSPL JOURNAL OF HEART AND LUNG TRANS50601074-5521CHEM BIOL CHEMISTRY & BIOLOGY生物42360959-4388CURR OPIN NEUROBIOL CURRENT OPINION IN NEUROBIOLOGY医学化学15120079-6786PROG SOLID STATE CH PROGRESS IN SOLID STATE CHEMISTR66181535-9476MOL CELL PROTEOMICS MOLECULAR & CELLULAR PROTEOMIC生物75431750-1326MOL NEURODEGENER Molecular Neurodegeneration医学82652041-210X METHODS ECOL EVOL Methods in ecology and evolution ECOLOGY环境科学62071466-822X GLOBAL ECOL BIOGEOGR GLOBAL ECOLOGY AND BIOGEOGRAPH59381388-9842EUR J HEART FAIL EUROPEAN JOURNAL OF HEART FAILU医学49671066-5099STEM CELLS STEM CELLS医学17070129-0657INT J NEURAL SYST International Journal of Neural Systems工程技术25530278-0046IEEE T IND ELECTRON IEEE TRANSACTIONS ON INDUSTRIAL E工程技术42900962-1083MOL ECOL MOLECULAR ECOLOGY生物21560194-911X HYPERTENSION HYPERTENSION医学19340165-9936TRAC-TREND ANAL CHEM TRAC-TRENDS IN ANALYTICAL CHEMIS化学18990163-7525ANNU REV PUBL HEALTH ANNUAL REVIEW OF PUBLIC HEALTH医学41020950-1991DEVELOPMENT DEVELOPMENT生物医学7220021-9630J CHILD PSYCHOL PSYC JOURNAL OF CHILD PSYCHOLOGY AND物理46661040-8436CRIT REV SOLID STATE CRITICAL REVIEWS IN SOLID STATE AN81451945-4589AGING-US Aging CELL BIOLOGY 59271387-6473NEW ASTRON REV NEW ASTRONOMY REVIEWS地学天文74431743-5390NANOTOXICOLOGY Nanotoxicology医学41970956-5663BIOSENS BIOELECTRON BIOSENSORS & BIOELECTRONICS工程技术11870034-4257REMOTE SENS ENVIRON REMOTE SENSING OF ENVIRONMENT环境科学43220964-6906HUM MOL GENET HUMAN MOLECULAR GENETICS生物64871523-7060ORG LETT ORGANIC LETTERS化学61691462-8902DIABETES OBES METAB DIABETES OBESITY & METABOLISM医学81681949-2553ONCOTARGET Oncotarget Oncogenes 47971053-8119NEUROIMAGE NEUROIMAGE医学24860270-6474J NEUROSCI JOURNAL OF NEUROSCIENCE医学63411474-9718AGING CELL AGING CELL生物72451674-2052MOL PLANT Molecular Plant生物8020022-2593J MED GENET JOURNAL OF MEDICAL GENETICS医学79091874-9399BBA-GENE REGUL MECH Biochimica et Biophysica Acta-Gene Regul生物15320090-3493CRIT CARE MED CRITICAL CARE MEDICINE医学81051941-1413ANNU REV FOOD SCI T Annual review of food science and technoloFOOD SCIENCE & 12760038-6308SPACE SCI REV SPACE SCIENCE REVIEWS地学天文81441945-0877SCI SIGNAL Science Signaling BIOCHEMISTRY & 26820303-4240REV PHYSIOL BIOCH P REVIEWS OF PHYSIOLOGY BIOCHEMIS医学84482162-4011ONCOIMMUNOLOGY OncoImmunology ONCOLOGY生物51631084-9521SEMIN CELL DEV BIOL SEMINARS IN CELL & DEVELOPMENTAL9450025-6196MAYO CLIN PROC MAYO CLINIC PROCEEDINGS医学工程技术57541359-0286CURR OPIN SOLID ST M CURRENT OPINION IN SOLID STATE & M79001873-9946J CROHNS COLITIS Journal of Crohns & Colitis医学64971525-0016MOL THER MOLECULAR THERAPY医学14460065-308X ADV PARASIT ADVANCES IN PARASITOLOGY医学68991560-2745FUNGAL DIVERS FUNGAL DIVERSITY生物51441083-4419IEEE T SYST MAN CY B IEEE TRANSACTIONS ON SYSTEMS MA工程技术81121941-7640CIRC-CARDIOVASC INTE Circulation: Cardiovascular Interventions CARDIAC & CARDI 30630378-7753J POWER SOURCES JOURNAL OF POWER SOURCES工程技术生物33830748-3007CLADISTICS CLADISTICS-THE INTERNATIONAL JOUR生物58771383-5742MUTAT RES-REV MUTAT MUTATION RESEARCH-REVIEWS IN MU7250021-972X J CLIN ENDOCR METAB JOURNAL OF CLINICAL ENDOCRINOLO医学62901471-4922TRENDS PARASITOL TRENDS IN PARASITOLOGY医学61641462-2912ENVIRON MICROBIOL ENVIRONMENTAL MICROBIOLOGY环境科学3240008-6223CARBON CARBON工程技术47121044-3983EPIDEMIOLOGY EPIDEMIOLOGY医学36600893-3952MODERN PATHOL MODERN PATHOLOGY医学18730161-5505J NUCL MED JOURNAL OF NUCLEAR MEDICINE医学68751558-3724POLYM REV Polymer Reviews工程技术67831549-9634NANOMED-NANOTECHNOL Nanomedicine-Nanotechnology Biology an工程技术18760161-6420OPHTHALMOLOGY OPHTHALMOLOGY医学29840370-2693PHYS LETT B PHYSICS LETTERS B物理63151473-0197LAB CHIP LAB ON A CHIP工程技术46281029-8479J HIGH ENERGY PHYS JOURNAL OF HIGH ENERGY PHYSICS PHYSICS, PARTICL10420029-6643NUTR REV NUTRITION REVIEWS医学79501879-6257CURR OPIN VIROL Current Opinion in Virology VIROLOGY 41750954-6820J INTERN MED JOURNAL OF INTERNAL MEDICINE医学76151754-6834BIOTECHNOL BIOFUELS Biotechnology for Biofuels工程技术医学62321469-493X COCHRANE DB SYST REV Cochrane Database of Systematic Reviews36740894-1491GLIA GLIA医学16730105-4538ALLERGY ALLERGY医学74331742-7061ACTA BIOMATER Acta Biomaterialia工程技术46641040-841X CRIT REV MICROBIOL CRITICAL REVIEWS IN MICROBIOLOGY生物76631757-7004WIRES RNA WIRES RNA CELL BIOLOGY工程技术35420885-8993IEEE T POWER ELECTR IEEE TRANSACTIONS ON POWER ELEC51121079-5642ARTERIOSCL THROM VAS ARTERIOSCLEROSIS THROMBOSIS AN医学7830022-1899J INFECT DIS JOURNAL OF INFECTIOUS DISEASES医学2090004-637X ASTROPHYS J ASTROPHYSICAL JOURNAL地学天文28390340-5761ARCH TOXICOL ARCHIVES OF TOXICOLOGY医学42700960-7412PLANT J PLANT JOURNAL生物49631065-9471HUM BRAIN MAPP HUMAN BRAIN MAPPING医学5280016-6731GENETICS GENETICS生物14410065-2776ADV IMMUNOL ADVANCES IN IMMUNOLOGY医学34470820-3946CAN MED ASSOC J CANADIAN MEDICAL ASSOCIATION JOU医学3260008-6363CARDIOVASC RES CARDIOVASCULAR RESEARCH医学11560033-2917PSYCHOL MED PSYCHOLOGICAL MEDICINE医学43190964-4563TOB CONTROL TOBACCO CONTROL医学57881364-0321RENEW SUST ENERG REV R ENEWABLE & SUSTAINABLE ENERGY工程技术医学25140272-6386AM J KIDNEY DIS AMERICAN JOURNAL OF KIDNEY DISEA58371369-5274CURR OPIN MICROBIOL CURRENT OPINION IN MICROBIOLOGY生物环境科学67121543-5938ANNU REV ENV RESOUR ANNUAL REVIEW OF ENVIRONMENT AN81081941-3289CIRC-HEART FAIL Circulation-Heart Failure医学35880889-1591BRAIN BEHAV IMMUN BRAIN BEHAVIOR AND IMMUNITY医学1350003-3022ANESTHESIOLOGY ANESTHESIOLOGY医学47501049-250X ADV ATOM MOL OPT PHY ADVANCES IN ATOMIC MOLECULAR AN物理医学53951180-4882J PSYCHIATR NEUROSCI JOURNAL OF PSYCHIATRY & NEUROSC47961053-5888IEEE SIGNAL PROC MAG IEEE SIGNAL PROCESSING MAGAZINE工程技术57551359-0294CURR OPIN COLLOID IN CURRENT OPINION IN COLLOID & INTE化学31020390-6078HAEMATOLOGICA HAEMATOLOGICA HEMATOLOGY地学天文63551475-7516J COSMOL ASTROPART P JOURNAL OF COSMOLOGY AND ASTRO生物60021420-682X CELL MOL LIFE SCI CMLS-Cellular and Molecular Life Sciences9170024-9297MACROMOLECULES MACROMOLECULES工程技术84962192-2640ADV HEALTHC MATER Advanced healthcare materials工程技术82952046-2441OPEN BIOL Open biology BIOCHEMISTRY &工程技术18870162-8828IEEE T PATTERN ANAL IEEE TRANSACTIONS ON PATTERN ANA54041198-743X CLIN MICROBIOL INFEC CLINICAL MICROBIOLOGY AND INFECT医学84402161-1653ACS MACRO LETT ACS Macro Letters化学62151467-7644PLANT BIOTECHNOL J PLANT BIOTECHNOLOGY JOURNAL生物62121467-3037CURR ISSUES MOL BIOL CURRENT ISSUES IN MOLECULAR BIOL生物65071525-7797BIOMACROMOLECULES BIOMACROMOLECULES化学36340891-5849FREE RADICAL BIO MED FREE RADICAL BIOLOGY AND MEDICIN医学40690947-6539CHEM-EUR J CHEMISTRY-A EUROPEAN JOURNAL化学50391072-4710ARCH PEDIAT ADOL MED ARCHIVES OF PEDIATRICS & ADOLESC医学医学24640269-2813ALIMENT PHARM THER ALIMENTARY PHARMACOLOGY & THER9590025-7974MEDICINE MEDICINE医学12840039-2499STROKE STROKE医学69011560-7917EUROSURVEILLANCE EUROSURVEILLANCE INFECTIOUS DISEASES 66541538-7933J THROMB HAEMOST JOURNAL OF THROMBOSIS AND HAEM医学84262157-6564STEM CELL TRANSL MED Stem cells translational medicine CELL & TISSUE ENGINEERING 15130079-6816PROG SURF SCI PROGRESS IN SURFACE SCIENCE工程技术84982192-8606NANOPHOTONICS-BERLIN N anophotonics NANOSCIENCE &66161535-7163MOL CANCER THER MOLECULAR CANCER THERAPEUTICS医学医学70591600-6135AM J TRANSPLANT AMERICAN JOURNAL OF TRANSPLANTA35210885-3185MOVEMENT DISORD MOVEMENT DISORDERS医学35010883-7694MRS BULL MRS BULLETIN工程技术26690301-9268PRECAMBRIAN RES PRECAMBRIAN RESEARCH地学71071615-4150ADV SYNTH CATAL ADVANCED SYNTHESIS & CATALYSIS化学42180957-9672CURR OPIN LIPIDOL CURRENT OPINION IN LIPIDOLOGY医学81131941-7705CIRC-CARDIOVASC QUAL Circulation: Cardiovascular Quality and Ou CARDIAC & CARDI1320003-2700ANAL CHEM ANALYTICAL CHEMISTRY化学26970304-3835CANCER LETT CANCER LETTERS医学84312158-3188TRANSL PSYCHIAT Translational Psychiatry PSYCHIATRY43740969-2126STRUCTURE STRUCTURE生物78331863-2653BRAIN STRUCT FUNCT Brain Structure & Function医学27410306-2619APPL ENERG APPLIED ENERGY工程技术450001-690X ACTA PSYCHIAT SCAND ACTA PSYCHIATRICA SCANDINAVICA医学17750143-5221CLIN SCI CLINICAL SCIENCE医学57371355-6037HEART HEART医学82852045-2322SCI REP-UK Scientific reports Natural Science Disciplines 47861052-9276REV MED VIROL REVIEWS IN MEDICAL VIROLOGY医学34040749-6419INT J PLASTICITY INTERNATIONAL JOURNAL OF PLASTIC工程技术24600268-960X BLOOD REV BLOOD REVIEWS医学64761522-8517NEURO-ONCOLOGY NEURO-ONCOLOGY医学18600160-4120ENVIRON INT ENVIRONMENT INTERNATIONAL环境科学24800269-9370AIDS AIDS医学2780007-1323BRIT J SURG BRITISH JOURNAL OF SURGERY医学13480043-1354WATER RES WATER RESEARCH环境科学57241354-3784EXPERT OPIN INV DRUG EXPERT OPINION ON INVESTIGATIONA医学7550022-0957J EXP BOT JOURNAL OF EXPERIMENTAL BOTANY生物7450022-0477J ECOL JOURNAL OF ECOLOGY环境科学77031759-9954POLYM CHEM-UK POLYMER CHEMISTRY POLYMER SCIENCE 82141994-0416CRYOSPHERE Cryosphere GEOGRAPHY, PHY 18630160-6689J CLIN PSYCHIAT JOURNAL OF CLINICAL PSYCHIATRY医学67821549-9618J CHEM THEORY COMPUT J ournal of Chemical Theory and Computat化学65401527-8999CHEM REC CHEMICAL RECORD化学61951465-542X BREAST CANCER RES BREAST CANCER RESEARCH ONCOLOGY。
