蛋白质的分选课件
合集下载
《蛋白质的分选》课件

细胞生物学技术
利用细胞生物学技术,如荧光标记、免疫荧光染色等,观察蛋白质在细胞内的定位和动 态变化,揭示蛋白质分选的细胞生物学过程。
蛋白质分选的潜在治疗策略
靶向治疗
针对异常表达的蛋白质或蛋白质分选相关基 因进行靶向治疗,以纠正异常的蛋白质分选 过程,治疗相关疾病。
基因治疗
通过基因工程技术,对相关基因进行修饰或敲除, 以改变蛋白质的表达和分选,达到治疗目的。
膜泡运输
01
指蛋白质在细胞质基质中形成膜泡,然后通过胞吐作用将膜泡
释放到细胞外或细胞内的其他部位。
膜泡运输的类型
02
包括内吞作用、外排作用和胞饮作用等,每种类型都有其特定
的运输途径和作用。
膜泡运输的机制
03
涉及多种蛋白质和细胞器的协同作用,如网格蛋白、细胞骨架
和溶酶体等。
蛋白质的细胞质运输途径
细胞质运输
针对蛋白质分选过程的治疗策略可能 对癌症治疗具有重要意义。
蛋白质分选与神经退行性疾病的关系
神经退行性疾病是指神经元或 神经胶质细胞逐渐退化并导致 功能障碍的一类疾病,如帕金
森病、阿尔茨海默病等。
某些神经退行性疾病可能与蛋 白质聚集物的形成有关,这些 聚集物可能干扰蛋白质的正常 分选和功能。
蛋白质分选异常可能导致神经 元死亡和神经退行性疾病的发
3
临床应用转化
将蛋白质分选的研究成果转化为临床应用,开发 新的治疗策略和方法,为疾病治疗提供更多选择 。
THANKS
THANK YOU FOR YOUR WATCHING
05
蛋白质分选的研究前景与展望
蛋白质分选的研究方法与技术
蛋白质组学技术
利用蛋白质组学技术,如质谱分析、蛋白质免疫印迹等,对蛋白质进行定性和定量分析 ,深入了解蛋白质分选的机制和过程。
利用细胞生物学技术,如荧光标记、免疫荧光染色等,观察蛋白质在细胞内的定位和动 态变化,揭示蛋白质分选的细胞生物学过程。
蛋白质分选的潜在治疗策略
靶向治疗
针对异常表达的蛋白质或蛋白质分选相关基 因进行靶向治疗,以纠正异常的蛋白质分选 过程,治疗相关疾病。
基因治疗
通过基因工程技术,对相关基因进行修饰或敲除, 以改变蛋白质的表达和分选,达到治疗目的。
膜泡运输
01
指蛋白质在细胞质基质中形成膜泡,然后通过胞吐作用将膜泡
释放到细胞外或细胞内的其他部位。
膜泡运输的类型
02
包括内吞作用、外排作用和胞饮作用等,每种类型都有其特定
的运输途径和作用。
膜泡运输的机制
03
涉及多种蛋白质和细胞器的协同作用,如网格蛋白、细胞骨架
和溶酶体等。
蛋白质的细胞质运输途径
细胞质运输
针对蛋白质分选过程的治疗策略可能 对癌症治疗具有重要意义。
蛋白质分选与神经退行性疾病的关系
神经退行性疾病是指神经元或 神经胶质细胞逐渐退化并导致 功能障碍的一类疾病,如帕金
森病、阿尔茨海默病等。
某些神经退行性疾病可能与蛋 白质聚集物的形成有关,这些 聚集物可能干扰蛋白质的正常 分选和功能。
蛋白质分选异常可能导致神经 元死亡和神经退行性疾病的发
3
临床应用转化
将蛋白质分选的研究成果转化为临床应用,开发 新的治疗策略和方法,为疾病治疗提供更多选择 。
THANKS
THANK YOU FOR YOUR WATCHING
05
蛋白质分选的研究前景与展望
蛋白质分选的研究方法与技术
蛋白质组学技术
利用蛋白质组学技术,如质谱分析、蛋白质免疫印迹等,对蛋白质进行定性和定量分析 ,深入了解蛋白质分选的机制和过程。
蛋白质的分选
4.细胞质基质起始和粗内质网合成的蛋白质主要包括:
(1)胞外分泌蛋白 常以分泌泡的形式通过细胞的外排作用输送到细胞外。 (2)膜整合蛋白 细胞质膜、内质网、高尔基体和溶酶体膜上的膜蛋白等都具有 方向性,其方向性在内质网上合成时就已确定。 (3)构成细胞器的可溶性驻留蛋白 有些驻留蛋白需要与其它细胞组分严格隔离,如溶酶体与植物 液泡中的酸性水解酶类;内质网、高尔基体和胞内体 (endosome)中固有的蛋白以及其它有重要生物活性的蛋白。
共翻译转运(Cotranslational translocation) 蛋白质类型:
(1)胞外分泌蛋白:分泌泡形式胞吐到胞外。 (2)膜整合蛋白:细胞质膜、内质网、高尔基体和溶酶 体的膜蛋白等,具有方向性。 (3)内膜系统细胞器内的可溶性驻留蛋白:需要与其它 细胞组分严格隔离,如溶酶体与植物液泡中酸性水解酶等。
1.蛋白质分选的两条基本途径:
