基于抑制消减杂交识别白化刺参和青刺参差异表达基因
不同色型仿刺参Apostichopus japonicus(Selenka)环境适应性与营养需求的比较研究.pptx

以vc磷酸酯l-ascorbyl-2-polyphosphate(lapp)为添加剂配制6 种不同维生素c含量的饲料(0、50、100、200、400和800mg维生 素c等价物kg-1饲料)分别饲养青色、白色和紫色三种色型刺参 60天,并测定其生长、能量收支和非特异性免疫酶活性。结果显 示,饲料维生素c含量对刺参的生长有显著影响(p<0.05)。
过氧化氢酶和谷草转氨酶活性与特定生长率呈显著正相关,表明 刺参的快速生长需要免疫系统和蛋白代谢能力的支撑。饲料蛋 白含量为1%时,紫刺参特定生长率、食物转化效率、粗蛋白表观 消化率、生长能以及过氧化氢酶活性显著高于青刺参,表明紫刺 参对于低蛋白饲料有更好的适应性。
白刺参特定生长率、食物转化效率、生长能、超氧化物歧化酶 和过氧化氢酶活性显著低于青刺参和紫刺参,而排泄能和体腔液 游离氨含量显著更高,表明白刺参可能在蛋白代谢方面有所不同。 4、不同饲料维生素c含量对不同色型刺参生长、能量收支及关 键免疫指标的影维生素c在刺参的生长和免疫过程中有着重要的 作用。
结果显示,温度对刺参的生长、能量收支及关键生理生化指标影 响显著(P<0.05)。三种色型刺参的特定生长率、食物转化效 率以及生长能比例均在18?C时达到峰值,而摄食率和摄入能量在 22?C最高。
耗氧率和溶菌酶活性随着温度的升高而升高。三种色型刺参的 超氧化物歧化酶、过氧化氢酶以及胰蛋白酶活性均在18?C时达 到最大值。
青刺参和紫刺参在饲料维生素c含量为200mgkg-1时特定生长率、 摄食率以及食物转化效率最高,而白刺参在100mgkg-1时达到峰 值,表明白刺参的维生素c需求低于青刺参和紫刺参。这可能是 因为白刺参较低的生长率和代谢率——其特定生长率、摄食率、 食物转化效率以及各项免疫指标均低于青刺参和紫刺参。
【国家自然科学基金】_抑制性消减杂交技术(ssh)_基金支持热词逐年推荐_【万方软件创新助手】_20140730
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40
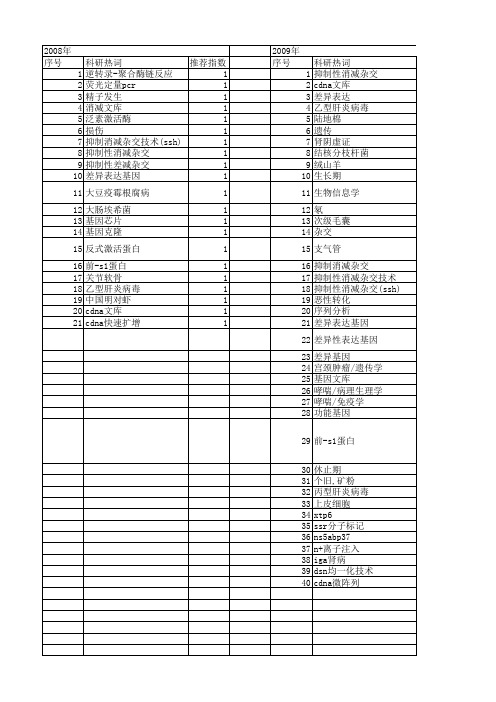
科研热词 抑制性消减杂交 cdna 差异表达 乙型肝炎病毒 陆地棉 遗传 肾阴虚证 结核分枝杆菌 绒山羊 生长期 生物信息学 氡 次级毛囊 杂交 支气管 抑制消减杂交 抑制性消减杂交技术 抑制性消减杂交(ssh) 恶性转化 序列分析 差异表达基因 差异性表因 前-s1蛋白 休止期 个旧,矿粉 丙型肝炎病毒 上皮细胞 xtp6 ssr分子标记 ns5abp37 n+离子注入 iga肾病 dsn均一化技术 cdna微阵列
2014年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
海参DNA条形码的构建及应用

中国水产科学 2011年7月, 18(4): 782-789 Journal of Fishery Sciences of China研究论文收稿日期: 2010-11-05; 修订日期: 2011-01-04.基金项目: 海洋公益性行业科研专项(201105029); 国际科技合作项目(2010DFA31330);“泰山学者”建设工程专项经费资助项目(JS200510013); 浙江省重大科技专项(2008C2009).