经典信号通路之PI3K-AKT-mTOR信号通 路
PI3KAktmTOR信号传导通路在肿瘤的研究进展

三、针对PI3K/Akt/mTOR信号通路的肿瘤治疗
然而,针对PI3K/Akt/mTOR信号通路的抑制剂治疗也面临着一些挑战。一方 面,这些抑制剂往往是非特异性的,可能会影响到其他与肿瘤无关的生理过程, 导致副作用的产生。另一方面,肿瘤细胞经常会对这些抑制剂产生抗性,这可能 需要为靶点的肿瘤治疗
然而,尽管已经有一些初步的临床试验显示出这些抑制剂对某些肿瘤的有效 性,但是大多数抑制剂在临床试验中并未达到预期的效果,并且往往伴随着严重 的副作用。这可能是因为PI3K/Akt/mTOR信号传导通路在人体正常生理过程中也 起着重要的作用,因此干扰这个通路可能会影响到机体的其他功能。
四、未来展望
总结来说,PI3K/Akt/mTOR信号传导通路在肿瘤的发生和发展过程中扮演了 关键的角色。理解这个通路的机制以及寻找以这个通路为靶点的有效治疗策略, 对于改善肿瘤患者的预后和生活质量具有重要的意义。我们期待着在这一领域中 取得更多的突破性进展。
参考内容
PI3K/Akt/mTOR信号通路与肿瘤
二、PI3K/Akt/mTOR信号传导通路在肿瘤中的作用
3、PI3K/Akt/mTOR信号传导通路与肿瘤的血管生成:PI3K/Akt/mTOR信号传 导通路在肿瘤的血管生成中也起着关键作用。肿瘤细胞通过激活PI3K/Akt/mTOR 信号传导通路来刺激血管内皮细胞生长和迁移,从而促进肿瘤的血管生成。这为 肿瘤提供了更多的营养和氧气供应,帮助其快速生长。
二、PI3K/Akt/mTOR信号传导通 路在肿瘤中的作用
二、PI3K/Akt/mTOR信号传导通路在肿瘤中的作用
1、PI3K/Akt/mTOR信号传导通路与肿瘤细胞的生长和增殖:许多肿瘤中存在 PI3K/Akt/mTOR信号传导通路的异常激活。这种激活可以促进肿瘤细胞的生长和 增殖,增加其存活能力。在肝癌、乳腺癌、肺癌等多种肿瘤中,PI3K/Akt/mTOR 信号传导通路的异常激活都被发现并与肿瘤的不良预后有关。
如何选择关键靶点PI3K、AKT和mTOR的抑制剂?针对癌症中的PI3K AKT信号通路

如何选择关键靶点PI3K、AKT和mTOR的抑制剂?针对癌症中的PI3K / AKT信号通路-07-23 订阅号APExBIOPI3K / AKT信号通路的异常激活会导致癌症的发展。
PI3K家族分为四类:I类,II类,III类和IV类。
I类可以进一步分为IA和IB类。
本文主要介绍IA类PI3K。
IA类PI3K是由调节性p85亚基和催化性p110亚基组成的异二聚体。
前者由PIK3R1(p85α,p55α和p50α)和PIK3R2(p85β)基因编码,后者由PIK3CA(p110α),PIK3CB(p110β)和PIK3CD (p110δ)编码。
PI3K的信号传导是在细胞表面受体激活时触发的。
p85与活化的受体酪氨酸激酶(RTK)或相关的衔接蛋白的结合激活催化亚基并使其更接近质膜。
在那里,p110磷酸化磷脂酰肌醇4,5-二磷酸(PIP2,质膜的次要组分)以产生磷脂酰肌醇3,4,5-三磷酸(PIP3),其作为激活各种效应蛋白(包括AKT)的第二信使。
AKT的活化是通过3-磷酸肌醇依赖性蛋白激酶-1(PDK1)和mTOR复合物2(TORC2)的磷酸化完成的。
反过来,AKT的激活导致TORC1的激活,AKT和TORC1均调节促进细胞的存活、生长和增殖等途径。
另外,p110α和p110δ可以被Ras(GTP酶转换蛋白)进一步激活,Ras不与p110β结合。
Rac和Cdc42蛋白有助于激活p110β以响应G蛋白偶联受体(GPCR)激活。
TORC1信号传导还充当负反馈调节机制以抑制来自RTK的信号传导。
最后,肿瘤抑制因子PTEN使PIP3去磷酸化并返回PIP2(图1)。
图1 PI3K(IA类) / AKT信号通路的激活(此图列出了多种抑制剂,不知如何选择抑制剂的可作为参考)图2 PI3K(IA 类) / AKT信号通路的药理学抑制。
PI3K通路可以在多个节点上被药理学抑制,包括pan-PI3K和同种型特异性抑制剂,只靶向TORC1或同时靶向TORC1和TORC2的mTOR抑制剂以及AKT抑制剂,这些抑制剂可以直接抑制PI3K活性。
PI3K_Akt_mTOR信号转导通路与卵巢癌研究进展_朱颖军