共翻译转运(Cotranslational translocation),在细 胞质基质,蛋白质合成起始后,转移至粗面内质网,至高 尔基体,溶酶体,分泌泡,细胞膜乃至胞外。
翻译后转运(post-translational translocation), 在细胞质基质中完成多肽链的合成,然后转运至细胞质基 质特定部位或膜围绕的细胞器。包括线粒体、叶绿体、过 氧化物酶体、内质网及细胞核,细胞质基质驻留蛋白、骨 架蛋白等。
膜泡运输为细胞内膜系统进行物质转运的主要方式, 定向运输取决于膜和膜泡(至少十数种)的表面识别特征。
转运膜泡类型(按功能分类): 1.COPII有被小泡 2.COPI有被小泡 3.网格蛋白有被小泡 内膜系统细胞器保持其特异性蛋白组成的两种机制 有被小泡与靶膜的融合:选择性融合 内膜系统各结构保持形态与膜成分相对稳定的方法
第8章-蛋白质分选和膜泡运输

• Rabs促进和调节运输小泡的停泊和融合。 • Rabs还有许多效应因子,帮助运输小泡聚集和靠
近靶膜,触发SNAREs释放它的效应因子。许多运 输小泡只有在包含了特定的Rabs和SNAREs之后才 能形成。
Rabs in docking
四、受体介导的内吞
• 批量内吞(Bulk-phase endocytosis):非特异性的 摄入细胞外物质,穴样内陷(caveolae)是发生批 量内吞的部位。
三种衣被小泡的功能
衣被类型 GTP酶 组成与衔接蛋白 clathrin ARF Clathrin重链与轻链,AP2
运输方向 质膜→内体
Clathrin重链与轻链,AP1 高尔基体→内体
COP I
ARF
Clathrin重链与轻链,AP3 COPαββ’γδεζ
高尔基体→溶酶体, 植物液泡
高尔基体→内质网
COP II Vesicles
二、衣被形成
• 衣被召集GTP酶:为G蛋白,活化状态可引起衣被 蛋白聚集,包括ARF和SAR 1。存在于细胞质,激活 后转位到膜上。
• ARF:参与clathrin和COP I衣被的形成。 • SAR 1:参与COP II衣被的形成。
• ER上形成COPII小泡时,SAR1交换GDP/GTP而激活。 • 激活的SAR1暴露出脂肪酸链尾巴,插入ER膜,促
输入质体
+H3N-Met-Val-Ala-Met-Ala-Met-Ala-Ser-Leu-Gln-Ser-Ser-Met-Ser-Ser-LeuSer-Leu-Ser-Ser-Asn-Ser-Phe-Leu-Gly-Gln-Pro-Leu-Ser-Pro-Ile-Thr-LeuSer-Pro-Phe-Leu-Gln-Gly-
近靶膜,触发SNAREs释放它的效应因子。许多运 输小泡只有在包含了特定的Rabs和SNAREs之后才 能形成。
Rabs in docking
四、受体介导的内吞
• 批量内吞(Bulk-phase endocytosis):非特异性的 摄入细胞外物质,穴样内陷(caveolae)是发生批 量内吞的部位。
三种衣被小泡的功能
衣被类型 GTP酶 组成与衔接蛋白 clathrin ARF Clathrin重链与轻链,AP2
运输方向 质膜→内体
Clathrin重链与轻链,AP1 高尔基体→内体
COP I
ARF
Clathrin重链与轻链,AP3 COPαββ’γδεζ
高尔基体→溶酶体, 植物液泡
高尔基体→内质网
COP II Vesicles
二、衣被形成
• 衣被召集GTP酶:为G蛋白,活化状态可引起衣被 蛋白聚集,包括ARF和SAR 1。存在于细胞质,激活 后转位到膜上。
• ARF:参与clathrin和COP I衣被的形成。 • SAR 1:参与COP II衣被的形成。
• ER上形成COPII小泡时,SAR1交换GDP/GTP而激活。 • 激活的SAR1暴露出脂肪酸链尾巴,插入ER膜,促
输入质体
+H3N-Met-Val-Ala-Met-Ala-Met-Ala-Ser-Leu-Gln-Ser-Ser-Met-Ser-Ser-LeuSer-Leu-Ser-Ser-Asn-Ser-Phe-Leu-Gly-Gln-Pro-Leu-Ser-Pro-Ile-Thr-LeuSer-Pro-Phe-Leu-Gln-Gly-
蛋白质的分选ppt课件

(肉豆蔻酸,形成脂蛋白)
22
可编辑课件PPT