Appl. Environ. Microbiol.-2003-Williams-5463-71

A PPLIED AND E NVIRONMENTAL M ICROBIOLOGY,Sept.2003,p.5463–5271Vol.69,No.9 0099-2240/03/$08.00ϩ0DOI:10.1128/AEM.69.9.5463–5271.2003Copyright©2003,American Society for Microbiology.All Rights Reserved.Growth of Escherichia coli in Model Distribution System Biofilms Exposed to Hypochlorous Acid or MonochloramineMargaret M.Williams†and Ellen B.Braun-Howland*Wadsworth Center,New York State Department of Health,Albany,New York12201Received18September2002/Accepted9June2003Bacteria indigenous to water distribution systems were used to grow multispecies biofilms within continu-ous-flow slide chambers.Sixflow chambers were also inoculated with an Escherichia coli isolate obtained frompotable water.The effect of disinfectants on bacterial populations was determined after exposure of establishedbiofilms to1ppm of hypochlorous acid(ClOH)for67min or4ppm of monochloramine(NH2Cl)for155min.To test the ability of bacterial populations to initiate biofilm formation in the presence of disinfectants,weassessed the biofilms after2weeks of exposure to residual concentrations of0.2ppm of ClOH or4ppm ofstly,to determine the effect of recommended residual concentrations on newly established biofilms,we treated systems with0.2ppm of ClOH after5days of growth in the absence of disinfectant.Whole-cell insitu hybridizations usingfluorescently tagged,16S rRNA-targeted oligonucleotide probes performed on cryo-sectioned biofilms permitted the direct observation of metabolically active bacterial populations,includingcertain phylogenetic groups and species.The results of these studies confirmed the resistance of establishedbacterial biofilms to treatment with recommended levels of disinfectants.Specifically,Legionella pneumophila,E.coli,andand␦proteobacteria were identified within biofilms both before and after treatment.Further-more,although it was undetected using routine monitoring techniques,the observation of rRNA-containingE.coli within biofilms demonstrated not only survival but also metabolic activity of this organism within themodel distribution systems.The persistence of diverse bacterial species within disinfectant-treated biofilmssuggests that current testing practices underestimate the risk to immunocompromised individuals of contract-ing waterborne disease.Assessment of the microbiological safety of drinking water is based largely on the routine monitoring of water supplies for the presence of total coliforms and Escherichia coli.Detection of E.coli is considered indicative of recent fecal contamination and of the potential presence of enteric pathogens,while the presence of total coliforms is indicative of poor water quality. Whereas routine water quality measurements assess the presence of planktonic bacteria,the vast majority of bacteria indigenous to aquatic environments exist attached to solid par-ticles or surfaces.Within water distribution systems,significant bacterial populations exist as complex,structurally heteroge-neous biofilms attached to pipe surfaces.Residence within these complex matrices provides organisms with higher local-ized nutrient concentrations than are commonly found in drinking waters(10,14,18,22),and recent studies have shown that attached bacteria are more metabolically active than are their free-living counterparts(26,29).Furthermore,biofilms afford bacteria significant protection from disinfecting agents (3,31,35),including hypochlorous acid and monochloramine. Biofilms are dynamic in nature,and portions are frequently sloughed off pipe surfaces for a variety of reasons.They can then provide an effective inoculum for previously disinfected waters.The ability of total coliforms and E.coli to survive in biofilms is of marked significance to the water treatment industry,not only because the detection of these organisms in distribution water gives an incorrect indication of recent fecal contamina-tion but also because the persistence of these organisms and their release from biofilms may mask true breakthrough events in water treatment.Several lines of evidence suggest the long-term survival of total coliforms and/or E.coli in distribution system biofilms.Among these are the recovery of total coli-forms from distribution systems in which water leaving the treatment plant tested negative for coliforms(20,21);the observation that environmentally derived strains of total coli-forms and E.coli can grow in unsupplemented distribution system water(7);and the recovery of increased numbers of E. coli organisms from drinking water distribution pilot plants after the calculated inoculum should have theoretically washed out(12).Other studies have shown that E.coli can attach and become incorporated into biofilms within model distribution systems(8,11),as well as into biofilms derived from ground-water populations(2).In recent years,the advent of molecular detection tech-niques has greatly facilitated our understanding of bacteria indigenous to the environment.Whole-cell in situ hybridiza-tions usingfluorescently labeled oligonucleotides targeted to regions of the16S rRNA molecule have permitted the identi-fication of environmental bacteria with little disturbance of their surroundings(16,29,30).Because the ribosomal content of bacteria can be correlated with the growth rate,metabolic activity in environmental bacteria can be assessed by measur-ingfluorescence after hybridization(32).Whole-cell in situ hybridizations have been used for the direct identification ofand␥proteobacteria in biofilms formed on glass slides in*Corresponding author.Mailing address:Wadsworth Center,New York State Department of Health,P.O.Box509,Albany,NY12201. Phone:(518)473-7925.Fax:(518)402-5683.E-mail:bhowland@.†Present address:U.S.Environmental Protection Agency,NRMRL/ WSWRD/MCCB,Cincinnati,OH45268.5463 on September 17, 2015 by INSTITUTE OF URBAN ENVIRONMENT,CAS / Downloaded fromdrinking water (26)and for the examination of bio film struc-ture using embedded bio film sections (36,37,39).The studies reported here used whole-cell in situ hybridiza-tion on bio film sections obtained from model distribution sys-tems to assess the survival and growth capabilities of E.coli within multispecies bio films after exposure to chlorine disin-fectants.The responses of other organisms indigenous to water distribution systems,including Legionella pneumophila ,sulfate-reducing bacteria (SRB)belonging to the ␦subgroup of the proteobacteria,and the proteobacteria,were also deter-mined.MATERIALS AND METHODSModel distribution systems.A 50-liter polypropylene carboy (Nalgene Nunc International,Rochester,N.Y.),acting as a primary reservoir,supplied irrigant to the model systems through a series of Tygon tubing,Y-connectors,and pipettes.Up to six 1-liter Wheaton bottles were used in parallel as secondary reservoirs.The system flow rate was controlled by a peristaltic pump (Master flex L/S [Cole Parmer,Inc.,Vernon Hills,Ill.]or Rabbit peristaltic pump [Rainin Instrument Co.,Inc.,Woburn,Mass.])at 0.2ml/min or a velocity of 1.2mm/s.Two injection ports,located between the peristaltic pump and flow chamber,were constructed by filling the center opening of a T connector with silicone adhesive.Flow chambers (5)comprised a glass slide (76by 25by 1.5mm),two coverslips (50by 22mm,no.1thickness),and a ductile iron coupon (55by 18by 1.4mm [BioSurface Technologies,Inc.,Bozeman,Mont.]).Glass components were cleaned with acid-alcohol and ductile iron coupons were cleaned with 95%ethanol prior to assembly.The chambers were sealed using silicone adhesive and sterilized by baking at 160°C for at least 4h.Silicone tubing (inner diameter,1.02mm)provided in flow and out flow to and from the chambers,which had an approximate volume of 0.2ml (0.15by 0.3by 4.5cm).Figure 1presents a diagram of the model systems used in these studies.Flow chambers were irrigated at room temperature (RT)with up to 30liters of autoclaved tap water buffered with 0.54mM phosphate (0.48mM NaH 2PO 4,0.06mM Na 2HPO 4[pH 7.0to 7.5];Sigma Chemical Co.,St.Louis,Mo.)for 2weeks on a flat surface in the dark.The city of Albany,N.Y.,maintains distri-bution water at pH 8.5for corrosion control.To increase the ef ficacy of chlorine disinfectant in a consistent manner,the pH within the systems was maintained between 7.0and 7.5.The initial sterility of each model system was ascertained by plating sub-samples collected from the waste reservoirs onto R2A agar (Difco Laboratories,Detroit,Mich.)