近年研究发现, 生长因子的细胞内信号转导通 路同肿瘤发生、发展及转移密切相关。 其中磷脂酰肌 醇 3 激酶 (phosphatidylinositol 3-kinase,PI3K)/蛋白 激 酶 B (serine/threonine kinase,Akt;protein kinase B,PKB)/哺 乳 动 物 雷 帕 霉 素 靶 体 蛋 白 (mammalian target of rapamycin,mTOR) 信号转导通路的研究取 得 了 一 定 成 果 ,PI3K/Akt/mTOR 信 号 转 导 通 路 在 恶 性肿瘤细胞增殖、血管新生和转移以及肿瘤对放、化 疗的拮抗中起着重要作用, 本文就该通路与卵巢癌 的关系综述如下。
信号通路3—PI3K-AKT-mTOR

信号通路3 —PI3K/AKT/mTORAPExBIO一、PI3K/Akt/mTORPI3K/AKT/mTOR是调节细胞周期的重要细胞内信号通路。
PI3K/AKT/mTOR信号通路与细胞的休眠、增殖、癌变和寿命直接相关。
PI3K激活后磷酸化并激活AKT,将其定位在质膜中。
信号通过AKT传递到下游不同的靶点,如激活CREB,抑制p27,将FOXO定位于细胞质中,激活PtdIns-3ps,及激活mTOR(影响p70或4EBP1的转录)。
该通路的激活因子包括EGF、shh、IGF-1、胰岛素和CaM。
该信号通路的拮抗因子,包括PTEN、GSK3B、和HB9。
在多种癌症中,PI3K/AKT/mTOR通路是过度活化的,因此减少凋亡并促进增殖。
然而,该通路在成人干细胞尤其是神经干细胞的分化过程中促进细胞生长和增殖。
1. PI3KPhosphatidylinositide 3-kinases,是一种胞内磷脂酰肌醇激酶。
由调节亚基p85和催化亚基p110构成。
与v.sre和v.ras等癌基因的产物相关。
PI3K本身具有丝氨酸/苏氨酸(Ser/Thr)激酶的活性,也具有磷脂酰肌醇激酶的活性。
2. Akt又称PKB(protein kinase B)。
是一种丝氨酸/苏氨酸特异性蛋白激酶,在多种细胞生长过程中发挥关键作用,如葡萄糖代谢、凋亡、细胞增殖、转录和细胞迁移。
Akt的Ser473可以被PDK1磷酸化。
PKB与PKA和PKC均有很高的同源性,该激酶被证明是反转录病毒安基因v-akt 的编码产物,故又称Akt。
3. mTORMammalian target of rapamycin。
mTOR与其它蛋白质结合,形成两种不同蛋白质复合物,mTOR复合物1(mTORC1,)和mTOR复合物2(mTORC2),它们调节不同的细胞过程。
mTORC1由mTOR、mTOR调节相关蛋白Raptor、MLST8和非核心组分PRAS40、DEPTOR 组成。
PI3K—AKT—mTOR信号通路的研究进展

PI3K—AKT—mTOR信号通路的研究进展PI3K-AKT-mTOR信号转导通路是哺乳动物肿瘤免疫中重要的信号通路,在多种恶性肿瘤的演变过程中发挥了极其重要的作用。
近几年来,随着肿瘤分子生物学的发展,恶性肿瘤的靶向治疗成为研究热点,通过研究探讨PI3K-AKT-mTOR信号通路在肿瘤发生、发展过程中的信号转导机制,联合多种抑制剂或者寻找作用于多种信号通路、多靶点的新药,对于肿瘤的靶向治疗有重要意义。
标签:PI3K-AKT-mTOR;信号转导;肿瘤;抑制剂恶性肿瘤严重危害人类健康,随着社会、经济的发展以及人口老龄化的加剧,我国大多数恶性肿瘤发病率、死亡率呈明显上升趋势。
与此同时,随着人们对恶性肿瘤的研究不断深入,越来越多肿瘤信号通路被发现,其中PI3K-AKT-mTOR 信号转导通路是哺乳动物肿瘤免疫中的重要信号通路,此信号通路对于调节细胞的生长、增殖、自噬以及凋亡有着重要的作用。
1 PI3K-AKT-mTOR 信号通路的组成在各种生物体中,细胞之间相互识别及相互作用,都是通过细胞信号的传导来实现,细胞信号转导指细胞通过细胞膜或者胞内相应受体感受信息分子刺激,通过细胞内信号转导系统进行转换,从而引发一系列生物化学反应及蛋白相互作用,直到细胞生理反应所需基因表达开始、各种生物学效应形成。
1.1 磷脂酰肌醇-3激酶(PI3K)PI3K存在于细胞质中,具有蛋白激酶及磷脂激酶的双重活性。