2.蛋白质分选途径分四种基本类型(据转运方式或机制):
1)蛋白质的跨膜转运(transmembrane transport):细胞质基 质中合成,再转运至线粒体、质体、过氧化物酶体等。
涉及到信号序列(寻靶序列targetting sequence,空间定
Sar1p(GTP结合蛋白,分子开关)。
COPII有被小泡组装、出芽调控
27
可编辑课件PPT
2.COPI有被小泡
功能:介导细胞内逆向运输(retrograde transport),负 责从高尔基体膜泡间、CGN到内质网的物质运输。
回收再循环的膜脂双层、v-SNAREs(vesicle membrane
15
可编辑课件PPT 16
可编辑课件PPT
共翻译转运(cotranslocation)
决定蛋白定位和构象的氨基酸序列: 开始转移序列(start transfer sequence):引导肽链
穿过内质网膜的信号肽。 停止转移序列(stop transfer sequence):有些序列
与内质网膜有很强的亲和力,而结合在脂双层之中,不再 转入内质网腔,形成跨膜蛋白。 各类型跨膜蛋白
peptide,transit peptides/sequences)转移到细胞器中。
转移过程,需ATP使多肽去折叠,需其他蛋白质(如
热休克蛋白Hsp70,分子伴侣)使其能够正确折叠为有功
能的蛋白。
20
可编辑课件PPT
信号因子:
信号序列(signal sequence):存在于蛋白质一级结构上的 线性序列,通常由15-60个氨基酸残基组成,有些信号序列 在完成蛋白质的定向转移后被信号肽酶(signal peptidase) 切除;通常信号序列对所引导的蛋白质没有特异性要求。
22
可编辑课件PPT
2.蛋白质分选途径分四种基本类型(据转运方式或机制):
1)蛋白质的跨膜转运(transmembrane transport):细胞质基 质中合成,再转运至线粒体、质体、过氧化物酶体等。
涉及到信号序列(寻靶序列targetting sequence,空间定
Sar1p(GTP结合蛋白,分子开关)。
COPII有被小泡组装、出芽调控
27
可编辑课件PPT
2.COPI有被小泡
功能:介导细胞内逆向运输(retrograde transport),负 责从高尔基体膜泡间、CGN到内质网的物质运输。
回收再循环的膜脂双层、v-SNAREs(vesicle membrane
15
可编辑课件PPT 16
可编辑课件PPT
共翻译转运(cotranslocation)
决定蛋白定位和构象的氨基酸序列: 开始转移序列(start transfer sequence):引导肽链
穿过内质网膜的信号肽。 停止转移序列(stop transfer sequence):有些序列
与内质网膜有很强的亲和力,而结合在脂双层之中,不再 转入内质网腔,形成跨膜蛋白。 各类型跨膜蛋白
peptide,transit peptides/sequences)转移到细胞器中。
转移过程,需ATP使多肽去折叠,需其他蛋白质(如
热休克蛋白Hsp70,分子伴侣)使其能够正确折叠为有功
能的蛋白。
20
可编辑课件PPT
信号因子:
信号序列(signal sequence):存在于蛋白质一级结构上的 线性序列,通常由15-60个氨基酸残基组成,有些信号序列 在完成蛋白质的定向转移后被信号肽酶(signal peptidase) 切除;通常信号序列对所引导的蛋白质没有特异性要求。
《蛋白分选》PPT课件

高尔基体 → 细胞各部位。
1.膜泡运输
膜泡运输是蛋白运输的一种特有的方式,普遍存在 于真核细胞中。在转运过程中不仅涉及蛋白本身的 修饰、加工和组装,还涉及到多种不同膜泡定向运 输及其复杂的调控过程。
三种不同类型的包被小泡具有不同的物质运输作用
网格蛋白包被小泡 、 COPII包被小泡 、COPI包被小泡
上的线性序列,通常15-60个氨基酸残基,每一种信号序 列决定特殊的蛋白质转运方向。 信号斑(signal patch):存在于完成折叠的蛋白质中, 构成信号斑的信号序列之间可以不相邻,折叠在一起构 成蛋白质分选的信号。
一、蛋白的分选
二、蛋白的分选途径
跨膜转运:基质中合成的蛋白转运至线粒体等细胞器 门控转运:蛋白的核输入和核输出(核质运输) 膜泡运输:转运小泡进行,介导蛋白从内质网 →
