吉林蛟河天然红松阔叶林的空间结构分析
吉林蛟河针阔混交林树木生长的空间关联格局

吉林蛟河针阔混交林树木生长的空间关联格局郝珉辉;张忠辉;赵珊珊;张春雨;赵秀海【摘要】以吉林蛟河21.12hm2(660m×320m)针阔混交林样地为对象,利用2009年和2014年森林生长观测数据,研究树木生长的空间自相关格局及其生境影响机制.在样地生境型划分结果的基础上,采用Ripley's L(r)函数分析不同生境型中树木种群空间分布特征;利用标记相关函数分析不同生境型中树木生长特征的空间关联格局.研究结果表明:(1)红松(生境型3∶1-5m)、蒙古栎(生境型3:1-3m)、胡桃楸(生境型2:1-2m;生境型3:1-7m)、黄檗(生境型2∶1-3m;生境型4:1-5m)、水曲柳(生境型3∶1-2m;生境型4:1-2m)、瘤枝卫矛(生境型2∶1-15m)在特定生境和空间尺度上呈随机分布,但空间格局仍以聚集性分布为主;其余10个物种则在全部0-30m尺度上呈聚集分布.(2)标记相关函数分析显示春榆、毛榛、色木槭、瘤枝卫矛和千金榆的径向生长至少在一个生境中表现出正相关格局;暴马丁香、胡桃楸、裂叶榆、瘤枝卫矛、水曲柳、紫椴、糠椴、毛榛、色木槭和白牛槭的径向生长至少在一个生境中表现出负相关格局;红松、黄檗、蒙古栎和簇毛槭的径向生长在全部尺度上均未检测到显著的空间关联格局.因此,不同树种径向生长的空间自相关特征不同,树种生长特征的空间关联格局具有明显的生境依赖性.%This study aimed to determine the spatial autocorrelation patterns of tree growth and the mechanisms influencing habitat heterogeneity.A dataset of observed forest growth was used,with data between 2009 and 2014 from a 21.12 hm2 stem-mapped mixed forest plot (temperate coniferous and broadleaved forest) in Jiaohe,Jilin Province.Based on habitat type classifications,Ripley's L(r) function was adopted to evaluate the spatial distribution characteristics of tree locations.A mark correlation function(MCF) was used to evaluate the spatial autocorrelation characteristics of tree gro wth.Our results showed that (1) Pinus koraiensis (habitat type 3 ∶ 1-5 m),Quercus mongolica (habitat type 3 ∶ 1-3 m),Juglans mandshurica (habitat type 2 ∶ 1-2 m;habitat type 3 ∶ 1-7 m),Phellodendron amurense (habitat type 2 ∶ 1-3 m;habitat type 4∶1-5 m),Fraxinus mandshurica (habitat type 3∶1-2 m;habitat type 4∶1-2 m),and Euonymus verrucosus (habitat type 2∶1-15 m) were randomly distributed at particular scales and for certain habitat types.Nonetheless,they mainly showed clumped distribution patterns.For ten other species,the spatial distribution of tree positions showed an aggregated pattern at a scale of 0-30 m.(2) The mark correlation function analysis indicated that the radial growth of Ulmus davidiana var.japonica,Corylus mandshurica,Acer mono,E.verrucosus,and Carpinus cordata were positively autocorrelated for at least one of the habitat types.In contrast,the radial growth of Syringa reticulatavar.amurensis,J.mandshurica,Ulmuslaciniata,E.verrucosus,F.mandshurica,Tilia amurensis,Tilia mandshurica,C.mandshurica,A.mono,and Acer mandshuricum were negatively autocorrelated at least in one of habitat types.Other species did not show any significant autocorrelations at any of the studied scales.Our results indicate that different species had different spatial autocorrelation characteristics of radial growth,as well as significantly different habitat dependencies.【期刊名称】《生态学报》【年(卷),期】2017(037)006【总页数】9页(P1922-1930)【关键词】径向生长;生境差异;竞争作用;生境依赖性;Ripley's L(r)函数【作者】郝珉辉;张忠辉;赵珊珊;张春雨;赵秀海【作者单位】北京林业大学森林资源与生态系统过程北京市重点实验室,北京100083;吉林省林业科学研究院,长春130033;吉林省林业科学研究院,长春130033;北京林业大学森林资源与生态系统过程北京市重点实验室,北京 100083;北京林业大学森林资源与生态系统过程北京市重点实验室,北京 100083【正文语种】中文森林中普遍存在着种内和种间相互作用,林木个体之间的相互作用可大体分为正向作用和负向作用[1- 2]。
阔叶林发展红松果材林的探索与思考

阔叶林发展红松果材林的探索与思考随着社会经济的不断发展和人们对环境保护的重视,林业资源的合理利用和保护也成为了一个备受关注的话题。
阔叶林是我国森林资源的重要组成部分,而红松是一种生长迅速、木质材料优质的松树品种。
本文将对阔叶林发展红松果材林的探索与思考进行讨论,探索如何更好地利用阔叶林资源,促进林业的可持续发展。
一、红松果材林的特点与优势红松是一种生长快、枝绿叶厚、木材紧密、质地硬、不易龟裂、纹理美观、图案独特,在木材市场上有很高的销售价值。
红松的果实也是一种高蛋白、高脂肪、高营养价值的野生食品,具有广阔的市场前景。
将红松与阔叶林相结合,发展红松果材林,有利于提高林业资源的综合利用率,促进经济效益。
二、阔叶林发展红松果材林的可能性1. 土壤条件与环境适应性红松对土壤的要求并不高,对地下水位的要求较低,适应性广。
我国东北地区的阔叶林分布广泛,土壤肥沃,水土资源丰富,气候适宜,与红松生长要求相符,因此在这些地区发展红松果材林具有较大的可行性。
2. 生长速度和木材质量红松是一种生长速度快的树种,一般3-5年就可以达到很好的经济效益。
而红松的木材质量也相当高,耐冷性强,适应性强,深得市场青睐。
在阔叶林地区种植红松,可以利用其生长速度快的特点,加速木材资源的更新与利用。
3. 综合利用价值红松的果实可以作为野生食品,具有很高的经济价值,可以作为地方特色农产品进行开发和销售。
而红松的木材也非常适合制作家具、建材等产品,具有很高的市场价格。
阔叶林发展红松果材林,不仅可以提高林地资源的综合利用率,还可以促进当地的经济发展。
三、发展红松果材林存在的问题1. 种植技术和经验不足由于红松果材林的种植经验相对较少,尤其是在阔叶林地区的种植,因此需要对种植技术和经验进行深入研究和总结。
如何选择合适的育苗和栽种地点、如何科学施肥、如何控制病虫害等问题都需要找到合适的解决方法,这是发展红松果材林面临的一个重要问题。
2. 市场销售和品牌建设虽然红松的木材具有很高的市场价格,但是目前红松木材市场却并不够活跃,产品积压严重。
吉林省蛟河林业实验区管理局天然林胸径及生长量研究
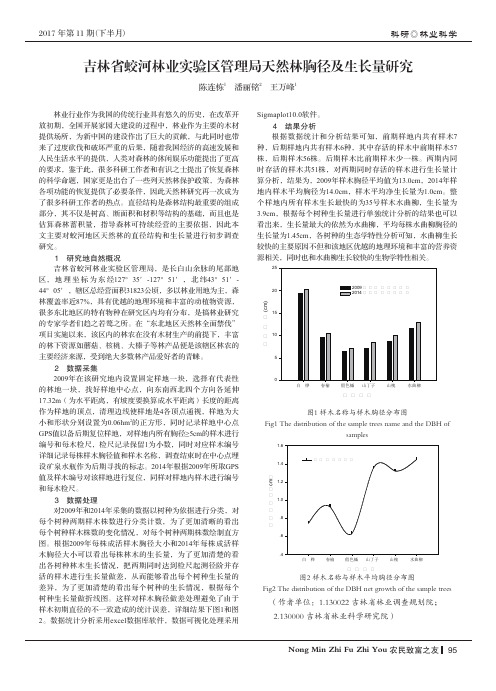
2017 年第 11 期(下半月)Nong Min Zhi Fu Zhi You 农民致富之友95科研◎林业科学吉林省蛟河林业实验区管理局天然林胸径及生长量研究陈连栋1 潘丽铭2 王万峰1林业行业作为我国的传统行业具有悠久的历史,在改革开放初期,全国开展家园大建设的过程中,林业作为主要的木材提供场所,为新中国的建设作出了巨大的贡献,与此同时也带来了过度砍伐和破坏严重的后果,随着我国经济的高速发展和人民生活水平的提供,人类对森林的休闲娱乐功能提出了更高的要求,鉴于此,很多科研工作者和有识之士提出了恢复森林的科学命题,国家更是出台了一些列天然林保护政策,为森林各项功能的恢复提供了必要条件,因此天然林研究再一次成为了很多科研工作者的热点。
直径结构是森林结构最重要的组成部分,其不仅是树高、断面积和材积等结构的基础,而且也是估算森林蓄积量,指导森林可持续经营的主要依据,因此本文主要对蛟河地区天然林的直径结构和生长量进行初步调查研究。
1 研究地自然概况吉林省蛟河林业实验区管理局,是长白山余脉的尾部地区,地理坐标为东经127°35′-127°51′,北纬43°51′-44°05′,辖区总经营面积31823公顷,多以林业用地为主,森林覆盖率近87%,具有优越的地理环境和丰富的动植物资源,很多东北地区的特有物种在研究区内均有分布,是搞林业研究的专家学者们趋之若鹜之所。
在“东北地区天然林全面禁伐”项目实施以来,该区内的林农在没有木材生产的前提下,丰富的林下资源如蘑菇、核桃、大榛子等林产品便是该辖区林农的主要经济来源,受到绝大多数林产品爱好者的青睐。
2 数据采集2009年在该研究地内设置固定样地一块,选择有代表性的林地一块,找好样地中心点,向东南西北四个方向各延伸17.32m (为水平距离,有坡度要换算成水平距离)长度的距离作为样地的顶点,清理边线使样地是4各顶点通视,样地为大小和形状分别设置为0.06hm 2的正方形,同时记录样地中心点GPS 值以备后期复位样地,对样地内所有胸径≥5cm 的样木进行编号和每木检尺,检尺记录保留1为小数,同时对应样木编号详细记录每株样木胸径值和样木名称,调查结束时在中心点埋设矿泉水瓶作为后期寻找的标志。
采伐对吉林蛟河阔叶红松林生态功能的短期影响

采伐对吉林蛟河阔叶红松林生态功能的短期影响吉林省蛟河市的阔叶红松林是一片非常珍贵的自然资源,拥有着丰富的生态功能和重要的生态价值。
然而,此地面临的危机是,由于人类活动而引发的采伐导致了生态系统的扰动和破坏。
本文将从短期影响的角度,对采伐对吉林蛟河阔叶红松林生态功能产生的影响进行分析。
首先,采伐会直接影响到这片阔叶红松林的物种多样性。
由于采伐会大量砍伐树木,导致林木被破坏,生态系统的内部结构发生变化,进而影响森林内部的生物多样性。
在短期内,采伐会直接导致森林生态系统的生物多样性的减少。
其次,采伐会对吉林蛟河阔叶红松林的土壤和水的质量产生直接和间接的影响。
首先,由于采伐破坏了森林的物理结构,导致土壤极度缺水,以及性质的变化。
在短期内,土壤中的有机物质等元素会被大量削减,降低了土壤的生产力和清洁度。
其次,由于采伐破坏了阔叶红松林的树木与树冠的结构,导致雨水的水循环过程发生变化。
长期下来,这些生态功能受到破坏,使得森林无法正常自行保护自然生态系统,最终会直接影响到整个生态系统的健康和稳定。
最后,采伐也会对吉林蛟河阔叶红松林的风景和游憩功能产生影响。
阔叶红松林作为一片赏心悦目、让人神清气爽的美丽风景线,对人们的身心健康有着很大的益处。
然而,采伐破坏了阔叶红松林的美丽景色和游憩环境,影响了人们的心情和健康。
综上所述,采伐对吉林蛟河阔叶红松林的生态功能将产生一系列的短期影响,对森林的物种多样性、土壤和水的质量和森林景观和游憩功能都产生了直接和间接的影响。
因此,保护和维护阔叶红松林的生态环境是非常重要的,需要更加注重环保的策略和措施,避免这一珍贵的自然资源被进一步破坏和损坏。
吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应

第41卷第7期2021年4月生态学报ACTAECOLOGICASINICAVol.41,No.7Apr.,2021基金项目:国家自然科学基金项目(31971650,31600480)收稿日期:2020⁃06⁃05;㊀㊀网络出版日期:2021⁃01⁃28∗通讯作者Correspondingauthor.E⁃mail:zcy_0520@163.comDOI:10.5846/stxb202006051462夏梦洁,王晓霞,郝珉辉,赵秀海,张春雨.吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应.生态学报,2021,41(7):2794⁃2802.XiaMJ,WangXX,HaoMH,ZhaoXH,ZhangCY.Distributionpatternoffunctionaltraitsanditsresponsetotopographicfactorsinaconiferandbroad⁃leavedmixedforestinJiaohe,Jilinprovince.ActaEcologicaSinica,2021,41(7):2794⁃2802.吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应夏梦洁1,王晓霞2,郝珉辉1,赵秀海1,张春雨1,∗1北京林业大学国家林业和草原局森林经营工程技术研究中心,北京㊀1000832吉林省林业勘察设计研究院,长春㊀130022摘要:森林群落的构建过程及其内在机制是生态学研究的热点问题㊂植物功能性状是指能够代表植物的生活史策略,反映植物对环境变化响应的一系列植物属性㊂通过植物功能性状的分布格局及其对环境因素的响应有助于推测群落的构建过程及其内在作用机制㊂以吉林蛟河21.12hm2温带针阔混交林样地为研究对象,采集并测量了样地内34种木本植物的6种不同的功能性状㊂以20mˑ20m的样方为研究单元,通过计算平均成对性状距离指数(meanpairwisetraitdistance;PW)和平均最近邻体性状距离指数(meannearestneighbortraitdistance;NN)来探讨群落中单个性状和综合性状的分布格局㊂同时结合地形因子采用回归分析探讨功能性状的分布格局对局域生境变化的响应㊂基于PW的结果显示:单个性状中除叶面积外,其余性状的分布格局均为聚集分布多于离散分布;基于NN的结果显示:除叶面积和最大树高外,其余性状的分布格局为聚集分布多于离散分布㊂此外,由6种单个性状组成的综合性状的分布格局同样为聚集分布多于离散分布㊂基于回归分析的结果显示:森林群落中功能性状的分布格局受到海拔㊁坡度和坡向等因素的显著影响,而凹凸度的影响则不显著㊂研究结果表明包括环境过滤和生物相互作用的非随机过程能够影响温带针阔混交林的群落构建过程,中性过程对该区域群落构建过程的影响不显著㊂关键词:功能性状;功能性状结构;群落构建机制;环境因素Distributionpatternoffunctionaltraitsanditsresponsetotopographicfactorsinaconiferandbroad⁃leavedmixedforestinJiaohe,JilinprovinceXIAMengjie1,WANGXiaoxia2,HAOMinhui1,ZHAOXiuhai1,ZHANGChunyu1,∗1ResearchCenterofForestManagementEngineeringofStateForestryandGrasslandAdministration,BeijingForestryUniversity,Beijing100083,China2ForestrySurveyandDesignInstituteofJilinProvince,Changchun130022,ChinaAbstract:Understandingthemechanismandprocessofcommunityassemblyisakeyissueinecologicalresearch.Functionaltraits,whichrepresentplantphysiological,morphological,andphenologicalattributes,havedecisiveeffectsonplantgrowth,survival,reproductionanddispersal.Examiningthespatialdistributionpatternofplantfunctionaltraitsanditsresponsetolocalenvironmentalvariationishelpfultospeculatetheunderlyingprocessesandmechanismsthatregulatecommunityassembly.Inthisstudy,weexploredtheunderlyingmechanismsofcommunityassemblyusingthedatasetfroma21.12hm2temperateconiferandbroad⁃leavedmixedforestplotinJiaohe,NortheastChina.Fourtopographicattributes,includingelevation,convexity,slope,andaspect,weremeasuredaslocalenvironmentalfactors.Sixfunctionaltraits,namelyleafarea,specificleafarea,leafcarboncontent,leafnitrogencontent,leafcarbontonitrogenratio,andmaximumtreeheight,werecollectedandmeasuredforall34woodyspeciesthatpresentinthistemperateforestplot.Twofunctionalstructureindices,includingthemeanpair-wisetraitdistance(PW)andthemeannearestneighbortraitdistance(NN),wereusedtoexplorethespatialdistributionpatternofplanttraits.Furthermore,regressionanalyseswereusedtoinvestigatetheresponseoftraitpatternstoenvironmentalvariation.BasedontheresultsofPW,wefoundthat,exceptfortheleafareatraits,theremainingtraitsmainlyshowedtheclumpedspatialpatterns.TheresultsbasedonNNshowedthat,exceptfortheleafareaandmaximumtreeheight,theremainingtraitspresentedtheclumpedspatialpatterns.Theresultsofregressionanalysesshowedthatthedistributionpatternsoffunctionaltraitsweresignificantlyaffectedbyelevation,slope,andaspect.Incontrast,convexityhadnosignificanteffectontraitdistributioninthistemperateforestcommunity.Ourresultsindicatethatthenon⁃randomprocessesincludingenvironmentalfilteringandbiologicalinteractionsplayedimportantrolesinregulatingforestcommunityassembly,whiletheroleofneutralprocesswasnotsignificantinthistemperateconiferandbroad⁃leavedmixedforest.KeyWords:functionaltraits;functionaltraitstructures;communityassembly;environmentalvariables群落构建机制与构建过程是生态学研究的热点问题,生态位理论和中性理论是解释群落构建过程的两个主要理论㊂生态位理论认为生态位完全相同的物种不能共存于群落中,环境过滤和生物间相互作用是影响生物多样性格局的两个重要过程[1⁃2]㊂而中性理论则认为群落中的物种多样性是由长时间物种的随机迁入迁出和生态漂移形成的[3⁃5]㊂扩散限制是中性理论的重要推论,即随着空间距离的增加,群落的相似性会降低,因此扩散限制也是影响群落构建的重要过程㊂植物功能性状是指能够影响植物本身的生理㊁生态学过程,反映植物对环境变化响应的一系列功能属性,因此常被用来探讨群落的构建机制与构建过程[6⁃7]㊂例如Freschet等在不同的森林类型中利用两种叶片性状即叶氮含量和比叶面积验证了生物和非生物因子共同驱动群落的构建[8]㊂有研究显示不同性状在相同空间尺度下的分布格局不同,例如:比叶面积在局域尺度下呈聚集分布时,而种子大小则表现为离散分布,表明不同功能性状对不同生态过程的响应不同[9⁃11]㊂此外,近年来生态学家试图将植物性状的分布格局与特定生境条件相联系[12],这对于森林群落的经营管理和多样性保护具有十分重要的意义㊂将群落功能性状的分布与局域尺度的环境因子联系起来,有助于检验出不同环境因子对性状分布格局的影响,并推测相应的生态学过程㊂本研究通过植物的叶面积㊁比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比和最大树高6类主要功能性状,以21.12hm2大面积森林固定监测样地群落调查数据为基础,分析了20mˑ20m空间尺度下的群落功能性状分布格局,并评估生态位过程和中性过程在吉林蛟河针阔混交林群落构建机制中的相对重要性㊂本研究同时结合样地内海拔高度㊁坡度㊁坡向和凹凸度四种地形因子,探讨不同地形因子对森林群落中性状分布的影响㊂研究旨在揭示吉林蛟河温带针阔混交林中功能性状的分布格局及其对生境变化的响应,并推测其背后的群落构建机制和构建过程㊂1㊀研究方法1.1㊀研究区概况与样地设置本研究区位于吉林省蛟河林业试验区管理局,地理位置为43ʎ57.897ᶄ 43ʎ58.263ᶄN,127ʎ42.789ᶄ 127ʎ43.310ᶄE㊂该气候属于受季风影响的温带大陆性气候,年均温为3.8ħ,最热月为7月,平均气温21.7ħ,最冷月为1月,平均气温-18.6ħ,年均降水量695.9mm㊂该地区林分类型为典型的次生针阔混交林,土壤类型为棕色森林土,土层厚度为20 