PI3K包括Ⅰ型、Ⅱ型、Ⅲ型,Ⅰ型的底物主要为磷脂酰肌醇(PI)、3-磷酸磷脂酰肌醇(PIP)及3,4-二磷酸磷脂酰肌醇(PIP2);Ⅱ型的底物主要为PI及PIP,Ⅲ型的底物主要为PI,但是只有Ⅰ型PI3K与肿瘤形成有着密切关联[1-2]。
Ⅰ型PI3K包括IA和IB亚型,它们从酪氨酸激酶连接受体、G蛋白连接受体进行信号传递,IA型PI3K由调节亚基(P58)和催化亚基(P110)组成,其中调节亚基(P58)包含SH2、SH3两个重要结构域,在正常情况下P58与P110结合导致PI3K失活。
小分子抑制剂、激动剂、拮抗剂--PI3KAktmTOR信号通路
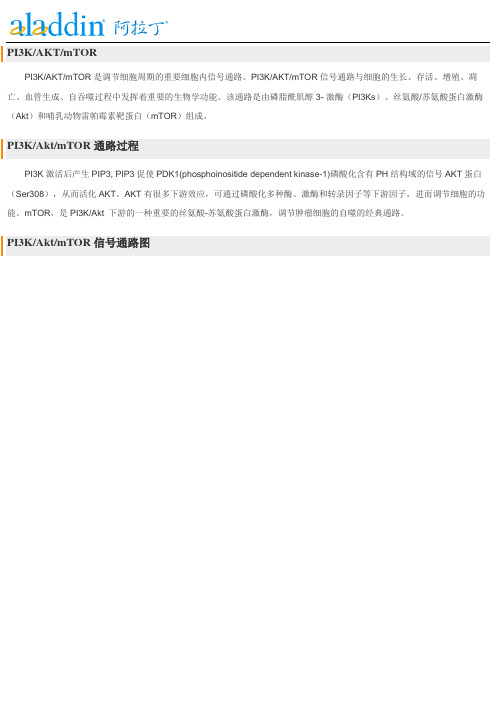
PI3K/AKT/mTORPI3K/AKT/mTOR是调节细胞周期的重要细胞内信号通路。
PI3K/AKT/mTOR信号通路与细胞的生长、存活、增殖、凋亡、血管生成、自吞噬过程中发挥着重要的生物学功能。
该通路是由磷脂酰肌醇3- 激酶(PI3Ks)、丝氨酸/苏氨酸蛋白激酶(Akt)和哺乳动物雷帕霉素靶蛋白(mTOR)组成。
PI3K/Akt/mTOR通路过程PI3K激活后产生PIP3, PIP3促使PDK1(phosphoinositide dependent kinase-1)磷酸化含有PH结构域的信号AKT蛋白(Ser308),从而活化AKT。
AKT有很多下游效应,可通过磷酸化多种酶、激酶和转录因子等下游因子,进而调节细胞的功能。
mTOR,是PI3K/Akt 下游的一种重要的丝氨酸-苏氨酸蛋白激酶,调节肿瘤细胞的自噬的经典通路。
PI3K/Akt/mTOR信号通路图按靶点分类:*PI3KPI3K,是一种胞内磷脂酰肌醇激酶,也具有丝氨酸/苏氨酸(Ser/Thr)激酶的活性。
能够通过PI3K诱发PIP3生成的激活因子,则能够激活Akt 信号途径,包括受体酪氨酸激酶、整合素、B 细胞和T 细胞受体、细胞因子受体、G 蛋白偶联受体等等。
*Akt又称PKB或Rac,是一种丝氨酸/苏氨酸特异性蛋白激酶B,在细胞存活和凋亡中起重要作用,如葡萄糖代谢、凋亡、细胞增殖、转录和细胞迁移。
Akt的Thr308可以被PDK1磷酸化,而被部分激活。
或者473位点上的丝氨酸被mTORC2磷酸化,激发Akt的完全酶活性。
*mTORmTOR是细胞生长和增殖的重要调节因子。
mTOR与其它蛋白质结合,形成两种不同蛋白质复合物,mTORC1和mTORC2,参与调节不同的细胞过程。
*GSK-3。
PI3K-AKT-mTOR信号通路与前列腺癌

PI3K-AKT-mTOR信号通路与前列腺癌PI3K / Akt / mTOR信号通路具有多种功能,包括调节细胞存活,分化和干细胞样特性,生长,增殖,代谢,迁移和血管生成。
成员:PI3K、AKT、mTOR、PTENPI3K,是磷脂酰肌醇-3-激酶的缩写,可分为3类,Ⅰ、Ⅱ、Ⅲ类。
其中研究最广泛的为Ⅰ类PI3K,Ⅰ类PI3K由酪氨酸激酶受体(RTK)、G蛋白偶联受体和一些致癌基因(如大鼠肉瘤癌基因RAS)激活。
Ⅰ类又可分ⅠA和ⅠB类,其中ⅠA类PI3K与癌症最相关。
p85α ( 由PIK3R1编码)β (由PIK3R2编码)γ ( 由PIK3R1编码)p110α( 由PIK3CA编码)p110β(由PIK3CB编码)p110δ(由PIK3CD编码)AKT(蛋白激酶B,PKB,又称为Akt)是PI3K重要的下游分子,被活化后,通过多种方式来影响其下游的相关分子,使细胞存活,是关键的抗凋亡调控分子mTOR(哺乳动物雷帕霉素靶蛋白)是一种丝氨酸-苏氨酸蛋白激酶,细胞内存在mTORC1和mTORC2两种不同的复合体。
其属于PI3K/Akt 下游的重要成分。
PTEN(磷酸酶和张力蛋白同源物))是一种肿瘤抑制蛋白。
可将PIP3去磷酸化生成PIP2。
当PTEN 下调时,可引起PI3K / Akt / mTOR 途径的激活。
PI3K-AKT-mTOR信号通路PI3K的激活导致4,5-二磷酸磷脂酰肌醇[PI(4,5)P2]磷酸化为3,4,5-三磷酸磷脂酰肌醇[PI(3,4,5)P3],随后将Akt募集到质膜上并激活。
(当配体与膜受体结合后,受体激活p85并招募p110,进而催化膜内表面的PIP2生成PI3P。
PI3P作为第二信使,进一步激活AKT和PDK1。
)Akt活化通过两个残基的磷酸化介导:T308通过磷酸肌醇依赖性蛋白激酶-1(PKD1)磷酸化;S473通过mTOR复合物2(TORC2)磷酸化。
Akt的完全激活是必需通过上述两者磷酸化。