后转移:蛋白合成完成后再转移的方式 如线粒体蛋白 其指导转移的序列称导肽
转移序列与蛋白的跨膜结构
转移序列与蛋白的跨膜结构
4.几种典型的蛋白分选信号
信号功能 蛋白进入ER
滞留在ER中 蛋白进入线粒体
进入细胞核 进入过氧化物酶体
信号序列
+H3N-Met-Met-Ser-Phe-Val-Ser-Leu-Ler-Leu-ValGly-Ile-Leu-Phe-Trp-Ala-Thr-Glu-Ala-Glu-GlnLeu-Thr-Lys-Cys-Glu-Val-Phe-GlnLys-Asp-Glu-Leu-COO-
在受体介导的胞吞途径中,将物质从质膜→ 内吞泡(细胞质) → 胞内体 → 溶酶体
高尔基体TGN是网格蛋白包被小泡形成的发源地
网格蛋白有被小泡形成示意图
②、 COPII包被小泡
负责从内质网 → 高尔基体的物质运输; COPII包被蛋白由5种蛋白亚基组成;包被蛋白的
1.膜泡运输
膜泡运输是蛋白运输的一种特有的方式,普遍存在 于真核细胞中。在转运过程中不仅涉及蛋白本身的 修饰、加工和组装,还涉及到多种不同膜泡定向运 输及其复杂的调控过程。
三种不同类型的包被小泡具有不同的物质运输作用
网格蛋白包被小泡 、 COPII包被小泡 、COPI包被小泡
上的线性序列,通常15-60个氨基酸残基,每一种信号序 列决定特殊的蛋白质转运方向。 信号斑(signal patch):存在于完成折叠的蛋白质中, 构成信号斑的信号序列之间可以不相邻,折叠在一起构 成蛋白质分选的信号。
一、蛋白的分选
二、蛋白的分选途径
跨膜转运:基质中合成的蛋白转运至线粒体等细胞器 门控转运:蛋白的核输入和核输出(核质运输) 膜泡运输:转运小泡进行,介导蛋白从内质网 →
后转移:蛋白合成完成后再转移的方式 如线粒体蛋白 其指导转移的序列称导肽
转移序列与蛋白的跨膜结构
转移序列与蛋白的跨膜结构
4.几种典型的蛋白分选信号
信号功能 蛋白进入ER
滞留在ER中 蛋白进入线粒体
进入细胞核 进入过氧化物酶体
信号序列
+H3N-Met-Met-Ser-Phe-Val-Ser-Leu-Ler-Leu-ValGly-Ile-Leu-Phe-Trp-Ala-Thr-Glu-Ala-Glu-GlnLeu-Thr-Lys-Cys-Glu-Val-Phe-GlnLys-Asp-Glu-Leu-COO-
在受体介导的胞吞途径中,将物质从质膜→ 内吞泡(细胞质) → 胞内体 → 溶酶体
高尔基体TGN是网格蛋白包被小泡形成的发源地
网格蛋白有被小泡形成示意图
②、 COPII包被小泡
负责从内质网 → 高尔基体的物质运输; COPII包被蛋白由5种蛋白亚基组成;包被蛋白的
蛋白质分选,膜泡运输2.ppt

膜泡运输的定向机制
各类运输小泡之所以能够被准确地和靶膜融合,是因 为运输小泡表面的标志蛋白能被靶膜上的受体识别, 其中涉及识别过程的两类关键性的蛋白质是SNAREs (soluble NSF attachment protein receptor)和Rabs( targeting GTPase)。其中:
第三节 细胞内蛋白质的分选与膜泡运输
二、蛋白质分选的基本途径和类型
共
翻
翻
译
译
转
后
运
转
的
运
蛋
的
白
非
质
分
分
泌
泌
途
途
径
径
二、蛋白质分选的基本途径和类型
蛋白质的转运类型:
1. 跨膜转运(transmembrane transport):蛋白质通 过跨膜通道进入目的地。如细胞质中合成的蛋白质 通过线粒体上的转位因子(translocator)进入线粒 体。 2. 膜泡运输(vesicular transport):被运输的物质在 内质网或高尔基体中加工成衣被小泡,选择性地运 输到靶细胞器。 3. 选择性的门控转运(gated transport):如通过核 孔复合体的运输(输入或输出)。 4. 细胞质基质中的蛋白质转运:与细胞骨架系统密 切相关。
• 许多膜标志蛋白存在于不止一种细胞器,不同的膜 标志蛋白组合,决定膜的表面识别特征。
三类有被小泡及其功能
电镜下的衣被小泡
三类有被小泡及其功能
1. COP II 有被小泡 介导从内质网到高尔基体的物质运输。 COP II衣被由5种蛋白质亚基构成: Sec13, Sec31,
Sec23, Sec24和Sar1(GTP酶)。
蛋白质分选651.pptx
第三十一Fra bibliotek,共四十二页。