(34)and incubating them at RT for 10days.Subsamples were also fixed with 3.7%formaldehyde and examined microscopically after fluores-cent whole-cell hybridizations.Inoculation of model systems.To concentrate bacteria indigenous to water distribution systems,we filtered 5liters of tap water onto sterile,prepackaged Millipore GS membrane filters (pore size,0.2m [Millipore Corp.,Bedford,Mass.]).Filters were placed in a sterile 50-ml disposable centrifuge tube con-taining 10ml of tap water adjusted to pH 7to 7.5with 0.5mM phosphate buffer and vortexed for 1min to resuspend the particulate material.Flow chambers were slowly inoculated with 1ml of tap water particulate by injection through surface-sterilized injection ports.Inoculation of flow chambers with distribution system concentrates was typically repeated on each of the first 5to 7days of each 2-week incubation period.For experiments investigating the incorporation of E.coli into bio films,strain 01571was grown overnight in brain heart infusion (BHI)broth (Difco Labora-tories,Detroit,Mich.)at 35°C.To acclimate the E.coli to the low-nutrient conditions within the flow chambers,2ml of log-phase culture was centrifuged at 11,750ϫg for 5min.The resulting pellet was resuspended in autoclaved tap water,inoculated into approximately 50ml of autoclaved tap water,and incu-bated overnight at RT with shaking.On the following day,for each chamber,1ml of preconditioned E.coli was centrifuged at 11,750ϫg for 5min and resuspended in 1ml of tap water containing the concentrated distribution system bacteria described above.A similarly prepared mixture of distribution system populations and E.coli was injected into the flow chambers through the injection port on four successive days.Treatment with disinfectants.Secondary reservoirs (Wheaton bottles)were filled with 1liter of buffered tap water before the addition of either hypochlorous acid (ClOH;The Clorox Co.,Oakland,Calif.)or monochloramine (NH 2Cl).ClOH and NH 2Cl concentrations in stock solutions were determined by the N ,N -diethyl-p -phenylenediamine (DPD)ferrous titrimetric method (1)with one modi fication,namely,that HgCl 2was omitted from the phosphate buffer due to its toxicity (9).A scaled-down version of the titration method was used to test flow chamber discharge by addition of 5ml of wastewater to a flask containing 250l each of phosphate buffer and DPD indicator solution.This mixture was titrated with a 1/20dilution of ferrous ammonium sulfate titrant.Measurements were obtained immediately after collection of thesamples.FIG.1.Diagram of reservoirs and flow chambers assembled for the growth of bio films formed by water distribution system populations.5464WILLIAMS AND BRAUN-HOWLAND A PPL .E NVIRON .M ICROBIOL .on September 17, 2015 by INSTITUTE OF URBAN ENVIRONMENT,CAS/Downloaded fromNH2Cl was produced by reacting ammonium chloride(Sigma Chemical Co.) and hypochlorite ion(OClϪ)in a3:1molar ratio at pH10(17).Concentrations in stock solutions were determined immediately prior to each experiment. Enumeration of bacteria indigenous to tap water.To estimate the number of culturable heterotrophic bacteria present in tap water concentrates,we serially diluted subsamples of each suspension10-fold in buffered dilution water(1), spread plated them onto R2A agar,and incubated them for10days at RT before counting the colonies.For determination of the number of rRNA-containing organisms,a second subsample of each concentrate wasfixed using3.7%form-aldehyde and stored at4°C for subsequent use in the whole-cell hybridization procedure.Tap water was analyzed for the presence of fecal coliforms using membrane filtration onto m-FC agar(1).On occasion,total coliforms and E.coli were assayed in the chamber outflows using the Colilert method in the Quanti-Tray format(IDEXX Laboratories,Inc.,Westbrook,Maine).Cryoembedding and sectioning.Biofilms were embedded withinflow cham-bers by using Tissue-Tek O.C.T.embedding compound(Sakura Finetek U.S.A. Inc.,Torrance,Calif.),frozen on dry ice,and stored atϪ80°C(39).Before sectioning of the biofilms theflow chambers were snapped apart to release the embedded biofilm.Cross-sections(6m thick)of embedded biofilm were obtained using a cry-ostat(Miles,Inc.,Elkhart,Ind.).The sections were collected on baked Teflon-welled slides(14-mm-diameter wells[Erie Scientific,Portsmouth,N.H.])that had been treated with0.1%poly-L-lysine(Sigma)for5min at RT. Oligonucleotide probe synthesis and labeling.Oligodeoxynucleotide probes having a six-carbon linker containing a free amino terminus(Aminolink2[Ap-plied Biosystems,Foster City,Calif.])attached to the5Јend were synthesized by the Molecular Genetics Core of the Wadsworth Center using a DNA synthesizer (model8909;Perceptive Biosystems,Framingham,Mass.).Probes were de-blocked and stored atϪ20°C until needed for labeling reactions.Methods for labeling probes with tetramethyl rhodamine-5(and6)-isothiocyanate(TRITC; Molecular Probes,Inc.,Eugene,Oreg.)have been described previously(4).The probes used in these studies were as follows:primer342,5Ј-CTG CTG CSY CCC GTAG(38);Beta,5Ј-TCA CTG CTA CAC GYG(positions680to694);Delta, 5Ј-CGY GCG CCR CTY TACT(positions90to105);Leg5,5Ј-ACC GGA AAT TCC ACT ACC(positions667–684);Leg8,5Ј-GCT GCG CCA CTA ATT ATT T(positions845to863);and Eco3,5Ј-ACT TTA CTC CCT TCC TCC CCG (positions443to463).Primer342is a universal probe,whereas Beta and Delta are specific for some members of the beta-and delta-proteobacteria.Leg5and Leg8are specific for Legionella spp.,and Eco3allowed identification of E.coli. Probe specificity was empirically determined using the Ribosomal Database Project(25).Bacterial cultures and growth conditions.The bacterial strains used in these studies included strains Acinetobacter calcoaceticus ATCC23055,Agrobacterium radiobacter ATCC19358,Comamonas testosteroni ATCC11996,Salmonella en-terica serovar Typhimurium ATCC14028,Enterobacter aerogenes ATCC33457 and13048,Enterobacter cloacae ATCC33457,Escherichia coli ATCC43651, 25922and795,E.fergusonii ATCC35469,E.vulneris ATCC33821,E.blattae ATCC29907,and E.hermannii ATCC33650.Aureobacterium sp.,Arthrobacter globiformis,Bacillus subtilis,Cytophaga pectinovorans,and Pseudomonasfluores-cens were provided by S.Nierzwicki-Bauer,at Rensselaer Polytechnic Institute (Troy,N.Y.).Thames Water Utilities(Thames,United Kingdom)supplied47 E.coli isolates,as well as77other coliforms,including several isolates each of Klebsiella terrigena,K.ornithinolytica,K.oxytoca,K.pneumoniae,K.ascorbata, Citrobacter freundii,Enterobacter aerogenes,E.sakazakii,E.intermedium,E.cloa-cae,E.agglomerans,E.amnigenus,E.taylorae,and Aeromonas schubertii.Desul-fovibrio desulfuricans was provided by M.Wolin of the Wadsworth Center, Albany,N.Y.Legionella pneumophila isolates1164,1169,and1818were pro-vided by D.Schoonmaker-Bopp,also of the Wadsworth Center.E.coli01571was isolated from potable water collected in Steuben County,N.Y.,in July1998.The majority of cultures were maintained on standard plate count agar at4°C and transferred monthly.For whole-cell hybridizations,bacteria were grown in BHI broth at RT or37°C.Log-phase cultures werefixed in3.7%formaldehyde and prepared for the hybridization procedure.L.pneumophila isolates were grown on Legionella agar base medium(Difco Laboratories)supplemented with0.2g of L-cysteine-HCl and0.125g of ferric pyrophosphate per liter and adjusted to pH 7.1to7.2.Cultures were incubated at37°C in a candle extinction jar(13).D. desulfuricans was grown anaerobically at37°C in serum bottles containing LSTY broth reduced