胶质瘤PI3K/AKT/mTOR 信号通路靶向治疗研究进展

胶质瘤PI3K/AKT/mTOR 信号通路靶向治疗研究进展胶质瘤是一种发生于神经外胚层的常见原发性颅内肿瘤。
随着分子生物学的发展,相关信号传导通路特异分子靶向治疗成为胶质瘤治疗的热点并取得了一定的成果。
近年来,大量研究揭示PI3K/AKT/mTOR信号通路在胶质瘤发生及发展中发挥了重要的作用,因而针对该通路的靶向治疗表现出较好的临床应用前景。
本文着重针对PI3K/AKT/mTOR信号通路在胶质瘤中的作用及其相关抑制剂药物的靶向治疗效果进行综述。
标签:胶质瘤;PI3K/AKT/mTOR;靶向治疗;抑制剂胶质瘤是一种临床上常见的颅内肿瘤,可占中枢神经系统原发性恶性肿瘤的80%。
其中,中间变性胶质瘤以及胶质母细胞瘤往往伴随着较低的治愈率及较高的死亡率[1]。
临床上,胶质瘤的治疗主要是以外科手术结合放疗、化疗等措施为主,然而该类方案的治疗效果仍不理想。
随着分子生物学的发展,针对影响胶质瘤增殖、转移及迁移相关信号傳导通路的靶向治疗成为一种新的治疗模式并取得较好的疗效[2]。
其中,由于其促肿瘤作用,PI3K/AKT/mTOR信号通路分子靶向治疗备受关注。
接下来,本文着重对PI3K/AKT/mTOR信号通路在胶质瘤发生、发展中的作用及相关分子靶向治疗药物进行综述。
1 PI3K/AKT/mTOR信号通路与胶质瘤PI3K/AKT/mTOR信号通路主要由PI3K、AKT及mTOR三个关键因子组成。
该通路的主要传导机制如下:当PI3K受到受体酪氨酸激酶、Ras、整合素以及各种生长因子等多种因子作用而激活时,可通过磷酸化肌醇诱导3,4,5-三磷酸磷脂酰肌醇及3,4-二磷酸磷脂酰肌醇的生成[3];接下来,激活的Akt可通过3-磷脂酰肌醇依赖性蛋白激酶1及3-磷脂酰肌醇依赖性蛋白激酶2磷酸化,进而将信号传递给mTOR[4];而两种复合体形态的mTOR(雷帕霉素敏感mTORC1,对雷帕霉素不敏感mTORC2)可进而对下游相关mRNA的转录进行调节,最终影响细胞增殖、凋亡、分化、迁移等多种生理过程[5]。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
经典信号通路之PI3K-AKT-mTOR信号通路PI3K是一种胞内磷脂酰肌醇激酶,与v.src和v.ras等癌基因的产物相关,且PI3K本身具有丝氨酸/苏氨酸(Ser/Thr)激酶的活性,也具有磷脂酰肌醇激酶的活性。
由调节亚基p85和催化亚基p110构成。
磷脂酰肌醇3-激酶(PI3Ks)蛋白家族参与细胞增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节。
PI3K活性的增加常与多种癌症相关。
PI3K磷 酸化磷脂酰肌醇PI(一种膜磷脂)肌醇环的第3位碳原子。
PI在细胞膜组分中所占比例较小,比磷脂酰胆碱、磷脂酰乙醇胺和磷脂酰丝氨酸含量少。
但在脑细胞膜中,含量较为丰富,达磷脂总量的10%。
PI的肌醇环上有5个可被磷酸化的位点,多种激酶可磷酸化PI肌醇环上的4th和5th位点,因而通常在这两位点之一或两位点发生磷酸化修饰,尤其发生在质膜内侧。
通常,PI-4,5-二磷酸(PIP2)在磷脂酶C的作用下,产生二酰甘油(DAG)和肌醇-1,4,5-三磷酸。
PI3K转移一个磷酸基团至位点3,形成的产物对细胞的功能具有重要的影响。
譬如,单磷酸化的PI-3-磷酸,能刺激细胞迁移(cell trafficking),而未磷酸化的则不能。
PI-3,4-二磷酸则可促进细胞的增殖(生长)和增强对凋亡的抗性,而其前体分子PI-4-磷酸则不 然。
PIP2转换为PI-3,4,5-三磷酸,可调节细胞的黏附、生长和存活。
PI3K的活化PI3K可分为3类,其结构与功能各异。
其中研究最广泛的为I类PI3K, 此类PI3K为异源二聚体,由一个调节亚基和一个催化亚基组成。
调节亚基含有SH2和SH3结构域,与含有相应结合位点的靶蛋白相作用。
该亚基通常称为p85, 参考于第一个被发现的亚型(isotype),然而目前已知的6种调节亚基,大小50至110kDa不等。
催化亚基有4种,即p110α,β,δ,γ,而δ仅限于白细胞,其余则广泛分布于各种细胞中。
PI3K的活化很大程度上参与到靠近其质膜内侧的底物。