第十页,共四十二页。
发现信号(xìnhào)肽序列之后,人们有相继发现一系列蛋 白质分选信号(xìnhào)序列,统称信号(xìnhào)序列 〔signal sequence〕。有些信号序列还可形成三维结构 的信号斑〔signal-patch〕。
第十一页,共四十二页。
信号识别(shíbié)颗粒
合成位置:真核细胞中除线粒体和植物细胞叶 绿体中能合成少量蛋白质外,绝大多数蛋白质 都是由核基因编码,在游离核糖体上、或糙面 内质网膜结合核糖体上合成。
作用部位:蛋白质发挥结构或功能的部位几乎遍布
(biàn bù)细胞的各种区间或组份。
第三页,共四十二页。
因此,必然存在不同的机制以确保蛋白质的分选,转运 至细胞的特定部位。蛋白质也只有各就各位并组装成结 构与功能的复合体,才能参与实现细胞的各种生命 (shēngmìng)活动。
这种转运方式在蛋白质跨膜的过程中不仅需要消耗ATP使 多肽去折叠,而且还需要一些分子伴侣蛋白的帮助〔如热 休克蛋白Hsp70〕,以帮助蛋白正确折叠形成有功能的蛋 白质。
第十八页,共四十二页。
二、蛋白质分选转运的根本途径(tújìng)与类型
核基因编码的蛋白质的分选大体可分为(fēn wéi)2条途径: 〔一〕共翻译转运途径(tújìng):蛋白在信号肽的引导下,边翻译边
第七页,共四十二页。
Günter Blobel
Blobel 在1971 年首次提出内质网膜分泌蛋白的氨基端序列带有转运信 息。这一设想在1975 年扩展为信号假说,1980 年扩展为在细胞内蛋白 转运及膜生物(shēngwù)合成过程中的普遍学说。发现并分析了某些分 泌蛋白、整合膜蛋白、叶绿体、线粒体基质蛋白和溶酶体蛋白的信号序 列; 从内质网别离得到了信号识别颗粒SRP; 在线粒体和叶绿体膜上相 继别离得到了识别SRP 的受体; 纯化得到了内质网相关的信号肽酶; 用 电生理的方法证实了内质网上蛋白运输通道的存在。
第十页,共四十二页。
发现信号(xìnhào)肽序列之后,人们有相继发现一系列蛋 白质分选信号(xìnhào)序列,统称信号(xìnhào)序列 〔signal sequence〕。有些信号序列还可形成三维结构 的信号斑〔signal-patch〕。
第十一页,共四十二页。
信号识别(shíbié)颗粒
合成位置:真核细胞中除线粒体和植物细胞叶 绿体中能合成少量蛋白质外,绝大多数蛋白质 都是由核基因编码,在游离核糖体上、或糙面 内质网膜结合核糖体上合成。
作用部位:蛋白质发挥结构或功能的部位几乎遍布
(biàn bù)细胞的各种区间或组份。
第三页,共四十二页。
因此,必然存在不同的机制以确保蛋白质的分选,转运 至细胞的特定部位。蛋白质也只有各就各位并组装成结 构与功能的复合体,才能参与实现细胞的各种生命 (shēngmìng)活动。
这种转运方式在蛋白质跨膜的过程中不仅需要消耗ATP使 多肽去折叠,而且还需要一些分子伴侣蛋白的帮助〔如热 休克蛋白Hsp70〕,以帮助蛋白正确折叠形成有功能的蛋 白质。
第十八页,共四十二页。
二、蛋白质分选转运的根本途径(tújìng)与类型
核基因编码的蛋白质的分选大体可分为(fēn wéi)2条途径: 〔一〕共翻译转运途径(tújìng):蛋白在信号肽的引导下,边翻译边
第七页,共四十二页。
Günter Blobel
Blobel 在1971 年首次提出内质网膜分泌蛋白的氨基端序列带有转运信 息。这一设想在1975 年扩展为信号假说,1980 年扩展为在细胞内蛋白 转运及膜生物(shēngwù)合成过程中的普遍学说。发现并分析了某些分 泌蛋白、整合膜蛋白、叶绿体、线粒体基质蛋白和溶酶体蛋白的信号序 列; 从内质网别离得到了信号识别颗粒SRP; 在线粒体和叶绿体膜上相 继别离得到了识别SRP 的受体; 纯化得到了内质网相关的信号肽酶; 用 电生理的方法证实了内质网上蛋白运输通道的存在。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 主要成分:由3-4个Sec61蛋白复合体构成(哺乳动物),每 个Sec61蛋白由三条肽链组成。
学习交流PPT
13
二.蛋白质分选的基本途径与类型
• 1.蛋白质分选的基本途径:
•
根据蛋白转运与翻译的时空关系分类。
•
内质网内蛋白质合成过程:共翻译转运
•