with0.05%dithiothreitol(24).D.desulfuricans was transferred to new medium in serum bottles by using a sterile syringe and needle prereduced using1.25%cysteine sulfide.Fluorescent in situ hybridizations.16S rRNA-targeted oligodeoxynucleotide probes were tested for specificity by hybridization to the laboratory cultures described above.Fixed bacterial cultures were washed and prepared on gelatin-coated slides as described previously(4).Since L.pneumophila isolates were grown on agar medium instead of broth,an inoculating loopful of organisms from an isolated colony was placed in1ml of phosphate-buffered saline(PBS) containing3.7%formaldehyde and mixed well.Well slides containing biofilm sections were held for1h at RT in Coplin jars containing PBS(pH7.4)(Sigma Chemical Co.)with3.7%formaldehyde and then dehydrated by serial exposure to50,80,and100%ethanol for3min each (32).The slides were then treated for1min in90:10ethanol-formaldehyde and given a5-min wash infiltered(0.2-m pore size)and autoclaved distilled H2O. Hybridizations using the universal342,Beta,or Delta probes were carried out at37°C as described previously(4),with either5g of4Ј,6-diamidino-2-phe-nylindole dihydrochloride(DAPI)(Molecular Probes,Inc.)per ml or2.5g of 7-diethylamino-3-4Ј-isothiocyanatophenyl)-4-methylcoumarin(CPI)(Molecular Probes,Inc.)per ml as a total-cell counterstain.The Eco3probe,which has a T m of63°C,was resuspended in a mixture of400l of formamide and600l of hybridization mix for subsequent overnight hybridization at37°C.For biofilm sections,the wash times were increased to three30-min washes in1ϫSET. Planktonic distribution system populations or laboratory cultures underwent the same hybridization procedure,with the omission of the1-h formaldehyde incubation and serial ethanol washes.To confirm the presence of Legionella species in the biofilms,fluorescent-antibody staining was performed on biofilm cryosections by usingfluorescein-labeled goat anti-L.pneumophila serogroup1to14immunoglobulin G and goat anti-Legionella species b to p immunoglobulin G(Monoclonal Technologies, Inc.,Atlanta,Ga.)after hybridization with Leg5and Leg8.After the third30-min wash in1ϫSET,the slides were air dried.The slide well was covered with antibody solution(Monoclonal Technologies,Inc.)containing1%whole-goat serum(Organon Teknika Corp.,West Chester,Pa.)and incubated at RT in the dark for30min in a box containing a moistened Kim Wipe.The slide was washed twice for5min each in a Coplin jar containing PBS.Epifluorescence microscopy.The results of hybridizations were examined us-ing a Leitz DMR-B epifluorescence microscope(Leica Mikroskopie and System GmbH)equipped with a100-W power supply,phase andfilter sets appropriate for rhodamine(no.513810[Leica Inc.,Deerfield,Ill.]),fluorescein(no.513812), and DAPI(or CPI)(no.513808)detection.Bacteria were viewed through a 100ϫ/1.3Plan Fluotar oil immersion objective.Photomicrographs were obtained using a Nikon UFX-DX automatic camera and Fujichrome200-speed color slide film.Effects of chlorine on whole-cell hybridization.To determine the ability of the whole-cell hybridization procedure to detect the effects of chlorination on met-abolically active(rRNA-containing)cells,chlorine was added to pure cultures of E.coli and bacteria obtained from distribution water concentrates and micro-scopically evaluated over time.E.coli(Thames Water Utilities isolate159), grown to log phase in BHI broth and washed twice in sterile water,and bacteria obtained from distribution water concentrates were exposed to1ppm of ClOH. Subsamples were removed at0,10,and30min and immediately placed in sodium thiosulfate(Na2S2O3;final concentration0.01%)before undergoing the hybrid-ization procedure with the universal342probe.Control(untreated)samples of E.coli and distribution water bacteria were analyzed in parallel.RESULTSSpecificity of oligonucleotide probes.All E.coli strains,in-cluding ATCC cultures and environmental isolates,were de-tected by epifluorescence microscopy after hybridization with the Eco3probe.Of the77other coliform strains used as neg-ative controls,only two environmental strains of K.oxytoca hybridized with this probe.Biochemical identification of these strains was confirmed using API test strips(bioMe´rieux,Inc., Durham,N.C.).The Leg5,Leg8,Delta,and Beta probes hy-bridized with only the appropriate positive controls.The uni-versal probe342hybridized with the16S rRNA of all bacterial isolates and type strains tested.Effects of chlorine on whole-cell hybridization.The ability of the whole-cell hybridization procedure to measure the effects of chlorination on bacterial metabolic activity was tested on laboratory-grown and distribution system bacteria.As indi-cated in Table1,the hybridization procedure has the sensitivityV OL.69,2003GROWTH OF E.COLI IN BIOFILMS5465on September 17, 2015 by INSTITUTE OF URBAN ENVIRONMENT,CAS /Downloaded fromto measure the effect of1ppm of ClOH on bacteria within10 min of exposure.Log-phase,laboratory-grown E.coli cells, which showed a2-log-unit reduction in the numbers of bacteria detected,were more sensitive to chlorine treatment than were unamended bacterial populations freshly obtained from the distribution system.Metabolic activity within distribution system biofilms.The results offluorescent in situ hybridizations indicated that within distribution water concentrates,an average of30.72% (nϭ23,where n is the number of concentrated water samples examined,with a minimum of200bacteria enumerated per sample)of the bacterial cells staining with CPI also contained detectable amounts of rRNA.Percentages were not seasonal, ranging from a low of7.94%in April1999to a high of55.68% in April1998.Each milliliter of distribution water concen-trates,freshly prepared in an identical manner daily,contained an average of6.7ϫ106cells,based on microscopic enumera-tion.Determining viability from CFU on R2A plates indicated that approximately27.5%of total distribution water bacteria were culturable under the conditions used.In contrast,in situ hybridizations performed on cross-sections of distribution sys-tem biofilms obtained fromflow chambers after a2-week in-cubation period indicated that many bacteria contained detect-able levels of rRNA after hybridization with the universal342 probe(Fig.2A and B).Biofilms were structurally heteroge-neous,exhibiting pillars and interstitial channels.However, position within the biofilm,e.g.,proximity to either the sub-strate coupon or waterflow,had no apparent effect on the percentage of cells detected.Likewise,treatment with a variety of disinfectant regimens had no effect on the percentages of cells observed.Treatment regimens included exposure of bio-films to(i)1ppm of free chlorine for67min(1CT),(ii)a residual of0.2ppm of free chlorine after the biofilms grew for 5days without disinfectant,and(iii)4ppm of monochloramine for155min(Table2).However,whenflow chambers were treated continuously with a2ppm of NH2Cl,4ppm of NH2Cl, or0.2ppm of ClOH residual for2weeks,biofilm was not detected after embedding and sectioning.Presence of SRB in biofilms.Bacteria that were detected after hybridization with the Delta probe were considered to be SRB.SRB were present in significant quantities within each biofilm,even after exposure to free chlorine or monochlora-mine.In biofilm1,the majority of SRB detected were large, short rods(Fig.2D).In biofilms4and7to10,the cells were smaller than those previously detected,although numerous SRB were observed.In most sections,SRB were scattered throughout the biofilm structure,including near the interface between the biofilm and waterflow.L.pneumophila in biofilms.L.pneumophila and other Le-gionella species were detected in the treated and untreated biofilms7to10by using whole-cell in situ hybridizations(Fig. 2F)andfluorescent antibodies(data not shown).Because the Leg5and Leg8group probes hybridize with several Legionella species,the presence of L.pneumophila and nonpneumophila Legionella spp.was confirmed by usingfluorescent antibodies. More bacteria stained with the antibodies than hybridized with the oligonucleotide probes.In these experiments,total bacteria were detected after staining with DAPI instead of CPI(Fig. 2E).Survival and growth of E.coli