多种生长因子和信号传导复合 物,包括成纤维细胞生长因子(FGF)、血管内皮生长因子(VEGF)、人生长因子(HGF)、血管位蛋白I(Ang1)和胰岛素都能启始PI3K 的启动 过程。
这些因子启动受体酪氨酸激酶(RTK),从而引起自磷酸化。
受体上磷酸化的残基为异源二聚化的PI3Kp85亚基提供了一个停泊位点 (docking site)。
然而在某些情况下,受体磷酸化则会介导募集一个接头蛋白(adaptor protein)。
比如,当胰岛素启动其受体后,则必须募集一个胰岛素受体底物蛋白(IRS),来促进PI3K的结合。
相似的,当整连蛋白 integrin(非RTK)被启动后,粘着斑激酶(FAK) 则作为接头蛋白,将PI3K通过其p85停泊。
但在以上各情形下,p85亚基的SH2和SH3结构域均在一个磷酸化位点与接头蛋白结合。
PI3K募集到活化的受体后,起始多种PI中间体的磷酸化。
与癌肿尤其相关的PI3K转化PIP2为PIP3。
PIP3作为锚定物(anchor)许多蛋白含有一个Pleckstrin Homology(PH)结构域,因而可使其与PI-3,4-P2或PI-3,4,5-P3相结合。
这种相互作用可以控制蛋白与膜结合的时间与定位,通过这种方式来调节蛋白的活性。
蛋白与脂质间的这种相互作用亦可能引起蛋白构像的变化而改变蛋白的功能。
PI3K启动的结果是在质膜上产生第二信使PIP3, PIP3与细胞内含有PH结构域的信号蛋白AKT和PDK1(phosphoinositide dependent kinase-1)结合, 促使PDK1磷酸化AKT蛋白的Ser308导致AKT活化。
其它PDK1的底物还包括PKC(蛋白激酶C)、S6K(p70S6)和 SGK(serum/glucocorticoid regulated kinases)。
AKT, 亦称为蛋白激酶B(PKB),是PI3K下游主要的效应物。
AKT可分为3种亚型(AKT1、AKT2、AKT3或PKBα, PKBβ,PKBγ),3种亚型的功能各异,但也有重迭。
该家族主要有三个成员:AKT1,AKT2和AKT3。
其中,Akt1通过抑制细胞凋亡过程参与了细胞生存途径,Akt1酶也能诱导蛋白质的合成途径,因此是一个重要的信号蛋白介导组织的生长。
因为它可以阻止细胞凋亡,从而促进细胞的存活,AKT1参与了在许多类型的癌症发生。
AKT2是胰岛素信号转导通路中的一个重要信号分子,而AKT3则是主要表达在脑部。
它的启动机制是:PI3K可以被g蛋白偶联受体或者受体酪氨酸激酶启动,随后启动的PI3K可以把PIP2磷酸化从而形成PIP3。
PIP3可以募集AKT将其正确定位在膜上,然后AKT可以被PDPK1和 mTORC2磷酸化而被启动,并经一步参与接下来的各种功能。
AKT的作用活化的AKT通过磷酸化多种酶、激酶和转录因子等下游因子,进而调节细胞的功能。
譬如,AKT刺激葡萄糖的代谢:AKT启动AS160(AKT底 物,160kDa),进而促进GLUT4转座和肌细胞对葡萄糖的吸收。
AKT也磷酸化GSK3β而抑制其活性,从而促进葡萄糖的代谢和调节细胞的周期。
AKT 磷酸化TSC1/2(tuberous sclerosis complex),可阻止其对小G蛋白Rheb(Ras homology enriched in brain)的负调控,进而使得Rheb富集以及对纳巴霉素(rapamycin)敏感的mTOR复合体(mTORC1)的活化。
这些作用可启动蛋白的翻 译,增强细胞的生长。
AKT通过下游多种途径对靶蛋白进行磷酸化而发挥抗凋亡作用。
ATK启动IkB激酶(IKKα),导致NF-κB的抑制剂 IκB的降解, 从而使NF-κB从细胞质中释放出来进行核转位, 启动其靶基因而促进细胞的存活。
AKT磷酸化Bcl-2家族成员BAD,使其与14-3-3结合而阻止其与Bcl-XL结合起始凋亡。
此外,AKT能抑制 蛋白水解酶caspase-9的活性而阻止凋亡级联反应的启动。
肿瘤抑制因子p53为一转录因子,调控凋亡、DNA修复和细胞周期的停滞。
Akt能通过磷 酸化P53结合蛋白MDM2影响P53的活性, 磷酸化的MDM2转位到细胞核与P53结合, 通过增加P53蛋白的降解而影响细胞存活。
Forkhead转录因子FOXO1( 或FKHR)调节涉及多种细胞功能基因的表达,包括凋亡、DNA修复和 细胞周期的停滞和葡萄糖代谢等,AKT 磷酸化FOXO1,抑制其核转位而阻止其转录启动作用。