100cm[13]㊂2009年建立面积为21.12hm2(660mˑ320m)的固定森林监测样地,样地最低海拔为425.3m,最高海拔为525.8m,相对最大高差为100.5m,地势变化较大㊂样地群落调查过程中,记录了样地内所有胸径(DBH)大于1cm的木本植物的物种名称㊁胸径㊁树高㊁冠幅和相对位置坐标,并挂牌标记进行长期监测㊂样地内主要树种有胡桃楸(Juglansmandshurica)㊁紫椴(Tiliaamurensis)㊁蒙古栎5972㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀(Quercusmongolica)㊁色木槭(Acermono)和红松(Pinuskoraiensis)等㊂图1㊀样地地形示意图Fig.1㊀Schematicdiagramoftheforestplot1.2㊀地形因子测量地形因子可以综合反映样地内土壤养分和水分状况,并在一定程度解释群落内植物的生长状况[14],本研究中我们采用海拔高度㊁凹凸度㊁坡度和坡向这四个地形因子记为环境因子㊂将整个研究区域划分为528个20mˑ20m的样方,采用Harm等人的计算方法计算地形因子[15]㊂具体计算过程为:每个样方4个顶点海拔值的平均值为该样方的平均海拔高度;目标样方的海拔减去与之相邻的8个样方的海拔平均值为该样方的凹凸度,其中,山脊为正值,洼地为负值;分别连接每个样方的3个顶点形成4个三角形平面,每个平面与该样方投影夹角的平均值为目标样方的坡度;4个平面与正北方向的角度平均值为目标样方的坡向[15⁃17]㊂基于该样地,已开展的相关研究表明,森林中不同的树种具有不同的生境偏好,树种的生长和分布都会受到地形因子的显著影响[13,18⁃19],并且有研究表明环境因子对树种分布的影响可能是通过物种所具有的功能性状而发挥作用[16]㊂但地形因子如何影响树种功能性状的分布格局目前仍然不十分清楚,有开展必要进一步的研究㊂1.3㊀功能性状样品的采集与测定本研究选择样地内胸径(DBH)大于5cm的乔木和大灌木树种作为研究对象,共计34个种㊂于2017年生长季采集并测量了样地内34个树种的6类关键功能性状数据,包括:叶面积㊁比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比和最大树高㊂叶片性状反映植物的生活史策略及其对环境条件的响应[20],叶面积反映植物对光的捕获能力,比叶面积反映植物在生长过程中的资源获取能力;叶氮含量表示植物在叶经济型谱上的位置,反映植物的资源获取能力[6];叶碳含量是植物生长过程中维持碳平衡的重要参数[21];叶碳氮比反映林木的碳氮代谢情况以及营养利用率㊂最大树高作为茎干性状的代表,反映植物光学生态位和综合竞争能力[16,22]㊂功能性状样品的采集和测定均参照Cornelissen等人的要求进行,常见种的采样株数在30株以上,部分稀有种的采样株数在10株以上[23];最大树高为样地内每个树种最高的前5%个体的平均值[17]㊂1.4㊀构建功能性状树在获取叶面积㊁比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比和最大树高的性状数据,首先对原始性状数据进行对数转化,然后通过主成分分析获取能够代表所有功能性状的主成分,选取其中综合解释量在90%以上的主成分作为综合功能性状因子,通过计算欧氏距离生成性状距离矩阵,采用最远距离法对性状距离矩阵进行系统聚类,生成功能性状聚类树[24]㊂1.5㊀计算功能性状分布格局本文计算了分别基于6种单个性状和一种综合性状的性状聚类,采用平均成对性状距离指数(meanpairwisetraitdistance;PW)和平均最近邻体性状距离指数(meannearestneighbortraitdistance;NN)计算样地内每个20mˑ20m的样方的功能性状分布格局[25⁃26]㊂这两种算法均需要通过性状聚类树随机转换实现,利用观察到的PW值和NN值量化群落性状分布结构,保持物种数量及个体数不变,将聚类树的末端树种随机转换999次,计算零模型下的功能性状分布,采用随机分布结果将观测值标准化,使我们获得标准条件下的PW和NN值即S.E.S.PW和S.E.S.NN,计算S.E.S.PW和S.E.S.NN的公式如下:S.E.S.PW=-1ˑ(PWact-mean(PWrand))/sd(PWrand)S.E.S.NN=-1ˑ(NNact-mean(NNrand))/sd(NNrand)式中,PWact代表实际观察到的平均成对性状距离,mean(PWrand)代表零模型随机转换999次物种后的成对性状距离模拟值的平均值,sd(PWrand)代表零模型模拟值的标准差;同理,NNact表示实际观察到的群落平均最近6972㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀邻体性状距离,mean(NNrand)代表零模型随机转换999次物种后的最近邻体性状距离模拟值的平均值,sd(NNrand)代表零模型模拟值的标准差㊂S.E.S.PW(或S.E.S.NN)值大于零代表功能性状呈聚集分布格局,小于零代表功能性状呈离散分布格局,等于零代表功能性状随机分布㊂1.6㊀功能性状分布对地形变化的响应本研究采用海拔高度㊁坡度㊁坡向和凹凸度这4个地形因子为环境变量,结合组合功能性状的分布格局,运用逐步回归的方法分析不同地形因子对综合功能性状格局的解释程度,探讨影响样地内群落构建的主要驱动因子㊂本研究相关计算均在统计软件R.3.5.1中完成,分别采用了 vegan ㊁ picante 软件包[27⁃28]㊂2㊀结果2.1㊀植物功能性状树本研究采用最远距离法进行系统聚类得到功能性状聚类树(图2),与直接反映植物进化关系的系统发育树不同,功能性状树主要反映了植物生态功能与生态策略之间的相似性(即性状与性状之间的相似性)㊂例如:本研究中山杨㊁白桦都属于先锋树种,具有较大的比叶面积㊁较高的叶氮含量和较低的木质密度,因此在功能性状树上的位置相近;色木槭㊁拧筋槭㊁糠椴㊁紫椴等属于硬阔树种,其叶片为单叶,具有较小的叶面积和较高的木质密度,因此在功能性状树上的位置相近;水曲柳㊁花曲柳和黄檗同属于硬阔树种,但其叶片为复叶,具有较大的叶面积和较高的木质密度,因此在功能性状树上的位置相近;而水榆花楸㊁稠李㊁山丁子㊁青楷槭㊁簇毛槭㊁千金榆㊁鼠李㊁黑樱桃㊁毛榛和花楸等灌木或小乔木则聚为一类,这一类树种的最大特征为其最大树高较小,是样地内典型的亚林层树种㊂此外,还有部分树种以其独特的性状特征单独划分为一类,例如胡桃楸(复叶树种)以其极大的叶面积,怀槐(豆科固氮树种)以其极高的叶氮含量等特征单独聚为一类,反映了此类树种在样地内独特的生态地位㊂2.2㊀植物功能性状分布格局基于平均成对性状距离(PW)的研究结果显示(表1;图3):单个性状中,叶面积的分布格局为离散分布(即图3中蓝色区域)多于聚集分布(即图3中红色区域),在全部528个样方中,有432个样方(81.8%)呈现为离散分布;除叶面积外,其余性状的分布格局均为聚集分布多于离散分布㊂基于平均最近邻体性状距离(NN)的研究结果显示(表1;图4):单个性状中,叶面积和最大树高分布格局为离散分布多于聚集分布,在全部528个样方中,分别有79.9%和60.0%的样方呈现为离散分布,而其余性状的分布格局为聚集分布多于离散分布(表1;图4)㊂总体而言基于平均最近邻体性状距离和平均成对性状距离的研究结果具有较高的一致性㊂此外,由6种单个性状组成的综合性状因子的分布格局同样为表现为聚集分布多于离散分布(表1;图5)㊂表1㊀性状呈聚集分布和离散分布的样方比例/%Table1㊀ProportionofaggregationanddispersionpatternsS.E.S.PWS.E.S.NN结构聚集(显著聚集)Aggregation(significant)结构分散(显著分散)Dispersion(significant)结构聚集(显著聚集)Aggregation(significant)结构分散(显著分散)Dispersion(significant)叶面积Leafarea18.2(1.5)81.8(25.6)20.1(0.8)79.9(22.7)比叶面积Specificleafarea98.1(27.3)1.9(0.0)73.5(25.2)26.5(0.0)叶氮含量Leafnitrogen92.8(7.8)7.2(0.0)89.0(4.9)11.0(0.0)叶碳含量Leafcarbon69.3(3.8)30.7(0.2)76.7(8.3)23.3(1.1)叶碳氮比Carbontonitrogenratio81.1(10.0)18.9(0.4)86.9(6.6)13.1(0.4)最大树高Maximumtreeheight61.4(9.7)38.6(5.3)40.0(4.5)60.0(23.3)综合性状因子Alltraits48.9(4.4)51.1(0.8)60.4(4.0)39.6(0.6)㊀㊀S.E.S.NN和S.E.S.PW分别表示PW和NN标准化效应值,括号数值内表示性状分布呈显著聚集或显著离散(P<0.05)的样方比例2.3㊀环境变量对功能性状分布格局的影响本研究利用综合性状因子计算了局域环境变量,包括海拔高度(ELE)㊁坡度(SLO)㊁坡向(ASP)和凹凸度7972㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀8972㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀图2㊀综合性状聚类树Fig.2㊀Clusteringtreebasedonfunctionaltraits(CON)对功能性状分布格局的相对解释程度㊂研究基于逐步回归的算法,结果发现局域环境变量对S.E.S.PW的解释量为37.4%,对S.E.S.NN的解释量为23.6%,其具体公式为:S.E.S.PW=0.380ELE+0.366SLO+0.088ASP;S.E.S.NN=0.306ELE+0.288SLO+0.074ASP,结果显示在已知的环境因子中,海拔高度㊁坡度和坡向对群落功能性状分布格局的影响显著,而凹凸度则对功能性状分布无显著影响,被排斥在回归模型之外㊂群落功能性状分布格局与局域环境变量显著相关,表明环境过滤在该研究区域内发挥着的重要作用㊂3㊀讨论3.1㊀平均成对性状距离指数和平均最近邻体性状距离指数平均成对性状距离指数(PW)和平均最近邻体性状距离指数(NN)都可以用来探讨群落功能性状的分布格局,这两种方法都是基于距离矩阵来量化群落功能性状的分布格局[29]㊂不同的是PW侧重于对整体群落的分析,是分析性状分布格局的基础[25],它用于衡量群落中所有物种的平均相关性[10],侧重于整个群体的平均值;而NN指数则是测量亲缘关系最近物种对之间的相关性[30],显示的是群落功能性状聚类树的终端性状分布格局㊂有研究认为这两种方法都具有检测环境过滤的作用,而NN指数更易检测相似性限制带来的影响[30]㊂PW指数和NN指数计算同一区域的性状分布格局,结果出现偏差,可能是性状在样地区域内分布不均造成的,本研究结果显示最大树高在不同计算方法下的结果存在差异,可能与该性状在样方内的分布不均匀有关[31]㊂图3㊀植物功能性状分布格局(基于平均成对性状距离法)Fig.3㊀Distributionpatternsofplantfunctionaltraits(basedonthemeanpairwisetraitdistance)红色表示性状聚集分布,蓝色表示性状离散分布,颜色越深表示性状聚集或离散分布的程度越明显图4㊀植物功能性状分布格局(基于平均最近邻体性状距离法)Fig.4㊀Distributionpatternsofplantfunctionaltraits(basedonthemeannearestneighbortraitdistance)红色表示性状聚集分布,蓝色表示性状离散分布,颜色越深表示性状聚集或离散分布的程度越明显9972㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀图5㊀综合性状因子的分布格局Fig.5㊀Distributionpatternsbasedonallsixtraits红色表示性状结构聚集,蓝色表示性状结构离散,颜色越深表示性状的聚集或离散程度越明显3.2㊀功能性状分布格局与群落构建机制已开展的相关研究表明,森林中不同的树种具有不同的生境偏好,树种的生长和分布都会受到地形因子的显著影响[13,18⁃19],并且有研究表明环境因子对树种分布的影响可能是通过物种所具有的功能性状而发挥作用[16]㊂因此,生态学家通过量化群落中功能性状的分布格局来推断影响群落构建的生态学机制㊂不同的功能性状反映了植物不同的生理㊁生态特征和生活史策略,因此其在群落中的分布格局以及其对生境变化的响应也会存在差异㊂环境过滤和相似性限制可以通过作用于不同类型的功能性状来影响群落中物种和性状的分布格局[32]㊂研究显示,环境会过滤掉不适生的性状,导致适生性状呈聚集状态[33],而离散的性状分布则是由功能性状之间的相似性限制造成,其与生态位分化有关㊂一般认为物种所具有的性状越相似,物种间的竞争就越激烈,而竞争导致某些具有相似性状的物种占据不同的生态位,使物种或性状的分布呈离散格局[34]㊂在此基础上,有研究认为与生产力相关的功能性状在小尺度上是聚集的,尤其是与叶的相对生长速率,叶片寿命和叶片的营养物质相关的性状[35]㊂本研究显示叶碳含量㊁叶氮含量和叶碳氮比等直接反映叶片营养物质的性状以及反映植物生长过程的比叶面积均呈显著聚集状态,综合性状结构同样呈显著聚集分布格局,与前人的研究结果较为一致[36⁃37]㊂研究显示竞争型性状在群落内容易呈离散分布的结构[26],而叶面积呈显著离散分布,可能与研究区域的林分类型有关㊂此外,最大树高在两种不同运算下结果有差异,该研究结果与Swenson等的研究类似,可能是由于最大树高在不同样方内分布不均,导致PW和NN计算结果不同[31]㊂本研究结果显示叶氮含量㊁叶碳含量和比叶面积等性状在群落中呈聚集分布,反映了环境过滤在群落构建中的重要作用,该结果与温带和热带地区的研究结果类似[9],但组合性状分布结构显示,虽然呈显著聚集结构的样方较多,但样地中也有很多有离散趋势的样方,这表明除环境过滤外,生物之间的相互作用对该地区的群落构建也有影响,非随机过程是影响该地区群落构建的主要驱动因子[38]㊂3.3㊀地形因子与群落功能性状分布本文从综合功能性状的角度,结合局域的地形因子,利用不同的功能性状结构指数,对两种计算结果回归分析显示,地形因子对性状分布格局影响显著㊂有研究指出,在环境条件较差的区域,例如低海拔干旱和高海拔霜冻地区,由于环境过滤作用的影响,群落的功能性状结构通常呈聚集分布,而在环境条件适宜的低海拔地区,生物间的竞争作用强烈,而环境过滤的作用并不明显,因此群落的功能性状结构多呈离散分布[39]㊂性状分布呈现的这种差异既来自生态位保守的影响,