in biofilms.The ability of E. coli to survive in model distribution biofilms was examined using fluorescent in situ hybridizations performed on cross sections of biofilms obtained fromflow chambers into which an environmen-tal strain of E.coli had been inoculated.The results of whole-cell hybridizations with the universal342probe indicated that the total number of E.coli cells inoculated into eachflow chamber was approximately106,70%of which were detected after hybrid-ization and subsequently considered metabolically active.Mix-tures of distribution water populations and E.coli injected into theflow chambers each day contained approximately3%E.coli. As shown in Fig.3,E.coli cells were detected within2-week-old biofilms seeded with the environmental isolate.Similar to the results of studies using the universal probe,E.coli was detected in a seeded control(untreated)biofilm as well as in two seeded biofilms exposed to1CT free chlorine(Fig.3B)and one biofilm exposed to10CT monochloramine(Fig.3D).Since biofilms were embedded10days after thefinal inoculation of E.coli into the systems,E.coli is clearly able to survive for at least10days within a biofilm.Fecal coliforms were not detected in116distribution system water samples(11.6liters total tested,obtained between No-vember1997and July1999)using membranefiltration onto m-FC agar.However,a biofilm developed solely from distri-bution system populations contained bacteria that were de-tected after hybridization with the Eco3probe(Fig.3F)and confirmed by standard culturing methods.These indigenous E. coli cells were easily differentiated morphometrically from E. coli01571grown in the laboratory and seeded into biofilms in the experiments described above.The indigenous cells were lessfluorescent and much smaller,suggesting that they had been exposed to adverse conditions for some time.The ability of bacteriological tests used in the routine monitor-ing of potable water supplies to detect E.coli sloughing from biofilms was examined.Table3presents the occurrence of E.coli in chamber outflow samples using the Colilert Quanti-Tray method,beginning1day after thefinal inoculation of bacteria intoflow chamber3and7days afterfinal inoculation intoflow chambers7,8,10,11,and12.E.coli was detected in biofilm3on two occasions during the incubation period,although only bacte-rial concentrates obtained from distribution water were used as inocula.Incubation of subsamples of the E.coli-positive medium overnight on LES-Endo agar at35°C and m-FC agar at44.5°C gave rise to growth with a green metallic sheen and to blue colonies,respectively.In particular,the blue growth on m-FC agar at44.5°C indicated the presence of fecal coliforms in theTABLE1.Effects of1ppm of ClOH on the hybridization efficiency of Universal342probe to16S rRNA in planktonic bacteria aTime (min)%of cells positive after hybridization cE.coli control E.coli,1ppm of ClOHTW bcontrolTW,1ppm of ClOH0108.5Ϯ2.12104.5Ϯ2.1220.7Ϯ15.331.7Ϯ7.1410103.5Ϯ0.7070.63Ϯ0.89125.8Ϯ11.5 4.25Ϯ4.603098.2Ϯ0.848 1.69Ϯ2.3939.9Ϯ22.3 4.32Ϯ2.66a The E.coli isolate,obtained from Thames Water Utilities,was grown to logphase in BHI broth and washed twice prior to treatment.The distribution systembacteria were treated and analyzed directly after they were sampled.b TW,bacteria present in tap waterfilter concentrates.c Two experiments were averaged,and the meanϮstandard deviation isshown.5466WILLIAMS AND BRAUN-HOWLAND A PPL.E NVIRON.M ICROBIOL.on September 17, 2015 by INSTITUTE OF URBAN ENVIRONMENT,CAS /Downloaded from。
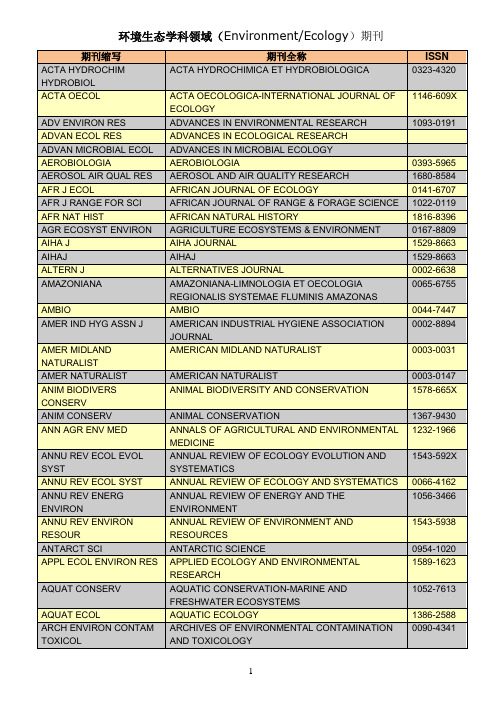
Environment Ecology领域期刊清单
ISSN
0003-9896 0324-8461 1523-0430 0004-0843 0004-1254 1532-4982 0890-3069 0307-692X 0004-9573 1442-9985 1448-6563 1439-1791 0276-5055 0960-3115 1939-1234 0168-2563 1726-4170 0006-3207 0178-2762 0961-9534 0895-3988 1088-9868 0006-3606 0959-2709 1239-6095 0007-4861 0008-3550 0008-4271 0701-1784 0341-8162 1612-1872 0275-7540 0954-2299 0937-7409 0045-6535 1618-954X 1863-0650 1469-3062 0936-577X 0892-0753 0010-3624
期刊全称
ACTA HYDROCHIMICA ET HYDROBIOLOGICA ACTA OECOLOGICA-INTERNATIONAL JOURNAL OF ECOLOGY ADVANCES IN ENVIRONMENTAL RESEARCH ADVANCES IN ECOLOGICAL RESEARCH ADVANCES IN MICROBIAL ECOLOGY AEROBIOLOGIA AEROSOL AND AIR QUALITY RESEARCH AFRICAN JOURNAL OF ECOLOGY AFRICAN JOURNAL OF RANGE & FORAGE SCIENCE AFRICAN NATURAL HISTORY AGRICULTURE ECOSYSTEMS & ENVIRONMENT AIHA JOURNAL AIHAJ ALTERNATIVES JOURNAL AMAZONIANA-LIMNOLOGIA ET OECOLOGIA REGIONALIS SYSTEMAE FLUMINIS AMAZONAS AMBIO AMERICAN INDUSTRIAL HYGIENE ASSOCIATION JOURNAL AMERICAN MIDLAND NATURALIST AMERICAN NATURALIST ANIMAL BIODIVERSITY AND CONSERVATION ANIMAL CONSERVATION ANNALS OF AGRICULTURAL AND ENVIRONMENTAL MEDICINE ANNUAL REVIEW OF ECOLOGY EVOLUTION AND SYSTEMATICS ANNUAL REVIEW OF ECOLOGY AND SYSTEMATICS ANNUAL REVIEW OF ENERGY AND THE ENVIRONMENT ANNUAL REVIEW OF ENVIRONMENT AND RESOURCES ANTARCTIC SCIENCE APPLIED ECOLOGY AND ENVIRONMENTAL RESEARCH AQUATIC CONSERVATION-MARINE AND FRESHWATER ECOSYSTEMS AQUATIC ECOLOGY ARCHIVES OF ENVIRONMENTAL CONTAMINATION AND TOXICOLOGY 1
厌氧氨氧化

Journal of Hazardous Materials 250–251 (2013) 431–438Contents lists available at SciVerse ScienceDirectJournal of HazardousMaterialsj o u r n a l h o m e p a g e :w w w.e l s e v i e r.c o m /l o c a t e /j h a z m atDenitrification performance and microbial diversity in a packed-bed bioreactor using biodegradable polymer as carbon source and biofilm supportZhiqiang Shen a ,b ,Yuexi Zhou a ,Jun Hu b ,Jianlong Wang b ,c ,∗aChinese Research Academy of Environmental Sciences,Beijing 100012,PR ChinabLaboratory of Environmental Technology,INET,Tsinghua University,Beijing 100084,PR China cBeijing Key Laboratory of Fine Ceramics,Tsinghua University,Beijing 100084,PR Chinah i g h l i g h t sStarch/PCL (SPCL)blend was prepared and used for biological denitrification.The microbial community of attached biofilm was analyzsed by metagenomic method. The vast majority of species biofilm (99.71%)belonged to six major phyla. Proteobacteria were the most abundant phylum (85.50%).Diaphorobacter and Acidovorax was 52.75%of identified denitrifying bacteria.a r t i c l ei n f oArticle history:Received 15October 2012Received in revised form 10February 2013Accepted 13February 2013Available online 20 February 2013Keywords:Nitrate BiofilmMicrobial community DenitrificationSolid carbon sourcea b s t r a c tA novel kind of biodegradable polymer,i.e.,starch/polycaprolactone (SPCL)was prepared and used as carbon source and biofilm support for biological denitrification in a packed-bed bioreactor.The deni-trification performances and microbial diversity of biofilm under different operating conditions were investigated.The results showed that the average denitrification rate was 0.64±0.06kg N/(m 3d),and NH 3–N formation (below 1mg/L)was observed during denitrification.The nitrate removal efficiency at 15◦C was only 55.06%of that at 25◦C.An initial excess release of DOC could be caused by rapid biodegra-dation of starch in the surfaces of SPCL granules,then it decreased to 10.08mg/L.The vast majority of species on SPCL biofilm sample (99.71%)belonged to six major phyla:Proteobacteria,Bacteroidetes,Chloroflexi,Firmicutes,Spirochaetes and Actinobacteria.Proteobacteria were the most abundant phylum (85.50%)and mainly consisted of -proteobacteria (82.39%).Diaphorobacter and Acidovorax constituted 52.75%of the identified genera which were denitrifying bacteria.