细胞质基质合成蛋白质的转运方式:翻译后转运
• 2.蛋白质的转运类型:
• 决定蛋白定位和构象的氨基酸序列:
• 开始转移序列(start transfer sequence):引导肽链 穿过内质网膜的信号肽。
• 停止转移序列(stop transfer sequence):有些序列 与内质网膜有很强的亲和力,而结合在脂双层之中,不再 转入内质网腔,形成跨膜蛋白。
• 含3个主要功能部位:信号肽识别结合位点(P54,多Met,
具GTP酶活)、翻译暂停位点(P9/P14)、SRP受体蛋白结合位
点(P68/P72)。
学习交流PPT
7
P9/P14
翻译暂停位点
信号肽序列结合位点
P54
含蛋氨酸侧链 结合信号肽疏水核心
SRP受体(停泊蛋白,DP)结合位点
学习交流PPT
8
2
细胞内蛋白质的分选与膜泡运输
• 一.信号假说与蛋白质分选信号 • 二.蛋白质分选基本途径与类型 • 三.膜泡运输
学习交流PPT
3
一.信号假说与蛋白质分选信号
• 1.信号肽的发现与信号假说的提出 • 2.信号肽的序列特点 • 3.信号识别相关因子:信号识别颗粒、停泊蛋白和移位子 • 4.信号假说的实验验证及rER蛋白质合成过程确定
4
+
++ +
信号肽切除,多肽链进入
微粒体中
“+”和“-”分别代表反应混合物中存在(+)或不存在(-)该物质。
学习交流PPT
11
信号序列
内质网内的可溶性蛋白质合成
信号识别 颗粒
信号 识别 颗粒 受体
易位子
信号肽酶
学习交流PPT
可溶性蛋白质
12
• 内质网膜上的移位子(translocon)为蛋白复合体, 直径约8.5nm,中心有一个直径为2nm的“通道”,其功能 与新合成的多肽进入内质网有关。
• 分泌蛋白N端序列作为信号肽(signal peptide),指导 分泌性蛋白到内质网膜上合成;在信号肽引导下,蛋白质边 合成边通过移位子蛋白复合体进入内质网腔;在蛋白合成结 束之前信号肽被切除。
• G. Blobel因此获得1999年诺贝尔医学和生理学奖。
学习交流PPT
5
• 信号肽(signal peptide):又称开始转移序列(start transfer sequence),位于新合成肽链N端,引导新合成的肽链转移到 内质网上合成的信号序列。
• SRP受体(SPR receptor):又称停泊蛋白(docking protein,DP),72KD,异二聚体蛋白,属内质网膜整合蛋白, 与SRP特异结合。
• 一个亚基(640个Aa)暴露于细胞质,亲水;
• 另一亚基(300个Aa)嵌入膜内,疏水。
学习交流PPT
9
• 4.信号假说的实验验证及rER蛋白质合成过程确定
• 信号肽一级序列:16-26Aa,包括疏水核心(h)、C端 (c)和N端(n)三个区域。
• 特点:无严格的专一性,可将引导不同蛋白序列。
学习交流PPT
6
• 信号识别颗粒(ຫໍສະໝຸດ RP):6种不同多肽和1个7S RNA(约300bp) 组成的复合物。
•
• • 含2个结构域:信号肽识别结构域,核糖体结合结构域
• 广义的过程包括:蛋白质从合成直到降解或分泌胞外。
细胞内合成的蛋白质、脂类等物质之所以能够定向的 转运到特定的细胞器,取决于两个方面。
信号序列
(signal sequeence)
蛋白质中包含特殊的信号序列。
识别(装so置rtin(g r分ece选pto受r)体)细胞器上具特定的信号识别装置。
学习交流PPT
•
• 翻译后转运(post-translational translocation), 在细胞质基质中完成多肽链的合成,然后转运至细胞质基 质特定部位或膜围绕的细胞器。包括线粒体、叶绿体、过 氧化物酶体、内质网及细胞核,细胞质基质驻留蛋白、骨 架蛋白等。
学习交流PPT
15
学习交流PPT
16
• 共翻译转运(cotranslocation)
学习交流PPT
4
• 信号肽(signal peptide)的发现:
• 1972年Milstein等用骨髓瘤细胞提取的免疫球蛋白较分 泌到胞外的成熟免疫球蛋白在N端多出一段肽链,推测起信 号作用。
• 信号假说(Signal hypothesis):分泌蛋白合成与定位
• 1975年美国G. Blobel和D. Sabatini正式提出。
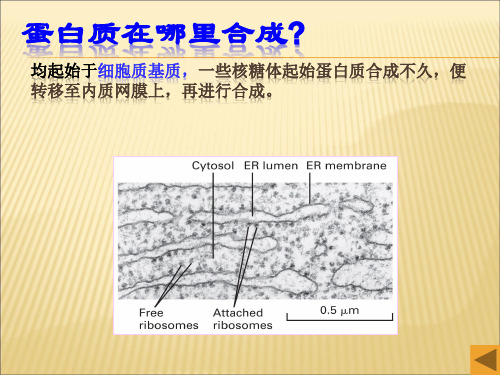
蛋白质在哪里合成?