PTEN: 一个关键磷酸酶PTEN (phosphatase and tensin homology deleted on chromosome 10),在广泛的人类肿瘤中发生基因突变或缺失。
PTEN是一个PIP3-磷酸酶,与PI3K的功能相反,它可以通过去磷酸化将PIP3转变为PI- 4,5-P2。
PTEN可减少AKT的活化而阻止所有由AKT调控的下游信号传导事件。
通过产生PIP2, PTEN的功能远不止仅仅阻止AKT的作用。
PIP2作为磷脂酶Cβ(PLCβ)的底物,产生DAG和IP3作为第二信使,升高胞内钙离子的水平并启动蛋 白激酶C(PKC)。
与膜结合的PIP2也能介导(增加)多种离子通道的活性,包括钙离子、钾离子和钠离子通道。
PIP也参与到膜泡的形成以及细胞骨架与 膜的相互作用过程中。
PIP2还影响多种参与脂代谢的酶活性,包括磷脂酶D和神经酰胺-1-磷酸。
通过从PIP产生PIP2, PTEN广泛的影响着细胞的多种功能途径。
PTEN在减弱PIP3信号传导而增加PIP2水平的过程中,发挥着重要的作用。
PDK的活化与抑制当接受来自酪氨酸激酶和G蛋白偶联受体的信号后,PI3K的p85调节亚基即被募集到临近质膜的部位,p110亚基通过与p85亚基结合把底物Ptd Ins(4,5)P2(PIP2)转化为Ptd Ins(3,4,5)P3(PIP3)。
PI(3,4,5)P3可以和蛋白激酶B(PKB,Akt)的N端PH结构域结合。
使Akt从细胞质转移到细胞膜上.并在3一磷酸肌醇依赖性蛋白激酶1(PDKI)的辅助下,通过使Akt蛋白上的苏氨酸磷酸化位点(Thr308)和丝氨酸磷酸化位点(Ser473)磷酸化而使其启动。
启动后的Akt通过直接和间接两种途径启动其底物雷帕霉素靶体蛋白(mTOR):直接磷酸化mTOR,或者通过失活结节性硬化复合物2(TSC2)从而维持Rheb的GTP结合态,然后增强roTOR的启动。
肿瘤抑制基因PrEN f Phosphatase and tensin homolog deleted o chromosome 10,第10号染色体磷酸酶和张力蛋白同源丢失性基因)编码的产物可以使PIP3在D3位去磷酸化生成PIP2,从而实现P13K/Akt信号通路的负性调节,抑制细胞增殖和促进细胞凋亡。
PI3K-AKT-mTOR is one of the three major signalling pathways that have been identified as important in cancer. mTOR is a key kinase downstream of PI3K/AKT, which regulates tumor cell proliferation, growth, survival and angiogenesis. Cancer cells escape normal biochemical systems regulating the balance between apoptosis (suicide) and survival. PI3K-AKT-mTOR generally acts to promote survival through inhibition of proapoptotic factors and activation of anti-apoptotic factors. Through phosphorylation, PI3K-AKT-mTOR inhibits the activity of proapoptotic members while activating anti-apoptotic members. To negatively regulate PI3K, cells contain PTEN phosphatase. A reduction in PTEN expression indirectly stimulates PI3K-AKT-mTOR activity thereby contributing to oncogenesis in human. Recent data suggests that the PI3K-AKT-mTOR signaling pathway plays an important role in cancer stem cell self-renewal and resistance to chemotherapy or radiotherapy, which is believed to be the root of treatment failure and cancer recurrence, as well as metastasis.