© 2013 Elsevier B.V. All rights reserved.1.IntroductionThe “solid-phase denitrification”is a new type of heterotrophic biological denitrification in which insoluble biodegradable poly-mers were used as biofilm carrier and carbon source simulta-neously [1,2].Solid substrates were used as alternatives to liquid carbon sources,which are accessible only by microbial enzymatic attack,so it can avoid the risk of overdosing in liquid carbon sources supported denitrification system [1].In the past few years,solid substrates have been investigated as a carbon source in the biolog-ical denitrification of drinking water [3,4],groundwater [5],landfill∗Corresponding author at:Laboratory of Environmental Technology,INET,Tsinghua University,Beijing 100084,China.Tel.:+861062784843;fax:+861062771150.E-mail addresses:wangjl@ ,wangjl@ (J.Wang).leachate [6,7]and recirculated aquaculture system [1,8].Water body is susceptible to the over use of fertilizers and pesticides in agriculture fields,then the simultaneous removal of nitrate and pesticides has been studied using solid substrates as carbon sources and biofilm carrier [9–11].Furthermore,some researchers have also studied the feasibility of using biodegradable compounds as carbon sources and biofilm carrier for nitrogen removal in simul-taneous nitrification and denitrification (SND)system [12–14].There are two kinds of solid carbon sources which have been investigated for denitrification,synthetic polymers and natural materials especially the cellulose-rich monly,syn-thetic polymers are thermoplastic which are easy processed to various shapes to fit the demand of different denitrification process.In the “solid-phase denitrification”system,synthetic polymers including polyhydroxyalkanoates (PHAs)[2]and polycaprolactone (PCL)[1,8,15]were prepared to granules for denitrification,mean-while,PCL also can be prepared to plates for use [16].Comparing with the expensive synthetic polymers,natural materials including0304-3894/$–see front matter © 2013 Elsevier B.V. All rights reserved./10.1016/j.jhazmat.2013.02.026432Z.Shen et al./Journal of Hazardous Materials250–251 (2013) 431–438 wheat straw[10,17],cotton[18],waste newspaper[19],pine bark[6],crab-shell chitin[5]were much cheaper but may bring ammo-nia[5],high DOC and color problems in effluent[3].Therefore,thekey issue of“solid-phase denitrification”is to develop new solidsubstrates with low denitrification cost and without deteriorationof effluent water quality.Blending with some cheap organic materials is a mostpotential method to lower the price of products especiallyin biodegradable plastics productionfiparing withother biodegradable thermoplastic polymers,starch is an abun-dant renewable polysaccharide with better biodegradability andlow cost.Aliphatic polyesters are biodegradable thermoplasticpolymers with good processability,thermal stability,excellentmechanical properties,good water resistance,and dimensional sta-bility[20].So it is a potential way to blend starch with aliphaticpolyesters for production of biodegradable plastics.Aliphaticpolyesters such as PCL[21–23],poly(butylene succinate)(PBS)[24],poly(hydroxybutyrate-covalerate)(PHBV)[25]and poly(lacticacid)(PLA)[26,27]were widely adopted to blend with starch forbiodegradable plastics production or medical application.Recently,we investigated the feasibility of using cross-linkedstarch/polycaprolactone blends as solid carbon source and biofilmcarrier for denitrification[28].The main objective of this study was:(1)to evaluate the denitrification ability of starch/polycaprolactone(SPCL)blends serving both as carbon source and biofilm carrier;(2)to assess the effect of operating conditions(i.e.nitrate loading rates,temperature and initial pH)on denitrification under continuous-mode;(3)to analyze the microbial community structure of biofilmattached on SPCL.2.Materials and methods2.1.MaterialsThe polycaprolactone(PCL)used in this study has a molecularweight of60,000g/mol(Dalton).The cornstarch used in this studyis technical grade.Starch/PCL(SPCL)blends were prepared by twin-screw extruder.The main characters of SPCL are listed as follows:Starch,55.44%;PCL,30.00%;Additives(plasticizer and couplingagent),14.56%;Calculated surface area,1833.33m2/m3.2.2.Experiment apparatusContinuous experiments were carried out in a laboratory-scalepacked-bed set-up(Fig.1)using50mm inner diameter by500mmheight cylindrical Plexiglas.A Plexiglas mesh disc(48mm diam-eter,2mm pore size)was placed at the lower end of the columnas support for the packing material.SPCL granules were usedasFig.1.Schematic display of the experimental set-up.carbon source and carrier for biofilm growth,and packing the col-umn up to a height of250mm(273g).2.3.Experiment proceduresSPCL granules were used as biofilm carrier and electron donor to support biological denitrification.Synthetic water(adding NaNO3 and KH2PO4to tap water,and N:P(w/w)=5:1)seeded with acti-vated sludge which collected from a local municipal wastewater treatment plant(with thefinal concentration of800mg/L MLSS) was pumped into the bottom of column at aflow rate of4.1mL/min (HRT=2h).Unless otherwise indicated,temperature was25±1◦C, and pH and DO were not controlled.After a stable denitrification performance obtained,flow rate was then increased stepwise to study the effect of nitrate loading rates(lasted184d).Then,deni-trification performance at low temperature(15±1◦C)was studied (lasted20d).The effect of pH on denitrification was investigated using1M hydrochloric acid and sodium hydroxide to control the pH of synthetic water to4.5,6,9and10.5(lasted74d).Samples were taken from the effluent to monitor NO3–N,NO2–N,NH3–N, pH and DOC.2.4.Analytical methodsSamples were taken andfiltered through0.45m membrane before analysis.NO3–N was determined by UV-spectrophotometer (Shimadzu UV-3100)at220nm and275nm,and NO2–N and NH3–N were assayed by hydrochloric acid naphthyl ethylenedi-amine spectrophotometry method and hypochlorite-salicylic acid spectrophotometry method,respectively[29].Dissolved organic carbon(DOC)was detected using a TOC analyzer(HACH,IL530TOC-TN).Samples were neither acidified nor sparged during analysis. The pH value was measured with pH meter.The morphology of the samples was examined using a SEM(Fei Quanta200).Biofilm was taken from the column reactor after being operated for280d.DNA from biofilm was extracted directly using glass beads to mechanical lysis of SPCL granulars(with biofilm).The volumetric denitrification rate R vd in kg/(m3d) NO3–N+NO2–N(total soluble oxidized nitrogen species)of the reactor is given by the Eq.(1):R vd=0.024×Q D×(C in−C ef)V(1) where C in is the influent NO3–N+NO2–N concentrations(mg/L)and C ef is the effluent NO3–N+NO2–N concentrations.Q D is theflow rate(L/h)and V is the reactor volume(L).Nitrate removal efficiency(N re)is defined by the equation(2): N re=100×(NO3−N in−NO3−N ef−NO2−N ef−NH3−N ef)NO3−N in(2) where NO3–N in is the influent NO3–N concentration,NO3–N ef, NO2–N ef and NH3–N ef are the effluent NO3–N,NO2–N and NH3–N concentrations,respectively(negligible accumulation of gaseous-N by-products and organic-N).2.5.Pyrosequencing2.5.1.DNA extraction and purificationGenomic DNA was extracted using E.Z.N.A.Soil DNA Kit (OMEGA).2.5.2.PCR amplificationFor each sample,we amplified V1-V3region of bacterial16S rRNA genes using a broadly conserved primer set(27F and533R). The forward primer(5 -GCC TTG CCA GCC CGC TCA GAG AGT TTGZ.Shen et al./Journal of Hazardous Materials250–251 (2013) 431–438433ATC CTG GCT CAG-3 )contained the454Life Sciences primer B sequence and the broadly conserved bacterial primer27F.The reverse primer(5 -GCC TCC CTC GCG CCA TCA GNN NNN NNN NNT TAC CGC GGC TGC TGG CAC-3 )contained the454Life Sci-ences primer A sequence,a unique10-nt barcode used to tag each PCR product(designated by NNNNNNNNNN),and the broad-range bacterial primer533R.PCR reactions were carried out in tripli-cate20-L reactions with0.4M forward and reverse primers, 1-L template DNA,250nM dNTP and1×FastPfu Buffer.All dilu-tions were carried out using certified DNA-free PCR water.Thermal cycling consisted of initial denaturation at95◦C for2min followed by25cycles of denaturation at95◦C for30s,annealing at55◦C for 30s,and extension at72◦C for30s,with afinal extension of5min at72◦C.Replicate amplicons were pooled and visualized on2.0% agarose gels using SYBR Safe DNA gel stain in0.5×TBE.Amplicons were purified using AxyPrep TM DNA Gel Extraction Kit(AXYGEN) according to the manufacturer’s instructions.2.5.3.Amplicon,quantitation,pooling and pyrosequencingAmplicon DNA concentrations were measured using the Quant-iT PicoGreen dsDNA reagent and kit(Invitrogen).DNA samples were diluted in30L1X TE,an equal volume2X PicoGreen working solution was added in a total reaction volume of60L in minicell cuvette.Fluorescence was measured on a Turner Biosystems TBS-380Fluorometer using the465-485/515-575-nm excitation/emissionfilter pair.Following quantitation,cleaned amplicons were combined in equimolar ratios into a single tube.Pyrosequencing was carried out on a454Life Sciences Genome Sequencer FLX Titanium instrument(Roche)by Shanghai Majorbio Bio-pharm Biotechnology Co.Ltd.