均起始于细胞质基质,一些核糖体起始蛋白质合成不久,便 转移至内质网膜上,再进行合成。
学习交流PPT
1
细胞内蛋白质的分选与膜泡运输
• 蛋白质的定向转运(protein targeting)或分选 (protein sorting):结构与功能各异的蛋白质,在细胞 质基质或糙面内质网的核糖体上合成,然后转运到正确的 部位,并组装成结构与功能的复合体,参于生命活动的过 程。
• 根据蛋白的转运方式或机制分四类
• 3.细胞内合成蛋白质定向转运主要取决于两个方面:
•
其一蛋白质中包含特殊的信号序列
•
其二细胞器上特定的信号识别装置与分选受体
学习交流PPT
14
• 1.蛋白质分选的两条基本途径:
• 共翻译转运(Cotranslational translocation),在细 胞质基质,蛋白质合成起始后,转移至粗面内质网,至高 尔基体,溶酶体,分泌泡,细胞膜乃至胞外。
学习交流PPT
10
非细胞系统中(cell-free system)蛋白质的翻译 过程与SRP、DP和微粒体的关系
实验 含有编码信号 SRP DP 微粒体
结
果
组别 序列的mRNA
1
+
-- -
产生含信号肽的完整多肽
2
+
+- -
合成70~100氨基酸残基后,
肽链停止延伸
3
+
++ -
产生含信号肽的完整多肽
• • Blobel、Dobberstein和Walter用分离微粒体和非细胞体
系(cell-free system)实验证实:信号序列的存在,和分 泌蛋白合成过程。
• 实验方法:放射性氨基酸脉冲标记孵育、密度梯度离心获 得微粒体、去垢剂处理对照、蛋白酶降解等。
• RER合成蛋白与信号肽、SRP与DP间的关系实验
学习交流PPT
13
二.蛋白质分选的基本途径与类型
• 1.蛋白质分选的基本途径:
•
根据蛋白转运与翻译的时空关系分类。
•
内质网内蛋白质合成过程:共翻译转运
•
细胞质基质合成蛋白质的转运方式:翻译后转运
• 2.蛋白质的转运类型:
• 决定蛋白定位和构象的氨基酸序列:
• 开始转移序列(start transfer sequence):引导肽链 穿过内质网膜的信号肽。
• 停止转移序列(stop transfer sequence):有些序列 与内质网膜有很强的亲和力,而结合在脂双层之中,不再 转入内质网腔,形成跨膜蛋白。
• 含3个主要功能部位:信号肽识别结合位点(P54,多Met,
具GTP酶活)、翻译暂停位点(P9/P14)、SRP受体蛋白结合位
点(P68/P72)。
学习交流PPT
7
P9/P14
翻译暂停位点
信号肽序列结合位点
P54
含蛋氨酸侧链 结合信号肽疏水核心
SRP受体(停泊蛋白,DP)结合位点
学习交流PPT
8
2
细胞内蛋白质的分选与膜泡运输
• 一.信号假说与蛋白质分选信号 • 二.蛋白质分选基本途径与类型 • 三.膜泡运输
学习交流PPT
3
一.信号假说与蛋白质分选信号
• 1.信号肽的发现与信号假说的提出 • 2.信号肽的序列特点 • 3.信号识别相关因子:信号识别颗粒、停泊蛋白和移位子 • 4.信号假说的实验验证及rER蛋白质合成过程确定
4
+
++ +
信号肽切除,多肽链进入
微粒体中
“+”和“-”分别代表反应混合物中存在(+)或不存在(-)该物质。
学习交流PPT
11
信号序列
内质网内的可溶性蛋白质合成
信号识别 颗粒
信号 识别 颗粒 受体
易位子
信号肽酶
学习交流PPT
可溶性蛋白质
12
• 内质网膜上的移位子(translocon)为蛋白复合体, 直径约8.5nm,中心有一个直径为2nm的“通道”,其功能 与新合成的多肽进入内质网有关。
• 分泌蛋白N端序列作为信号肽(signal peptide),指导 分泌性蛋白到内质网膜上合成;在信号肽引导下,蛋白质边 合成边通过移位子蛋白复合体进入内质网腔;在蛋白合成结 束之前信号肽被切除。
• G. Blobel因此获得1999年诺贝尔医学和生理学奖。