作者简介: 律迎春(1988-), 男, 硕士研究生, 研究方向为水产品质量监督与控制. E-mail: oasis0212@ 通信作者: 唐庆娟(1971-), 讲师. E-mail: tangqingjuan@DOI: 10.3724/SP.J.1118.2011.00782海参DNA 条形码的构建及应用律迎春, 左涛, 唐庆娟, 段高飞, 王春霞, 李晋, 徐洁, 薛长湖中国海洋大学 食品科学与工程学院, 山东 青岛 266003摘要: 为研究线粒体细胞色素c 氧化酶亚基(Mitochondr Ⅰial cytochrome c oxidase subunit , CO )ⅠⅠ基因作为DNA 条形码鉴定海参品种的可行性, 本实验采集了7种海参(Holothuroidea)43个个体并获得其线粒体CO Ⅰ基因序列, 利用DNAstar 、DNAMAN 和MEGA 4.1软件分析计算了7种海参的碱基组成以及不同海参之间的种间遗传距离和种内遗传距离, 利用邻接法和最大简约法分别构建了分子系统树。
结果表明, 海参的种间遗传距离显著大于种内遗传距离, 不同海参在系统树中分别形成各自独立的分支, 表明以CO Ⅰ作为海参DNA 条形码进行品种鉴定具有一定的可行性。
在建立海参DNA 条形码的基础上, 设计了针对仿刺参(Apostichopus japonicas )的特异性探针。
对4种海参(仿刺参Apostichopus japonicas 、冰岛参Cucumaria frondosa 、加州拟刺参Parastichopus californicus 及梅花参Thelenota ananas )进行斑点杂交实验, 结果显示, 该探针具有较好的特异性和较高的灵敏度, 能够实现对仿刺参的鉴定。
211251883_菌酶联用处理对刺参体壁降解效果及风味特性的影响

邹媛婷,张健,刘芳,等. 菌酶联用处理对刺参体壁降解效果及风味特性的影响[J]. 食品工业科技,2023,44(11):118−126. doi:10.13386/j.issn1002-0306.2022080279ZOU Yuanting, ZHANG Jian, LIU Fang, et al. Effects of Combined Treatments of Bacteria and Enzyme on the Degradation and Flavor Components of Body Wall of Sea Cucumber[J]. Science and Technology of Food Industry, 2023, 44(11): 118−126. (in Chinese with English abstract). doi: 10.13386/j.issn1002-0306.2022080279· 研究与探讨 ·菌酶联用处理对刺参体壁降解效果及风味特性的影响邹媛婷1,2,张 健2, *,刘 芳2, *,井月欣2,王共明2,矫春娜2,赵云苹2(1.上海海洋大学食品学院,上海 201306;2.山东省海洋资源与环境研究院,山东烟台 264006)摘 要:目的:为探究不同菌酶联用处理方式对刺参体壁降解效果及产物风味特性的影响。
方法:采用不同处理方式:单菌(J )、单酶(M )、先菌后酶(JM )、先酶后菌(MJ )、菌酶同用(T )(菌为解淀粉芽孢杆菌,酶为复合蛋白酶)对刺参体壁进行降解,测定降解液中氨基酸组成、多肽相对分子质量分布等理化指标,同时利用气相色谱-离子迁移谱(GC-IMS )和电子舌分别测定不同处理方式下刺参体壁降解产物的挥发性和非挥发性风味成分。
结果:刺参体壁经菌酶联合(JM 、MJ 、T )处理后,与单独作用相比,相对分子质量>1000 Da 的肽含量减少了60.29%,<500 Da 的肽含量增加了33.36%,联合处理下游离氨基酸总量与风味核苷酸总量的平均值较菌酶单独作用时分别增加了1.3倍和4.1倍,降解液中挥发性化合物由未处理时的43种增加到联合处理后的67种。