(Shanghai,China),the sequenc-ing data was analyzed by Mothur[30].3.Results and discussion3.1.Effect of nitrate loading rates on denitrificationThe effect of nitrate loading rates on denitrification was studied varying the nitrate loading rates between0.60and1.20kg/(m3d) through changing the influent nitrate concentration and theflow rate(Figs.2and3,Table1).As the formation of biofilm on the surface of SPCL,nitrate removal efficiency increased gradually and reached98.42%at the7th d,and then it became stable during the period of days8–25(Fig.2).The lag time(=period of adaptation of denitrifying microorganisms)of PCL was16days in recirculated aquaculture systems[1].The lag time of SPCL was only7d,indi-cating that PCL blending with starch can significantly shorten the lag time in solid denitrification system.When the nitrate loading rate increased from0.60to1.20kg/(m3d)(at the25th d),the efflu-ent nitrate and nitrite concentrations increased significantly,and a very sharp decline of nitrate removal rate was observed simul-taneously(phase2).These changes may be due to the high nitrate loading rates which exceeded the denitrification capability of the system.Keeping a constantflow rate(8.2mL/min)and decreasing influent nitrate concentration to25mg/L at the60th d,the effluent nitrate and nitrite concentrations decreased sharply and ca.90% nitrate removal rate reached(phase3).Further increasing nitrate loading rate to0.72kg/(m3d)at the100th d,a littlefluctuationofFig.2.Denitrification performances of SPCL at different nitrate loading rates. denitrification performance was observed(phase4).Nitrite is an intermediate of nitrate reduction.In this study,nitrite concentra-tion was below1mg/L when high nitrate removal rate reached.The formation of NH3–N(below ca.1mg/L)was observed,possibly due to the dissimilatory nitrate reduction to ammonia(DNRA)process, which was also observed in anaerobic sediments by Kelso et al.[31]. Honda and Osawa[16]also found that a0.1mg/L NH3–N increased in denitrification system using PCL as substrate.Table1The volumetric denitrification rate R vd at different nitrate loading rates.Phase Operating date(d)Flow rate(mL/min)Influent NO3–N(mg/L)HRT(h)NO3–N loading rate(kg/(m3d))R vd(kg/(m3d)) 10–25 4.15020.600.53±0.09a226–598.2501 1.200.54±0.22360–1008.22510.600.59±0.03 4101–18416.4150.50.720.64±0.06a Mean±standard deviation.434Z.Shen et al./Journal of Hazardous Materials 250–251 (2013) 431–4380.60.8 1.0 1.20.60.81.01.2N i t r a t e r e m o v e d [k g /(m 3·d )]Nitrate load [kg/(m 3·d)]Fig.3.Average nitrate removal rates under different loading rates.The variations of DOC and pH in effluent are also depicted inFig.2.During the start-up period,a quick release of DOC occurred,and reached maximum 61.30mg/L at the 14th d.The high DOC might allow rapid microbial growth and the fast colonization on the substrate therefore high removal of nitrate was observed.DOC will accumulate when the amount of released dissolved organic carbon exceeded the need of microbes for both growth and denitrification.During the start-up period,once microbes stick and proliferate to SPCL granules,they secrete enzymes to biodegrade starch or PCL and use them as carbon source.Since starch is more biosuscep-tible than PCL in starch/PCL blends [21,32],so the accumulated of DOC may be mainly derived from the biodegradation of starch (on the surface of SPCL granules)during the start-up period.HRT may be an important factor affecting the release and accumulation of DOC.At the 24th d,the flow rate was changed to 8.2mL/min (HRT changed from 2h to 1h),the average DOC in the effluent decreased to 19.60mg/L (phases 2and 3).Further increasing the flow rate to 16.4mL/min (HRT =0.5h),the average DOC decreased to 10.08mg/L (phase 4).The previous work also found that HRT has an important influence on DOC accumulation using cross-linked starch/PCL blends as solid carbon and biofilm carrier for denitrifi-cation [28].Aslan and Türkman [3]also found that DOC decreased with increasing flow rate in fixed-bed denitrification system using wheat straw as substrate.Shear force has significant influences on the structure of the biofilm and mass transfer.A higher shear force may result in a thin-ner and denser biofilm [33],but it has a dual effect on the behaviors of mass transfer in biofilm,i.e.high turbulence would facilitate sub-strate diffusion in biofilms;however,shearforce-enhanced biofilm density in turn reduces the diffusivity of substrate in biofilms.The observed diffusivity of substrate would be a net result of these two phenomena [34].Celmer et al.[35]found that high shear force proved to be effective in improving denitrification rate by reducing the thickness of the biofiparing phases 1with 3(Table 1),under a constant nitrate loading rate (0.60kg/(m 3d))the average denitrification rate improved since flow rate increased from 4.1to 8.2mL/min.Thus,the need of carbon source for biological denitri-fication increased,which might be a main result of the decrease of DOC.DOC values did not exceed 5–7mg/L when using PCL as solid carbon source in recirculated aquaculture systems [1].In this study,DOC was higher than net PCL supported denitrification sys-tem,which probably due to the addition of starch and a high starch content (starch:PCL (w/w)=1.848)in the blends,meanwhile the difference of PCL may be another reason though it maybe play a less important role in the high DOC problem.The values of pH decreased slightly from a range of 6.89–7.87(influent)to a range of 6.47–7.48(effluent)in the period of days 26–184.The decrease of pH values probably be due to organic acids produced from carbon source by microbial metabolism,and neu-tralization alkalinity represented by denitrifiers [15].When PCL immersed in enzyme lipase solution,the quantity of acid liberated was coincided with its biodegradability [36].After immersed into lysozyme solution,the pH of media containing pure PCL scaffolds was lower than the initial pH due to the acidic degradation products of the PCL component [37].Increasing nitrate loading rate properly,the average denitrifica-tion rate increased (phases 3and 4)while over-loading of nitrate (phase 2)could not improve the denitrification performance (Fig.3and Table 1).Under the same nitrate loading rate,increasing the flow rate led to a higher average denitrification rate (phases 1and 3).3.2.Denitrification performance at low temperatureAt the 185th d,the temperature was decreased to 15◦C (except temperature,other operating parameters equaled to phase 4)to study the denitrification performance at low pared to phase 4at 25◦C (Fig.2),the effluent average nitrate concentrations increased significantly and reached 7.03±0.36mg/L,nitrate removal efficiency was 47.50%.Though nitrate removal was inhibited at low temperature,nitrite accu-mulation was low (below 0.6mg/L).The average denitrification rate was 0.34±0.01kg/(m 3d),indicating that temperature was an important parameter for denitrification performance.A very sharp decline of denitrification rate was also observed in wheat straw or cotton supported denitrification system when tempera-ture decreased [9,18].As denitrification rate decreased,the demand of carbon source for biological denitrification decreased.Meanwhile,the amount of released dissolved organic carbon should be reduced since the enzymatic degradation process of starch/PCL blends affected by temperature.PCL showed a slight biodegradability under aquatic conditions at the mesophilic temperature [38],but the biodegrad-ability of PCL was 92%in the diluted sludge at the thermophilic temperature [39].At 15◦C,average DOC was 4.59mg/L,which was only 45.54%of the value in phase 4at 25◦C.pH values slightly decreased from a range of 6.90–7.57(influent)to a range of 6.72–7.05(effluent).3.3.Effect of pH on denitrification performanceThe denitrification performance at different initial pH (4.5–10.5)was studied (except initial pH,other operating parameters equaled to phase 4).Compared with uncontrolled initial pH condition (phase 4,Fig.2),increase of effluent nitrate and decrease of nitrate removal rate were observed at both