癌症中的PI3K-mTOR信号途径Figure 1 | Minding your Ps: the PtdIns(4,5)P2–PtdIns(3,4,5)P3 cycle. Phosphatidylinositol phosphates are composed of a membrane-associated phosphatidic acid group and a glycerol moiety that is linked to a cytosolic phosphorylated inositol head group. Phosphatidylinositol 3-kinase (PI3K) can phosphorylate PtdIns(4,5)P2 (PIP2) at the D3 position to form the second messenger PtdIns(3,4,5)P3 (PIP3). Phosphorylation at the D3 position is necessary for binding to the pleckstrin-homology domain of AKT (not shown). Dephosphorylation of PIP3 to regenerate PIP2 is accomplished by the 3-phosphatase PTEN. Additionally, PIP3 can be dephosphorylated at the D5 position by SHIP1 or SHIP2 to generate PtdIns(3,4)P2, another potential second messenger.Figure 2 | PI3K的启动机制. Autophosphorylation of ligand-activated receptor tyrosine kinases (RTKs) causes recruitment of inactive heterodimeric class IA phosphatidylinositol 3-kinases (PI3Ks) through the interaction of phosphotyrosine residues on the receptor and SRC-homology 2 (SH2) domains on the PI3K p85 regulatory subunit, or the adaptor proteins IRS1 and IRS2. IRS1 and IRS2 are phosphorylated by the activated receptor, generating docking sites for the SH2 domains of p85 and inducing proper assembly of the signalling complex. These SH2–phosphotyrosine interactions bring PI3K in close proximity to its substrate at the plasma membrane and relieve the inhibitory action of p85 on the p110 catalytic subunit, which is then free to convert PtdIns(4,5)P2 (PIP2) into PtdIns(3,4,5)P3 (PIP3). Alternatively, binding of PI3K to activated RAS can also stabilize its membrane localization and activate the catalytic domain. This occurs