学习交流PPT
5
• 信号肽(signal peptide):又称开始转移序列(start transfer sequence),位于新合成肽链N端,引导新合成的肽链转移到 内质网上合成的信号序列。
• SRP受体(SPR receptor):又称停泊蛋白(docking protein,DP),72KD,异二聚体蛋白,属内质网膜整合蛋白, 与SRP特异结合。
• 一个亚基(640个Aa)暴露于细胞质,亲水;
• 另一亚基(300个Aa)嵌入膜内,疏水。
学习交流PPT
9
• 4.信号假说的实验验证及rER蛋白质合成过程确定
• 信号肽一级序列:16-26Aa,包括疏水核心(h)、C端 (c)和N端(n)三个区域。
• 特点:无严格的专一性,可将引导不同蛋白序列。
学习交流PPT
6
• 信号识别颗粒(ຫໍສະໝຸດ RP):6种不同多肽和1个7S RNA(约300bp) 组成的复合物。
•
• • 含2个结构域:信号肽识别结构域,核糖体结合结构域
• 广义的过程包括:蛋白质从合成直到降解或分泌胞外。
细胞内合成的蛋白质、脂类等物质之所以能够定向的 转运到特定的细胞器,取决于两个方面。
信号序列
(signal sequeence)
蛋白质中包含特殊的信号序列。
识别(装so置rtin(g r分ece选pto受r)体)细胞器上具特定的信号识别装置。
学习交流PPT
•
• 翻译后转运(post-translational translocation), 在细胞质基质中完成多肽链的合成,然后转运至细胞质基 质特定部位或膜围绕的细胞器。包括线粒体、叶绿体、过 氧化物酶体、内质网及细胞核,细胞质基质驻留蛋白、骨 架蛋白等。
学习交流PPT
15
学习交流PPT
16
• 共翻译转运(cotranslocation)
学习交流PPT
4
• 信号肽(signal peptide)的发现:
• 1972年Milstein等用骨髓瘤细胞提取的免疫球蛋白较分 泌到胞外的成熟免疫球蛋白在N端多出一段肽链,推测起信 号作用。
• 信号假说(Signal hypothesis):分泌蛋白合成与定位
• 1975年美国G. Blobel和D. Sabatini正式提出。
蛋白质在哪里合成?
均起始于细胞质基质,一些核糖体起始蛋白质合成不久,便 转移至内质网膜上,再进行合成。
学习交流PPT
1
细胞内蛋白质的分选与膜泡运输
• 蛋白质的定向转运(protein targeting)或分选 (protein sorting):结构与功能各异的蛋白质,在细胞 质基质或糙面内质网的核糖体上合成,然后转运到正确的 部位,并组装成结构与功能的复合体,参于生命活动的过 程。
• 根据蛋白的转运方式或机制分四类
• 3.细胞内合成蛋白质定向转运主要取决于两个方面:
•
其一蛋白质中包含特殊的信号序列
•
其二细胞器上特定的信号识别装置与分选受体
学习交流PPT
14
• 1.蛋白质分选的两条基本途径:
• 共翻译转运(Cotranslational translocation),在细 胞质基质,蛋白质合成起始后,转移至粗面内质网,至高 尔基体,溶酶体,分泌泡,细胞膜乃至胞外。
学习交流PPT
10
非细胞系统中(cell-free system)蛋白质的翻译 过程与SRP、DP和微粒体的关系
实验 含有编码信号 SRP DP 微粒体
结
果
组别 序列的mRNA
1
+
-- -
产生含信号肽的完整多肽
2
+
+- -
合成70~100氨基酸残基后,
肽链停止延伸
3
+
++ -
产生含信号肽的完整多肽
• • Blobel、Dobberstein和Walter用分离微粒体和非细胞体
系(cell-free system)实验证实:信号序列的存在,和分 泌蛋白合成过程。
• 实验方法:放射性氨基酸脉冲标记孵育、密度梯度离心获 得微粒体、去垢剂处理对照、蛋白酶降解等。
• RER合成蛋白与信号肽、SRP与DP间的关系实验