我国北方甘蓝型冬油菜抗寒性评价及冷冻胁迫下转录组分析

我国北方甘蓝型冬油菜抗寒性评价及冷冻胁迫下转录组分析我国北方甘蓝型冬油菜抗寒性评价及冷冻胁迫下转录组分析导语:冬油菜是我国北方广泛种植的经济作物,但其种植过程中抗寒性是一个关键的限制因素。
本研究通过对我国北方甘蓝型冬油菜的抗寒性评价以及冷冻胁迫下的转录组分析,旨在揭示冬油菜抗寒性的调节机制,为提高冬油菜的抗寒性提供科学依据。
一、引言冬油菜(Brassica napus L.)是我国北方重要的油料作物之一,其具有丰富的油脂含量和营养价值,广泛应用于植物油和饲料加工等领域。
然而,由于北方冬季寒冷的气候条件,冬油菜在种植过程中容易受到低温的影响而减产甚至死亡。
因此,提高冬油菜的抗寒性是农业生产中迫切需要解决的问题之一。
二、抗寒性评价方法本研究选择了在北方地区广泛种植的甘蓝型冬油菜品种进行抗寒性评价。
通过不同温度处理,包括常温(25℃)、低温(4℃)和冷冻处理(-5℃),观察和记录不同处理下冬油菜的生长状况和生物学指标,如株高、叶面积、叶片颜色、生长速率等。
根据观察结果,可以评价不同品种的抗寒性差异,并筛选出抗寒性较好的品种作为研究对象。
三、冷冻胁迫下转录组分析为了进一步研究冬油菜在冷冻胁迫下的抗寒机制,本研究采用转录组分析的方法,深入探究胁迫过程中基因的表达变化。
首先,我们提取了冷冻胁迫前后的冬油菜叶片样品总RNA,利用高通量测序技术对其进行测序,并对测序结果进行数据分析和生物信息学处理。
其次,基于差异表达基因筛选和功能富集分析,我们研究了在冷冻胁迫下表达变化的关键基因,并对其进行了进一步的验证实验。
根据转录组分析的结果,我们发现冷冻胁迫下冬油菜叶片基因表达发生显著变化。
一方面,一些与抗寒性相关的基因在冷冻胁迫条件下被激活,从而增强了植物对低温的适应能力;另一方面,一些与生长发育相关的基因则受到了抑制,导致植物的生长速率减缓。
此外,我们还发现了一些新的候选基因,这些基因可能具有与冷冻胁迫相关的特殊功能,值得进一步的研究。
刺参遗传多样性的微卫星及RAPD分析的开题报告

刺参遗传多样性的微卫星及RAPD分析的开题报告
一、研究背景
刺参是我国农渔业最重要的经济海参之一,其品种众多、产地广泛,以其高蛋白、低
脂肪、高营养、滋补养生等特点而备受消费者青睐。
然而,由于过度捕捞、环境污染、海洋气候等因素的影响,刺参的资源量已经大幅减少,因此开展对刺参的遗传多样性
研究,有助于为其保护和育种提供科学依据。
二、研究目的
本研究旨在通过微卫星和RAPD技术对刺参的遗传多样性进行分析,探讨其种群结构
和遗传多样性状况,为刺参保护和育种提供科学依据。
三、研究方法
1.样本采集
本研究采用不同产地的刺参样本,共计100个。
2. DNA提取
采用组织DNA提取试剂盒对样本进行DNA提取。
3. 微卫星分析
选取10对已发表的适用于刺参的微卫星引物,采用PCR扩增。
将扩增产物进行聚丙
烯酰胺凝胶电泳分析,检测分离大小和数量和PCR产物的等位基因。
4. RAPD分析
采用10个随机引物进行RAPD扩增。
将扩增产物进行聚丙烯酰胺凝胶电泳分析,检测分离大小和数量和PCR产物的等位基因。
5.数据分析
使用POPGENE软件分析微卫星分子多态性信息,包括等位基因数、平均杂合度、平均PIC等遗传学参数。
使用WingF1软件分析RAPD分子多态性信息,包括遗传距离、相似性系数等参数。
同时,对不同种群的遗传多样性进行比较和分析。
四、研究意义
通过对刺参遗传多样性的研究,可以为刺参保护和育种的工作提供可靠的遗传学依据。
同时,也为其他经济海参的遗传多样性研究提供参考。