也是性状对每个特定的环境条件的适应性反应[40]㊂在我们的研究区域内,如图5综合性状分布格局显示,样地东部和西北部海拔较高处的功能性状格局多呈聚集分布,样地中部多呈离散分布,从图1的地形图中可以看出,聚集分布多处于上坡带,海拔高度和地面凹凸度均有较大幅度的改变,而离散分布多处于地势低平㊁环境条件较为温和的区域㊂凹凸度㊁海拔高度㊁坡度和坡向这四种地形因子与综合性状结构逐步回归分析显示,海拔和坡度坡向对群落性状分布具有显著影响,凹凸度的影响很小,基本忽略不计,而在Liu等人[16]的研究中,发现海拔高度和地0082㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀形凹凸度对性状分布的影响最大,研究结果的差异可能与研究地的地形条件不同有关㊂有研究指出,海拔较高的区域,温度和大气压降低,太阳辐射增加,可以影响植物的生长进程[41]㊂样地地形图显示,整块样地的地形变化差异显著,将样地一分为二,明显东部地区海拔高,西部地区海拔趋于平缓,样地生境存在明显的异质性,这种生境异质性很大程度上影响了资源的利用情况[42],并进一步影响了功能性状的分布格局㊂在这种地形条件下,坡度能够显著影响到植物对土壤水分㊁养分以及光照等资源的获取与利用,从而对植物的性状进行选择和过滤,进而影响了群落的物种和性状分布格局㊂4㊀结论本研究从植物功能性状的角度出发,结合环境因子,探讨影响蛟河地区群落构建的主要驱动机制㊂研究表明,不同性状的分布格局不同,比叶面积㊁叶碳含量㊁叶氮含量㊁叶碳氮比在研究区域内呈明显的聚集分布,表示环境过滤对物种的功能性状分布格局具有重要作用;叶面积呈明显的离散分布,表示相似性限制的重要作用;最大树高在不同计算方法下的分布结果有差异㊂研究显示,环境因子对性状分布具有显著影响,其中海拔高度的影响最为显著,表明环境过滤的重要作用㊂综上所述,本研究认为非随机过程是蛟河针阔混交林群落构建的主要驱动因子,环境过滤和生物之间的相似性限制是群落内物种和功能性状分布格局的主要决定因素㊂参考文献(References):[1]㊀WeiherE,ClarkeGDP,KeddyPA.Communityassemblyrules,morphologicaldispersion,andthecoexistenceofplantspecies.Oikos,1998,81(2):309⁃322.[2]㊀SiefertA,RavenscroftC,WeiserMD,SwensonNG.Functionalbeta⁃diversitypatternsrevealdeterministiccommunityassemblyprocessesineasternNorthAmericantrees.GlobalEcologyandBiogeography,2013,22(6):682⁃691.[3]㊀EtienneRS,AlonsoD.Adispersal⁃limitedsamplingtheoryforspeciesandalleles.EcologyLetters,2005,8(11):1147⁃1156.[4]㊀RosindellJ,HubbellSP,EtienneRS.Theunifiedneutraltheoryofbiodiversityandbiogeographyatageten.Trendsinecology&evolution,2011,26(7):340⁃348.[5]㊀柴永福,岳明.植物群落构建机制研究进展.生态学报,2016,36(15):4557⁃4572.[6]㊀周道玮.植物功能生态学研究进展.生态学报,2009,29(10):5644⁃5655.[7]㊀孟婷婷,倪健,王国宏.植物功能性状与环境和生态系统功能.植物生态学报,2007,31(1):150⁃165.[8]㊀FreschetGT,DiasATC,AckerlyDD,AertsR,VanBodegomPM,CornwellWK,DongM,KurokawaH,LiuGF,OnipchenkoVG,OrdoñezJC,PeltzerDA,RichardsonSJ,ShidakovII,SoudzilovskaiaNA,TaoJP,CornelissenJHC.Globaltocommunityscaledifferencesintheprevalenceofconvergentoverdivergentleaftraitdistributionsinplantassemblages.GlobalEcologyandBiogeography,2011,20(5):755⁃765.[9]㊀SwensonNG,EnquistBJ.Ecologicalandevolutionarydeterminantsofakeyplantfunctionaltrait:wooddensityanditscommunity⁃widevariationacrosslatitudeandelevation.AmericanJournalofBotany,2007,94(3):451⁃459.[10]㊀WebbCO.Exploringthephylogeneticstructureofecologicalcommunities:anexampleforrainforesttrees.AmericanNaturalist,2000,156(2):145⁃155.[11]㊀房帅,原作强,蔺菲,叶吉,郝占庆,王绪高.长白山阔叶红松林木本植物系统发育与功能性状结构.科学通报,2014,59(24):2342⁃2348.[12]㊀AndersonMJ,CristTO,ChaseJM,VellendM,InouyeBD,FreestoneAL,SandersNJ,CornellHV,ComitaLS,DaviesKF,HarrisonSP,KraftNJB,StegenJC,SwensonNG.Navigatingthemultiplemeaningsofβdiversity:aroadmapforthepracticingecologist.EcologyLetters,2011,14(1):19⁃28.[13]㊀郝珉辉,张忠辉,赵珊珊,张春雨,赵秀海.吉林蛟河针阔混交林树木生长的空间关联格局.生态学报,2017,37(6):1922⁃1930.[14]㊀WestobyM,FalsterDS,MolesAT,VeskPA,WrightIJ.Plantecologicalstrategies:someleadingdimensionsofvariationbetweenspecies.AnnualReviewofEcologyandSystematics,2002,33:125⁃159.[15]㊀HarmsKE,ConditR,HubbellSP,FosterRB.Habitatassociationsoftreesandshrubsina50⁃haneotropicalforestplot.JournalofEcology,2001,89(6):947⁃959.[16]㊀LiuJJ,TanYH,SlikJWF.Topographyrelatedhabitatassociationsoftreespeciestraits,compositionanddiversityinaChinesetropicalforest.ForestEcologyandManagement,2014,330:75⁃81.1082㊀7期㊀㊀㊀夏梦洁㊀等:吉林蛟河针阔混交林功能性状分布格局及其对地形因素的响应㊀2082㊀生㊀态㊀学㊀报㊀㊀㊀41卷㊀[17]㊀郝珉辉,李晓宇,夏梦洁,等.抚育采伐对蛟河次生针阔混交林功能结构和谱系结构的影响.林业科学,2018,54(5):1⁃9.[18]㊀ZhangCY,ZhaoYZ,ZhaoXH,VonGadowK.Species⁃habitatassociationsinanortherntemperateforestinChina.SilvaFennica,2012,46(4):501⁃519.[19]㊀郝珉辉,张忠辉,赵珊珊,赵秀海,叶尔江㊃拜克吐尔汉,张春雨.吉林蛟河针阔混交林树木生长与生境的关联性.生态学报,2017,37(10):3437⁃3444.[20]㊀WrightIJ,ReichPB,WestobyM,AckerlyDD,BaruchZ,BongersF,Cavender⁃BaresJ,ChapinT,CornelissenJHC,DiemerM,FlexasJ,GarnierE,GroomPK,GuliasJ,HikosakaK,LamontBB,LeeT,LeeW,LuskC,MidgleyJJ,NavasML,NiinemetsÜ,OleksynJ,OsadaN,PoorterH,PootP,PriorL,PyankovVI,RoumetC,ThomasSC,TjoelkerMG,VeneklaasEJ,VillarR.Theworldwideleafeconomicsspectrum.Nature,2004,428(6985):821⁃827.[21]㊀BoernerREJ.Foliarnutrientdynamicsandnutrientuseefficiencyoffourdeciduoustreespeciesinrelationtositefertility.JournalofAppliedEcology,1984,21(3):1029⁃1040.[22]㊀MolesAT,WartonDI,WarmanL,SwensonNG,LaffanSW,ZanneAE,PitmanA,HemmingsFA,LeishmanMR.Globalpatternsinplantheight.JournalofEcology,2009,97(5):923⁃932.[23]㊀CornelissenJHC,LavorelS,GarnierE,DíazS,BuchmannN,GurvichDE,ReichPB,TerSteegeH,MorganHD,VanDerHeijdenMGA,PausasJG,PoorterH.Ahandbookofprotocolsforstandardisedandeasymeasurementofplantfunctionaltraitsworldwide.AustralianJournalofBotany,2003,51(4):335⁃380.[24]㊀PetcheyOL,GastonKJ.Functionaldiversity(FD),speciesrichnessandcommunitycomposition.EcologyLetters,2002,5(3):402⁃411.[25]㊀LiuXJ,SwensonNG,ZhangJL,MaKP.Theenvironmentandspace,notphylogeny,determinetraitdispersioninasubtropicalforest.FunctionalEcology,2013,27(1):264⁃272.[26]㊀WeiherE,KeddyPA.Assemblyrules,nullmodels,andtraitdispersion:newquestionsfromoldpatterns.Oikos,1995,74(1):159⁃164.[27]㊀OksanenJ,GuillaumeBF,KindtR,LegendreP,MinchinPR,OᶄHaraRB,SimpsonGL,SolymosP,StevensMHH,WagnerH.Vegan:communityecologypackage.Rpackageversion2.0⁃9,2013,2(9):1⁃295.http://vegan.r⁃forge.r⁃project.org.[28]㊀KembelSW,CowanPD,HelmusMR,CornwellWK,MorlonH,AckerlyDD,BlombergSP,WebbCO.Picante:Rtoolsforintegratingphylogeniesandecology.Bioinformatics,2010,26(11):1463⁃1464.[29]㊀SwensonNG,EnquistBJ.Opposingassemblymechanismsinaneotropicaldryforest:implicationsforphylogeneticandfunctionalcommunityecology.Ecology,2009,90(8):2161⁃2170.[30]㊀KraftNJB,CornwellWK,WebbCO,AckerlyDD.Traitevolution,communityassembly,andthephylogeneticstructureofecologicalcommunities.TheAmericanNaturalist,2007,170(2):271⁃283.