acid and basic condition,indi-cating that when pH was beyond the optimal range,denitrification enzymatic activity was inhibited.It was similar to the optimal pH range reported for denitrification [40,41].At acidic pH (4.5and 6),the average denitrification rates at pH of 4.5and 6were 0.46±0.05and 0.51±0.04kg/(m 3d),respectively.These values were significantly lower than the value in phase 4(average pH ca.7.23),which was 0.64±0.06kg/(m 3d)(Table 1).In addition,the effluent nitrite and DOC concentrations at pH of 4.5were higher than the values at pH of 6.It was interesting to note that the average denitrification rate increased from 0.35±0.03to 0.52±0.02kg/(m 3d)when pH increased from 9to 10.5,which might be due to the higher DOC release and accumulation at pH of 10.5than 9.At basic condition,more organic acid products were produced to neutralize the alkalinity,which confirmed by pH change.These excess release of dissolved organic carbon should beZ.Shen et al./Journal of Hazardous Materials250–251 (2013) 431–438435Fig.4.Photograph of fresh SPCL(a);SEM images of biofilm attached on SPCL(b);fresh SPCL(c);and used SPCL(d).exceeded the need of microbes for denitrification,so a higher DOC accumulation was observed at basic condition than acidic or neu-tral condition.Effluent pH tended to be neutral both at acidic and basic initial conditions,which would be a net result of acidity by acidic degradation products and alkalinity derived from denitrifi-cation.NH3–N was produced over the pH range of4.5–10.5,but the concentration was below1.00mg/L.3.4.SEM observationSPCL carriers used in this study were cylindrical granules (Fig.4a),and biofilm attached on their surfaces comprised predo-minately of rod bacteria from SEM observation(Fig.4b).The fresh SPCL displayed an irregular surface(Fig.4c),which will favor the attachment of bacterial cells on the surfaces.Generally,without deformed starch granules is homogeneously dispersed throughout the PCL/starch blends as droplet-like particles[22].The starch parti-cles present in SPCL showed a thermoplastic nature,indicating that it underwent significant paring the SEM images of the fresh SPCL with the used one,the later one(Fig.4d)showed that its surface covered with pits and pores,the result of biodegradation was visible.3.5.Microbial community of biofilmIn SPCL biofilm sample,most abundant sequences were assigned to the node of bacteria or to its descendants,and a few sequences of eukaryotic organisms were also received. In bacteria,the vast majority of sequences(99.71%)belonged to one of the six major phyla:Proteobacteria,Bacteroidetes,Chloroflexi,Firmicutes,Spirochaetes and Actinobacteria(Fig.5), and Proteobacteria was the most abundant phylum(85.50%) which was mainly-proteobacteria(82.39%).-proteobacteria were reported to be abundant in activated sludge of denitrifying reactors[42].ProteobacteriaBacteroidetesChloroflexiFirm icutesSpirochaetesActinobacteria 0153045607590Relativeabundance(%)Fig.5.Relative abundance of the main phyla identified on SPCL biofilm sample.Only phyla with a relative abundance greater than1%were shown.These six predomi-nant phyla together account for>99.71%of sequences identified.Total numbers of sequences was N=9623.436Z.Shen et al./Journal of Hazardous Materials 250–251 (2013) 431–438Di a p ho ro b a c t e r A c i d o v o r a x De c h l o r o m o n a s A l i c y c l i p h i l u s R o s e if l e x u s P r e v o t e l l ac e a e u n c u l t u r ed T re p o n e m a C l o a c i b a c t e r i u m P e c t i n a t u s S t e n o t r o p h o m o n a s C e l l u l o m o n a s D e s u lf o v i b r i o A z o s p i r a F l a v o b a c t e r i u m A n a e r o a r c u s 05101520253035R e l a t i v e a b u n d a n c e (%)Fig.6.Relative abundance of the main genera identified on SPCL biofilm sample.Only genera with a relative abundance greater than 1%are shown.These 15pre-dominant genera together account for >93.79%of genera identified.At the genus level,sequences from SPCL represented 58differ-ent genera,but 53.42%of the sequences were not related to the known bacteria.Diaphorobacter and Acidovorax constituted 52.75%of the identified genera in SPCL biofilm sample (Fig.6).Diaphorobac-ter was reported to be denitrifying bacteria [43].Both nitrate and nitrite reductase activities were presented in eight strains of Diaphorobacter isolated [44].However,the nitrate reduction rate was 1.5times more than the nitrite reduction in Diaphorobac-ter sp.,but a nitrite accumulation was also received especially at high nitrate concentration,suggesting that Diaphorobacter possi-bly transfers electron sequentially in the denitrification system from nitrate to dinitrogen formation [45].Acidovorax species are commonly observed in wastewater treatment reactors and have been shown to be able to metabolize several different carbon sources [46].For example,Acidovorax avenae subsp.avenae LMG 17238can be successfully used ethanol,methanol,sodium acetate,glucose and poly(-caprolactone)as carbon source for denitrifica-tion [47].Coates et al.[48]reported two Dechloromonas strains,RCB and JJ that can completely mineralize various mono-aromatic compounds including benzene to CO 2in the absence of O 2with nitrate as the electron acceptor.Hong et al.[49]reported that Alicycliphilus was one of the abundant genus in the denitrifying bioreactor.Alicycliphilus denitrificans was the main denitrifier that could use nitrate,nitrite,and oxygen as electron acceptors as reported by Mechichi et al.[50].Stenotrophomonas was isolated directly from a continuous up-flow fixed-bed denitrification reac-tor using poly(3-hydroxybutyrate-co-3-hydroxyvalerate)(PHBV)granules as biofilm carrier,carbon source and electron donor [51].Flavobacterium sp.S6was reported as amylolytic bacterial [52].Most species of Desulfovibrio can oxidize organic compounds such as volatile fatty acids (VFA)and lactate incompletely to acetate [53],and Cellulomonas as cellulose-hydrolysing bacterial was found in sludge from a methanogenic reactor treating paper mill wastewater [54].Rarefaction analysis was used to estimate the richness of total bacterial communities of SPCL biofilm.The steepness curves were received (Fig.7),suggesting that the sampling completeness is low and a large fraction of the species diversity has not yet beensampled.Fig.7.Rarefaction curves of SPCL biofilm sample.The number of OTUs with different cutoff values was plotted as a function of the number of sequences sampled.The 0.03,0.05and 0.1curves contain OTUs with differences that do not exceed 3%,5%and 10%,respectively.3.6.Mass balance of carbon and nitrogen for denitrificationAfter operation for 280d,nitrogen and carbon mass balances were calculated according to the carbon consumption and the removed nitrogen,the results are given in Table 2.For carbon bal-ance,the input and residual mass of SPCL in column,the carbon content of SPCL (42.65%)and the loss of C from effluent DOC were used to calculate the total carbon utilized by microorganism.For nitrogen balance,influent nitrate was total input N,while effluent nitrate,nitrite and ammonium concentration were integrated to calculate total output N,thus the total removed N was obtained by total input N subtracting total output N.The total output N mass was 31.14%of total input,and it mainly due to the over-loading nitrate in phase 2(Fig.2).A large amount loss of C was observed (from effluent DOC),probably mainly due to the high release and accumulation of dissolved organic carbon at start-up period.According to the carbon consumed by microorgan-isms,the removal of 1g nitrogen required 1.01g carbon (utilized by microorganism),which is equivalent of 2.36g SPCL.However,due to the release of organic compounds in effluent,the required mass of SPCL increased to 4.72g/g N.The consumption PCL for removing 1kg nitrate-N was calculated to be 1.33–1.77kg [1].Table 2Mass balance of nitrogen and carbon for denitrification.ItemMass (g)Nitrogen In N from influent nitrate 104.66OutN from effluent nitrate 26.42N from effluent nitrite4.01N from effluent ammonium 2.16TotalTotal N input 104.66Total N output 32.59Total N removed72.08Carbon In C from SPCL202.96OutC measured from DOC 72.46Residual Residual C in column 57.80TotalTotal C utilized72.69。