仿刺参(Apostichopusjaponicus)转录组分析与遗传图谱构建
谨以此文献给我敬爱的导师、父母和默默支持我的爱人杜慧霞仿刺参(Apostichopus japonicus)转录组分析与遗传图谱构建学位论文答辩日期:指导教师签字:答辩委员会成员签字:独创声明本人声明所呈交的学位论文是本人在导师指导下进行的研究工作及取得的研究成果。
据我所知,除了文中特别加以标注和致谢的地方外,论文中不包含其他人已经发表或撰写过的研究成果,也不包含未获得(注:如没有其他需要特别声明的,本栏可空)或其他教育机构的学位或证书使用过的材料。
与我一同工作的同志对本研究所做的任何贡献均已在论文中作了明确的说明并表示谢意。
学位论文作者签名:签字日期:年月日---------------------------------------------------------------------学位论文版权使用授权书本学位论文作者完全了解学校有关保留、使用学位论文的规定,并同意以下事项:1、学校有权保留并向国家有关部门或机构送交论文的复印件和磁盘,允许论文被查阅和借阅。
2、学校可以将学位论文的全部或部分内容编入有关数据库进行检索,可以采用影印、缩印或扫描等复制手段保存、汇编学位论文。
同时授权清华大学―中国学术期刊(光盘版)电子杂志社‖用于出版和编入CNKI《中国知识资源总库》,授权中国科学技术信息研究所将本学位论文收录到《中国学位论文全文数据库》。
(保密的学位论文在解密后适用本授权书)学位论文作者签名:导师签字:签字日期:年月日签字日期:年月日仿刺参(Apostichopus japonicus)转录组分析与遗传图谱构建摘要1 仿刺参的基因组大小测定本实验以栉孔扇贝Chlamys farreri为内标,用流式细胞仪测定了仿刺参Apostichopus japonicus (Selenka, 1867) 的不同组织(体壁、肌肉、肠道、呼吸树)的基因组大小(C值),结果发现用不同组织测出的仿刺参基因组大小均比较接近,最终得出仿刺参的基因组大小为0.878 ± 0.02 pg,即859 Mb(以1 pg DNA= 978 Mb计算)。
野生与养殖刺参营养成分的比较分析
C ie cdm i e c ne,Qn d o 60 hn ) hns A a e yo Fs r Si cs i a 6 7 ,C ia e f hy e g 2 1
a is sw l a 1 k d f lm ns nteb d a ew e i n utrdsa c c m e A ot hp s a o i s .T e rsl c el s1 i so ee e t i o yw l b t e nw l a dc l e e u u b r( p s co u p nc ) h eut d a n h l d u i j a s s o a i w l a d c l rd sac c mb r h o t t f ol e rt n( / h w t t n i n ut e e u u e ,tec ne l g np o i w w,dyw ih)i 3 . 3 a d4 . 7 ,rs e — h d u n oc a e r eg t s 9 4 % n 0 3 % ep c
氨 基酸含量 丰富 。
关 键 词 :刺 参 ;野 生 ;养 殖 ;营 养 成 分
中 图 分 类 号 :T 5 . S2 4 1 文 献 标 志 码 :A 文 章 编 号 :2 9 08 0 5— 7 0一( 02 0 0 6 0 2 1 ) 2— 0 4— 7