[31]㊀SwensonNG,EricksonDL,MiXC,BourgNA,Forero⁃MontañaJ,GeXJ,HoweR,LakeJK,LiuXJ,MaKP,PeiNC.Phylogeneticandfunctionalalphaandbetadiversityintemperateandtropicaltreecommunities.Ecology,2012,93(sp8):S112⁃125.[32]㊀IngramT,ShurinJB.Trait⁃basedassemblyandphylogeneticstructureinnortheastpacificrockfishassemblages.Ecology,2009,90(9):2444⁃2453.[33]㊀ÅgrenGI,FagerströmT.Limitingdissimilarityinplants:randomnesspreventsexclusionofspecieswithsimilarcompetitiveabilities.Oikos,1984,43(3):369⁃375.[34]㊀MacArthurRH.Populationecologyofsomewarblersofnortheasternconiferousforests.Ecology,1958,39(4):599⁃619.[35]㊀黄建雄,郑凤英,米湘成.不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响.植物生态学报,2010,34(3):309⁃315.[36]㊀ReichPB,UhlC,WaltersMB,EllsworthDS.Leaflifespanasadeterminantofleafstructureandfunctionamong23Amazoniantreespecies.Oecologia,1991,86(1):16⁃24.[37]㊀ReichPB,WaltersMB,EllsworthDS.Fromtropicstotundra:globalconvergenceinplantfunctioning.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1997,94(25):13730⁃13734.[38]㊀LakeJK,OstlingA.Commenton"Functionaltraitsandniche⁃basedtreecommunityassemblyinanAmazonianforest".Science,2009,324(5930):1015⁃1015.[39]㊀KlugeJ,KesslerM,KlugeJ.Phylogeneticdiversity,traitdiversityandniches:speciesassemblyoffernsalongatropicalelevationalgradient.JournalofBiogeography,2011,38(2):394⁃405.[40]㊀Diniz⁃FilhoJAF,BiniLM,RodríguezMÁ,RangelTFLVB,HawkinsBA.Seeingtheforestforthetrees:partitioningecologicalandphylogeneticcomponentsofBergmannᶄsruleinEuropeanCarnivora.Ecography,2007,30(4):598⁃608.[41]㊀KörnerC.Theuseof altitude inecologicalresearch.TrendsinEcology&Evolution,2007,22(11):569⁃574.[42]㊀GoldbergD,NovoplanskyA.Ontherelativeimportanceofcompetitioninunproductiveenvironments.JournalofEcology,1997,85(4):409⁃418.。
距离和密度制约对吉林蛟河阔叶红松林幼苗生长的影响

距离和密度制约对吉林蛟河阔叶红松林幼苗生长的影响
吉林蛟河阔叶红松林是中国东北地区重要的生态系统之一,阔叶红松林幼苗的生长状况对于该地区的生态环境保护和可持续发展有着至关重要的作用。
在阔叶红松林幼苗生长过程中,距离和密度制约因素对其生长发育起着重要的影响,下文将对此进行探讨。
首先,距离制约对于幼苗的生长影响很大。
由于距离影响了幼苗之间的竞争和相互作用,合理的距离安排可以促进其生长发育。
在阔叶红松林幼苗的生长过程中,苗木相互之间的距离过于接近会大大限制其生长发育,因为它们需要争夺土壤养分、水分和光线。
过于紧密的种植会使得苗木之间存在极大的竞争,以至于受到抑制。
但是,过度的间距也会对幼苗的生长造成不良的影响。
因为过大的间距会导致土壤水分和其他资源的散失,使得幼苗的生长发育受到限制,难以实现良好的生长状态。
其次,密度制约也是影响幼苗生长的重要因素。
幼苗的密度会直接影响它们之间的相互作用和竞争状况,进而影响其生长速率和生长状态。
阔叶红松林幼苗的种植密度过高会使得整个林区内的生态环境不平衡,导致不良的生长效果。
因此,合理的密度安排可以改善苗木的生长状况,提高其生长发育的质量与速率。
在这方面,可以通过适当调整栽植密度、减少植物种类的数量等方式来解决这个问题。
总体而言,距离和密度制约因素对于吉林蛟河阔叶红松林幼苗的生长发育有着很大的影响。
采取恰当的距离和密度安排方法,可以改善幼苗的生长质量和速率,同时提升阔叶红松林的生态
环境。
这样可以确保吉林蛟河阔叶红松林获得可持续生态发展,成为一个更加繁荣的地区。
吉林蛟河红松阔叶林群落结构特征及其生态位研究
吉林蛟河红松阔叶林群落结构特征及其生态位研究摘要:本文通过对吉林省蛟河市红松阔叶林群落的调查和研究,分析了其群落结构特征及其生态位,探讨了其在生态系统中的重要性。
研究结果表明,该红松阔叶林群落中树层植物物种丰富度高,垂直结构明显,主要树种为红松、榆树和白蜡等。
灌木层和草本层植物分别由18种和25种组成,种类较为丰富。
群落的物种多样性指数较高,达到了3.12。
群落中优势种为红松,其相对重要值为42.8%,其他主要物种包括榆树和白蜡等。
该红松阔叶林群落在生态系统中具有重要的生态位。
它不仅对维持生态系统的物质循环、水循环和能量流动等起着重要作用,还对生态系统的稳定性和完整性有着重要的影响。
研究结论表明,为了保护和利用该红松阔叶林群落,需要采取相应的保护措施和管理措施,加强群落多样性保护、物种保护和栖息地保护等方面的工作。
关键词:吉林省;蛟河市;红松阔叶林群落;群落结构;生态位Abstract:The study aimed to analyze the community structure and ecological niche of the red pine broad-leaved forest community in Jiaohe City, Jilin Province, and explore its importance in the ecosystem. The study found that the red pine broad-leaved forest community had a high tree species richness, obvious vertical structure, and the dominant tree species were red pine, elm, and white wax. The shrub layer and herb layer were composed of 18 and 25 species respectively. The species diversity index of the community was high, reaching 3.12, and the dominant species was red pine, with a relative importance value of 42.8%.The red pine broad-leaved forest community played an important ecological role in maintaining the material cycling, water cycling, and energy flow of the ecosystem, and had a significant impact on thestability and integrity of the ecosystem.The findings of the study suggest that appropriate conservation and management measures should be taken to protect and utilize the red pine broad-leavedforest community. Protection of community diversity, species, and habitat is necessary to maintain the community’s ecological niche and ecosystem service.Keywords: Jilin Province; Jiaohe City; red pine broad-leaved forest community; community structure; ecological nicheThe red pine broad-leaved forest community plays an essential role in sustaining and supporting the ecological system in Jiaohe City, Jilin Province. The community structure encompasses various vegetation types, including trees, shrubs, and herbs, that serve as a habitat for many animal species. The study found that the forest community has a high degree of species diversity and richness, with a total of 89 plant species identified in the area.It is evident that the ecological niche of the community is critical in maintaining the integrity of the ecosystem through water conservation, soil stability, carbon sequestration, and nutrient cycling. However, human activities, such as overgrazing, deforestation, and agricultural expansion, have adversely impacted the red pine broad-leaved forest community.In conclusion, it is essential to implement appropriate conservation and management measures to protect the red pine broad-leaved forest community.Conserving the biodiversity of the community will help sustain the local ecosystem, contributing to broader efforts to mitigate global environmental challenges such as climate change. More comprehensive research is also necessary to understand better the community's ecological niches and inform conservation actions for the long-term protection of the ecosystemFurthermore, there are several challenges that need to be addressed when implementing the conservation and management measures for