by recruitment of the adaptor proteins SHC, GRB2 and GAB2 to activated RTKs. C2, C2 domain; CD, catalytic domain; p85 BD, p85-binding domain;RBD, RAS-binding domain.Figure 3 |AKT信号的调控. Activation of AKT is initiated by membrane translocation,which occurs after cell stimulation and PtdIns(3,4,5)P3 (PIP3) production. Localization of AKT to the plasma membrane is accomplished by an interaction between its pleckstrin-homology (PH) domain and PIP3. At the membrane, association with carboxy-terminal modulator protein (CTMP) prevents AKT from becoming phosphorylated and fully active. Phosphorylation of CTMP by an asyet unidentified kinase releases CTMP from AKT and allows AKT to be phosphorylated by PDK1 and PDK2 at Thr308 and Ser473, respectively. Phosphorylation at these two sites causes ful activation of AKT. C2, C2 domain; CD, catalytic domain; p85 BD, p85-binding domain.Figure 3 |PI3K信号通路 Activation of class IA phosphatidylinositol 3-kinases (PI3Ks) occurs through stimulation of receptor tyrosine kinases (RTKs) and the concomitant assembly of receptor–PI3K complexes. These complexes localize at the membrane where the p110 subunit of PI3K catalyses the conversion of PtdIns(4,5)P2 (PIP2) to PtdIns(3,4,5)P3 (PIP3). PIP3 serves as a second messenger that helps to activate AKT. Through phosphorylation, activated AKT mediates the activation and inhibition of several targets, resulting in cellular growth, survival and proliferation through various mechanisms. Additionally, PI3K has been shown to regulate the activity of other cellular targets, such as the serum and glucocorticoid-inducible kinase (SGK), the small GTP-binding proteins RAC1 and CDC42, and protein kinase C(PKC), in an AKT-independent manner through poorly characterized mechanisms. The activity of these targets leads to survival, cytoskeletal rearrangement and transformation. GSK3。