the red pine broad-leavedforest community. One of the main challenges is the fragmentation of the forest due to human activities such as land-use change, agriculture, and urbanization. This fragmentation has resulted in the loss of habitat and connectivity, which in turn affects the movement and gene flow of the forest species.To address this challenge, it is essential toestablish protected areas around the red pine broad-leaved forest community. Protected areas can serve as a refuge for the forest species, allowing them tothrive and maintain their genetic diversity. Additionally, buffer zones can be established around the protected areas to minimize the impact of human activities on the forest ecosystem.Another challenge is the invasion of non-native species, which can outcompete and displace native species, leading to the loss of biodiversity. Non-native species such as Japanese knotweed and garlic mustard have been found to invade the red pine broad-leaved forest community, which can have detrimental effects on the forest species.To address this challenge, it is important to implement invasive species management strategies, such as early detection and rapid response, mechanical and chemical control, and ecological restoration. These measures can effectively control the spread and reduce the impact of non-native species on the forest ecosystem.In conclusion, the red pine broad-leaved forest community is a unique and valuable ecosystem that requires appropriate conservation and management measures to protect its biodiversity. The establishment of protected areas, the implementation of invasive species management strategies, and the conduct of comprehensive research are essential to sustain the forest ecosystem and mitigate global environmental challenges. By working together, we can ensure the long-term protection of the red pine broad-leaved forest community and promote a sustainable future for our planetBeside the aforementioned conservation and management strategies, there are other important measures that can be taken to sustain the red pine broad-leaved forest community. One of these measures is promoting sustainable forestry practices that promote biodiversity conservation while also supporting the economic livelihoods of local communities.Sustainable forestry practices that promote biodiversity conservation can be achieved through the implementation of selective logging methods, which involve the selective harvest of only mature trees, rather than clear-cutting entire areas. This approach allows for the maintenance of critical tree canopy cover, which can support the growth, reproduction, and habitat needs of other plant and animal species.Another important measure is community-based conservation, which involves the active participation of local communities in conservation efforts. This approach recognizes the importance of local knowledge and experience in the protection of the forest ecosystem and ensures that the needs of local communities are taken into consideration when planningand implementing conservation measures.Finally, the promotion of ecotourism can also play an essential role in forest conservation by providing economic incentives for the protection of the forest ecosystem. Ecotourism can generate revenue for local communities, which can be used to support conservation measures and sustain the long-term viability of the forest ecosystem.In conclusion, the red pine broad-leaved forest community is a critical ecosystem that requires appropriate conservation and management measures to protect its biodiversity. The implementation of protected areas, invasive species management strategies, sustainable forestry practices, community-based conservation, and ecotourism can all work together to ensure the long-term protection of this unique and valuable ecosystem. By working together, we can create a sustainable future for our planet, where biodiversity is conserved, local communities are supported, and the natural beauty of our world is preserved for future generationsIn conclusion, protecting the biodiversity of tropical rainforests is crucial for the sustainability of our planet's ecosystems. Through the implementation ofvarious conservation measures, we can achieve long-term protection of these valuable ecosystems. It is important that we work together to preserve the natural beauty of our world and support the local communities who depend on these forests for their livelihoods. By taking action now, we can ensure a brighter future for generations to come。
红松次生混交林中针叶树与阔叶树的结构差异
红松次生混交林中针叶树与阔叶树的结构差异李远发;明安刚;叶绍明;吕成群;惠刚盈【摘要】红松混交林是中国东北地区最重要的植被类型,在缓解全球气候变化、保证木质产品供给和维护生物多样性中扮演重要角色.该研究以吉林东大坡自然保护区第52林班的水曲柳红松混交林和第54林班的核桃楸-红松混交林为例,采用林分结构参数一元分布(UDSSP)、二元分布(BDSSP)和非线性模型(NLM)分析了它们的针叶树和阔叶树的结构特征,以探讨混交林中针、阔树种组之间的关系,为低效次生林的改造提供参考.结果显示:(1)多数针叶树同时处于高混交和优势状态,仅有极少数个体处于中庸或劣势;多数阔叶树亦处于高混交状态,且不同优势的树木总数几乎相等.(2)大部分树木都同时处于高度混交和随机分布状态,少量针叶树略偏向规则分布.(3)大部分针叶树同时处于随机分布和优势状态,且不同优势的阔叶树皆倾向随机分布,而处于规则或聚集分布的数量很少.(4)针叶树和阔叶树的径阶分布均能够较好地通过指数函数或幂函数描述.这些特征充分展示了空间格局和径阶分布在不同群落类型之间差异很小,但在不同树种组之间存在明显差异.%Korean pine mixed forest is one of the most important vegetation in northeast China.It plays an important role in alleviating global climate change,guaranteeing the wooden product supply and maintaining biological diversity.Four plots located at the 52thand 54th compartments,Dongdapo nature preserve,Jiaohe,Jilin province,were analyzed by using the univariate distribution (UDSSP) and bivariate distribution (BDSSP) of stand structural parameters and nonlinear model (NLM),with the purpose to clear the relationship between coniferous and hardwood species,which may contribute to the reconstruction of lowproduct secondary forests.Thesestands stood for Fraxinus mandschurica Rupr.and Pinus koraiensis Sieb et Zucc.mixed forest and Juglans mandshurica Maxim.and P.koraiensis mixed forest,respectively.The results showed that:(1) Most conifers were in a state of high mixture and dominance,and only few belonged to the middle or inferior plants;the majority of hardwoods were also intensely surrounded by other species and the ones with different dominance showed an equal number;(2) Many individuals were intensely mixed and randomly distributed at the same time,the slightly difference was conifers biased towards regularity when compared with hardwoods;(3) Most conifers were randomly distributed and had advantage on sizes,while few medium or inferior trees had the same pattern.Hardwoods with different dominance were randomly distributed and only few were regular or clump;(4) Their diameter classes were well expressed by power function or exponential functions.These features revealed that spatial patterns and diameter distributions were closely similar within different types of forest stand and differed from conifers to hardwood species groups.【期刊名称】《西北植物学报》【年(卷),期】2017(037)008【总页数】12页(P1645-1656)【关键词】混交林;空间结构;径阶分布;树种组;一元分布;二元分布【作者】李远发;明安刚;叶绍明;吕成群;惠刚盈【作者单位】广西大学林学院,南宁530004;中国林业科学研究院热带林业实验中心,广西凭祥532600;广西大学林学院,南宁530004;广西大学林学院,南宁530004;中国林业科学研究院林业研究所,北京100091【正文语种】中文【中图分类】Q948.15+7红松混交林是北半球温带地区的典型地带性植被,广泛分布在中国东北东部、俄罗斯远东南部、朝鲜半岛以及日本岛的四国和本州(124°~140°20′E,33°50′~52°N),总面积约500 000 km2[1-2],占据北半球森林面积和蓄积的很大比例,并在缓解全球气候变化、木材供给、生物多样性保存、生态效益发挥中扮演重要角色。
吉林蛟河天然阔叶红松林草本植物多样性及其与土壤因子的关系
吉林蛟河天然阔叶红松林草本植物多样性及其与土壤因子的关系龚艳宾;郭建斌;赵秀海;于诗卓【期刊名称】《浙江农林大学学报》【年(卷),期】2016(033)004【摘要】于2014年生长季(5-9月)对设置在天然阔叶红松Pinus koraiensis 林样地内的272个草本样方(1 m×1 m)进行物种及其植株高度、盖度以及土壤因子进行调查,应用Shannon-Wiener指数、Simpson指数、Gleason指数及相应的均匀度(Pielou指数)和生态优势度等指标分析天然阔叶红松林样地内草本植物物种多样性的生长期动态,并采用冗余分析方法(RDA)探讨了土壤因子对草本层多样性格局的影响。
结果表明:1样地内草本植物物种组成丰富,共有91种,隶属于34科80属,以毛茛科Ranunculaceae和百合科Liliaceae的物种数最多,物种单科单属、单属单种现象很明显,其中单种属占总属数的90%以上,在样地内占有绝对优势。
2草本植物物种组成及多样性月季变化较明显,物种Gleason指数、Simpson指数和Shannon-Wiener指数的月季变化趋势一致,在6月达到最大值,分别为13.727,0.924和3.093,9月取得最低值;Pielou指数也呈先增加后减小的变化趋势,在7月达到最大,9月最小。
生态优势度指数从5月到9月逐月递增,表明草本植物在7月物种分配最为均匀,9月优势种最为突出。
3冗余分析(RDA)排序图第1轴主要反映了土壤全钾质量分数与土壤pH值的变化趋势,RDA第2轴主要反映的是土壤速效磷质量分数的变化趋势,表明这些因子对草本植物多样性格局具有明显的影响。
通过向前选择和蒙特卡洛(Monte Carl permutation test)检验分析也发现其中具有显著性影响作用的是土壤全钾质量分数和p H值(P=0.002)。
【总页数】9页(P620-628)【作者】龚艳宾;郭建斌;赵秀海;于诗卓【作者单位】[1]北京林业大学水土保持学院,北京100083;[2]北京林业大学林学院,北京100083【正文语种】中文【中图分类】S718.5【相关文献】1.吉林蛟河天然阔叶红松林下5种灌木生物量估算模型 [J], 杨松;侯继华;赵秀海;魏丽萍;李颖;姚静;代海军;何怀江2.吉林蛟河阔叶红松林地表甲虫多样性时间动态分析 [J], 刘生冬;孟庆繁;高文韬;李燕;赵红蕊;李天琦3.吉林蛟河阔叶红松林样地种-面积关系 [J], 陈桂莲;谭凌照;范春雨;张新娜;张春雨;赵秀海4.长白山原始阔叶红松林林下草本植物多样性格局及其影响因素 [J], 夏富才;潘春芳2;赵秀海2;何海燕3;周海城35.我国沙拐枣属(Calligonum L.)天然群落物种多样性与土壤因子的耦合关系 [J], 谭勇;张强;潘伯荣;康晓珊因版权原因,仅展示原文概要,查看原文内容请购买。
2种类型阔叶红松林优势种群空间分布格局及其关联性
2种类型阔叶红松林优势种群空间分布格局及其关联性赵中华;惠刚盈;胡艳波;周红敏;张连金【摘要】以吉林蛟河2块1 hm2阔叶红松林样地全林每木定位数据为基础,运用基于RipleyK (d)函数和Mark相关函数的O-ring统计分析方法,分析了2块样地中红松种群与主要优势种群林分中层与林分上层空间分布格局及树种的空间关联性,结果表明:水曲柳红松林中的红松种群和水曲柳种群在小尺度上为聚集分布的趋势,林分中层的红松种群尤为明显,椴树红松林中的红松和椴树种群在多个尺度上呈聚集分布.2个类型阔叶红松林中相同林层中红松种群与水曲柳种群、椴树种群总体上呈现不相关的空间关系,但不同林层的种群间存在着一定的相关关系.水曲柳红松林中,林分上层中的红松与林分中层的红松在0~19 m的尺度内存在显著地负相关,在椴树红松林中不明显;林分上层水曲柳与林分中层水曲柳、林分上层椴树与林分中层椴树在小尺度上存在明显地负相关关系;林分上层的红松与林分中层的水曲柳及椴树都在较大尺度上存在显著负相关的空间关系,而林分中层的红松与林分上层的水曲柳、椴树的空间相关关系为不相关.%Based on the location data for each timber from two broadleaved Korean pine ( Pinus koraiensis) mixed forest plots which area is 1 hm2 at Jiaohe of Jilin province, the spatial distribution pattern and tree species spatial correlation of P. koraiensis and main dominate population in middle and upper vertical forest layer were chosen as study objects, the O-ring statistics based on RipleyK(d) function and Mark correlation function was used to analyze the spatial distribution and spatial correlation. The result showed that spatial distribution pattern of P. koraiensis and Fraxinus mandshurica were clumped at smaller scales in F. mandshurica-P. koraiensis forest, P. koraiensis was es-pecially inmiddle vertical layer. P. koraiensis and Tilia amurensis spatial distribution pattern were clumped at mul-tiple scales. At same vertical layer, P. koraiensis had no significant spatial association to F. mandshurica and T. amurensis, however, there had some association in different vertical layer of different population. P. koraiensis in upper layer has significant negative correlation with midstory layer at 0 ~ 19 m, this properties was not obviously in T. amurensis-P. koraiensis mixed forest. F. mandshurica and T. amurensis in upper vertical layer showed signifi-cant negative correlation with midstory layer at smaller scales. P. koraiensis in upper layer shower significant nega-tive correlation with F. mandshurica and T. amurensis in middle vertical layer lager scale, but P. koraiensis in mid-dle vertical layer had no correlation with F. mandshurica and T. amurensis in upper layer.【期刊名称】《林业科学研究》【年(卷),期】2011(024)005【总页数】9页(P554-562)【关键词】水曲柳红松林;椴树红松林;空间分布格局;空间关联性【作者】赵中华;惠刚盈;胡艳波;周红敏;张连金【作者单位】中国林业科学研究院林业研究所,北京100091;国家林业局林木培育重点实验室,北京100091;中国林业科学研究院林业研究所,北京100091;国家林业局林木培育重点实验室,北京100091;中国林业科学研究院林业研究所,北京100091;国家林业局林木培育重点实验室,北京100091;浙江省龙泉市林业科学研究所,浙江龙泉323700;中国林业科学研究院林业研究所,北京100091;国家林业局林木培育重点实验室,北京100091【正文语种】中文【中图分类】S718.54;S791.247种群是组成群落和生态系统的基本成分,种群内的所有个体组成有一定的规律性。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第 期
胡艳波等: 吉林蛟河天然红松阔叶林的空间结构分析
!!
式 ( ) 为:
其中: !
当参照树 {, , 否则
与第 株相邻木非同种时
( )
公 大小比数是指胸径、 树高或冠幅等大于参照树的相邻木占 株最近相邻木的株数比例, 如果参照树 {, , 否则 比相邻木 小
!
其中:
( )
用来描述相邻树木围绕参照树 的均匀性。任意两个邻接最近相邻木的夹角 角尺度 ( ) 有两个, 小角为", 最近相邻木均匀分布时的夹角设为标准角" 。角尺度 ( ) 被定义为" 角小 于标准角" 的个数占所考察的 个夹角的比例。表达式为: , 当第 个" 角小于标准角" , 否则 根据 ( ) 、 ) ( 和 ( ) 式中各参数的定义, 得出参数取值的含义如图 所示。
的现象 ( 株单木周围有 株相邻木属于其它树种, ) 。所有树种零度混交和轻度混 !" 交比例之和都不超过 。蒙古栎、 枫桦、 黄榆、 红松、 黄波罗和鱼鳞云杉等树种零度和轻度 混交都没有。在主要树种中, 除蒙古栎单木有 不超过 度混交的比例合计超过了 属于中度混交, 其它树种中度混交的比例 , 红松的强度和其强 , 也就是说, 各树种强度和极强度混交的比例合计都超过
传统林学中对于植被结构的研究, 主要集中在描述群落的树种组成和空间分布、 群落成层 性、 年龄组成、 生物量分配等。随着近自然森林经营的兴起, 森林的结构、 过程和诸多关系等详 细信息越来越成为森林经营的前提, 其中涉及单木之间空间关系的林分空间结构越来越受到 人们的重视。林分空间结构是指林木在林地上的分布格局及其属性在空间上的排列方式, 也 就是林木之间树种、 大小、 分布等空间关系。林分空间结构决定了树木之间的竞争势及其空间
, 黄波罗和鱼鳞云杉等甚至全都是强度和极强度混交。
第 期
胡艳波等: 吉林蛟河天然红松阔叶林的空间结构分析
图
几个主要树种的混交度分布
!"#
林木大小分化程度 天然林树种众多, 大小不同的各树种林
木错落地分布在一起, 分析各个树种大小比 数分布, 以此判断各树种林木的大小分化程 度。图 表现了 块样地中的一些珍贵针、 阔叶树种的大小比数分布。 根据大小比数的定义, !" 越大代表相邻 木越 大, 而 参 照 树 越 不 占 优 势。从 上 图 可 见, 针叶树种处于中庸状态的情况较少, 处 于优势的情况较多。也就是说, 针叶树大多
!
其中:
{
( )
图
结构参数取值及含义
以上概念都是针对一个空间结构单元而言的, 在分析整个林分的空间结构时, 需要计算林 分内所有结构单元的参数平均值, 并将其作为分析的基础。其中, 通过分析林分平均混交度和
[ ] 各树种混交度 , 研究天然林树种空间配置情况; 通过分析各树种大小比数研究该树种在林分 [ ] [ ] 内的生长状况 ; 通过分析样地平均角尺度研究林木水平地面上的分布格局 。
红松阔叶林空间结构分析
!"# 林木种间关系
林
业
科
学
研
究
第
卷
天然林树种丰富, 搭配复杂。图 展示了
块样地的林木平均混交度的分布。在每一块
样地中 ! 从 到 的各取值的比例逐渐增大, 这说明在这些林分里同树种聚集在一起的情况 为数不多, 多数树种不与本树种为伴。总的来看, 块样地的平均混交度都很高, 最高的平均 混交度甚至接近 。这说明天然林经营和保护的越好, 树种孤立的情况越少, 树种之间的 隔离程度越小, 树种组成的结构越多样, 林分的稳定性也会相应增强。
图 固定样地林木定位图
固定样地, 其调查数据用于计算所有结构参数。辅助分析样地 的临时样地, 其调查数据用于计 算混交度和大小比数。
描述天然林的空间结构: ( ) 树种的空间隔离程度, 或者说天然林的树种组成和空间配置情况 (描 述非同质性) ( ;) 林木个体大小分化程度, 或者说是树种的生长优势程度 (描述非均一性) ( ) ; 林
研究方法
!"# ( 调查方法 本研究采用了全面调查法, 调查样地内所有大于起测径阶 ) 的林木特征值, 即树木的相对 坐标、 树种、 胸径、 树 块有代表 的 高、 冠幅等 (图 ) , 利用这些特征值计算林分空间结构参数。样 地按照一般林业野外调查要求设置。主分析样地是 性、 能够反映该林区林分的空间结构状况、 面积为 是 !"! 块随机设置的 分析方法 按照森林资源清查和森林经理的观点, 从 个方面入手完整
(针叶树: 红松、 杉松、 鱼鳞云杉; 阔叶树: 水曲柳、 核桃楸) 图 针、 阔叶树种大小比数的比较
是大树。结合树种混交度分析可知, 针叶树种的混交度都很高, 大多针叶树种的大树周围聚集 了相对小的其它树种。核桃楸较占优势, 水曲柳受压较多。 进一步通过临时样地的数据对树种的大 小比数作分析。图 、 表 是各树种的大小比 到
[ ]
, 随机分布的角尺度取值范围是 [
树种 青楷槭 蒙古栎 色木 水曲柳 核桃楸 千金榆 香杨 紫椴 杉松 枫桦 裂叶榆 白扭槭 黄榆 红松 黄波罗 鱼鳞云杉
表 ! 由参照树数据求得各树种 平均混交度 ( !) 及其组成
!
的青楷槭仅以本种为伴) , 在色木、 核
桃楸、 千金榆等树种的单木中 !"
极低。在没有零度混交的树种中, 水曲柳、 紫 椴、 杉松、 裂叶榆、 白扭槭、 香杨等有轻度混交
起, 同种聚集的情况不多, 并且红松在生长上占有一定优势, 这与实地调查情况相符。核桃楸、 水曲柳和黄波罗也多与不同树种邻接, 黄波罗的混交程度最高。在大小配置上, 核桃楸和黄波 罗多处于优势状态, 水曲柳多处于劣势。 !"! 林木个体空间分布格局 用角尺度描述林分中的林木个体分布格局时, 关注林木个体之间的方位关系, 不必象分析 混交度和大小比数那样分树种统计, 只要考虑整个样地的取值情况即可。对于 株最近相邻 木而言, 最优标准角为 度小于
林业科学研究
,( ) :
文章编号:
(
)
吉林蛟河天然红松阔叶林的空间结构分析 !
胡艳波 ,惠刚盈 ,戚继忠 ,安慧君 ,郝广明
( 中国林业科学研究院林业研究所, 北京 ; 北华大学林学院, 吉林 吉林 ) ; 北京林业大学森林资源与环境学院,北京
摘要: 利用混交度、 大小比数和角尺度等三种结构参数, 分析了红松阔叶天然林的空间结构。用平均 混交度和混交度分布分析了林分的树种空间配置情况, 得到该种林分主要以中、 强度混交为主; 用平 均角尺度和角尺度分布分析了林木水平分布格局, 得到该种林分以随机分布为主等结论。同时还应 用大小比数和混交度分析了一些主要树种的空间属性, 发现两种参数的结合可以清楚地描述树种的 空间属性。 关键词: 红松阔叶混交林; 空间结构; 混交度; 大小比数; 角尺度 中图分类号: 文献标识码:
[ ] 生态位, 它在很大程度上决定了林分的稳定性、 发展的可能性和经营空间大小 。分析和重建
林分空间结构是研制新一代林分生长模型的重要基础, 也是制定森林经营规划方案的前提, 因
[ 此近年来对林分空间结构的研究倍受关注, 提出了多种描述林分空间结构的参数 ]
。红松
阔叶林是我国东部小兴安岭和长白山地区森林的顶级植被, 红松阔叶天然林是阔叶红松原始 林过伐几十年后经封育得到恢复的林分, 具有一定的植被代表性。为了更好地保护和经营红 松阔叶天然林, 并促进其向阔叶红松林转变, 必须清楚地了解林分的空间结构现状及发展趋 势。本文利用描述空间结构的混交度、 大小比数和角尺度等参数分析了红松阔叶天然林林分 的空间结构, 为制定和实施森林经营及保护规划提供可释性强的空间结构信息支持。
,,]
强度, 过渡阶段完整, 符合自然情况, 生物学意义明显, 空间结构信息比较完整。另外, 这种结
[ 构单元的可释性和可操作性都比较强, 较适宜于描述林分的空间结构
, 因此本文采用这
种结构单元定义所有结构参数, 并使用树种混交度描述树种空间隔离程度, 使用大小比数描述 林木大小分化程度, 使用角尺度描述林木水平分布格局。 树种混交度 ( ) 被定义为参照树 的 比例, 用公式 ( ) 表示为: 株最近相邻木中与参照树不属同种的个体所占的
张生、 徐炳芳、 孙国文等同志, 在此一并致谢! ! 参加野外调查的还有吉林省蛟河实验林场的陶丽霞、
林
业
科
学
研
究
第
卷
长白山系张广才岭支脉断块中山地貌, 山势浑圆, 东北部山高坡陡, 西南部地势平缓。相对海 拔在 以下, 主要地带土壤为暗棕壤。该区属温带大陆性季风湿润气候, 年平均气温 左右, 年蒸发量 以上。 ) 、 杉松 ) 、 色木 ) 、 ) 、 千金榆 ( ) 、 紫椴 ( ) 、 软枣猕 ) 、 五味子 ( ) ) 、 刺五加 ( ) 等。常见的下木有花楷槭 ( ) 、 楔叶绣线菊窄叶变种 ( ( ) ) 等。 ) 、 鱼鳞云杉 ( ) 、 水曲柳 ( ) 、 蒙古栎 ( ) 、 裂叶榆 ( ( ) 、 胡枝子 ( 该区属于长白山植物区系。试验地主要树种有: 红松 ( ( ( 黄波罗 ( ) 、 东北槭 ( ) 等。层外植物有山葡萄 ( 猴桃 ( ) 、 臭冷杉 ( ) 、 核桃楸 ( 以下。年平均降水量
[ 木个体在水平地面上的分布形式, 或者说是种群的空间分布格局 (描述非规则性) ]
。
空间结构分析以空间结构单元分析为基础。空间结构单元由任意 株单木与其最近几株 相邻木构成, 单元核心的单木被称作参照树。许多研究表明, 在 株参照树与 株相邻木组成 的结构单元中, 株最近相邻木与参照树构成的结构关系有 种, 即零度、 弱度、 中度、 强度、 极
试验地概况
吉林蛟河的红松阔叶林位于蛟河县前进乡境内的吉林林学院实验林场东大坡自然保护区 内, 其地理坐标为 , 。属于吉林省东部褶皱断山地地貌,