Co p r tv n l ss o u r tv o p sto e we n wid a d m a a i e a a y i f n t i e c m o ii n b t e l n i
运用形态学和分子生物学方法鉴定市场中伪劣海参产品
运用形态学和分子生物学方法鉴定市场中伪劣海参产品文菁;曾玲;王雁;田丽;周鲜娇【摘要】本研究利用扫描电子显微镜对新鲜仿刺参、市场上销售的仿刺参干品以及冷冻产品的背脊部骨片的组成和结构进行了观察.结果表明,鲜仿刺参和干仿刺参骨片形态和结构基本相似,而冷冻仿刺参骨片与它们存在明显差异.利用PCR扩增这三种海参的线粒体16S rRNA基因的部分序列,测序并研究其亲缘关系.其中鲜仿刺参和干仿刺参的序列完全相同,与之前学者报道的仿刺参序列同源性为99.8%,而冷冻仿刺参与它们的同源性仅为81.5%.进化树表明,鲜仿刺参和干仿刺参关系最近,冷冻仿刺参与它们关系较远.形态学和分子生物学方法都证明,干仿刺参产品为正宗仿刺参,冷冻仿刺参为假冒仿刺参.【期刊名称】《湛江师范学院学报》【年(卷),期】2012(033)003【总页数】5页(P95-99)【关键词】仿刺参;扫描电镜;骨片;16S rRNA;鉴定【作者】文菁;曾玲;王雁;田丽;周鲜娇【作者单位】湛江师范学院生命科学与技术学院,广东湛江524048【正文语种】中文【中图分类】Q178海参属于棘皮动物门(Echinodermata)、海参纲(Holothuroidea),是名贵的珍馐海味,名列于八大海珍之中,是我国传统的滋补品,具有补肾益精,生肌止血之功能,素有“海中人参”之称.海参体壁含有丰富的胶原成分和蛋白聚糖,并含有钙、镁盐及铁、锰等多种微量元素[1].随着人民生活水平的提高和保健意识的增强,海参的医疗保健作用得到广泛的认同和应用.仿刺参(Apostichopus japonicus)产于我国北方的辽宁、河北和山东,目前为我国海参的主要养殖品种,其中2000年我国仿刺参干品的产量为5800t[2].新鲜刺参往往根据体型和体色等外观因素肉眼鉴别,但海参往往加工成冷冻产品或干品,然后在市场上出售.深度加工严重损害了某些形态学特征,使得肉眼辨别海参产品真伪变得困难.在利益的驱使下,不法商贩对海参食品掺假,非法贴标,和用品质稍差的海参品种替换品质优良品种出售,使消费者蒙受欺骗和经济损失.因此,急需快速、可靠、方便的分析方法和技术对海参品种加以鉴别.近年来,国外已有不少报道运用分子生物学手段对鱼类加工食品中的物种进行鉴定[3-5].目前,关于海参食品的检测方法和技术尚未见报道,本研究利用电镜技术和序列测定技术对市场上出售的仿刺参食品加以鉴定.1 材料与方法1.1 主要材料、试剂与仪器新鲜仿刺参10头取自大连仿刺参养殖基地,参照廖玉麟[6]的描述加以鉴定.冷冻仿刺参10头和干仿刺参10头采购于广州本地水产市场.无水乙醇(分析纯)广州化学试剂厂;次氯酸钠(分析纯)广州化学试剂厂;海洋动物组织DNA提取试剂盒北京天根生物公司;Taq酶北京天根生物公司;dNTPs上海生工生物公司;AxyPrepTM DNA纯化试剂盒杭州爱思进生物公司;E-1010型离子溅射仪日本日立公司;S-3400N扫描电镜日本日立公司;U-1800分光光度计日本日立公司;PTC-100PCR仪美国BioRad公司.1.2 方法1.2.1 骨片扫描电镜制样从三种海参样品背脊中部,切取1cm2×1mm的体壁组织,每个样品3个重复,双蒸水洗净,灭菌吸水纸吸干水分,至于5mL小烧杯中(每个样品配有单独的烧杯和吸管,防止相互污染),滴加1mL 10%NaClO溶液消化30s左右,挑出未消化的组织块,滴加3mL双蒸水,待白色骨片沉淀,吸去上层溶液,滴加双蒸水,重复漂洗3次.吸取骨片悬液,小心滴加在滤纸上,65℃烘干4h,轻柔抖动滤纸,使骨片散落至胶膜上,日立公司E-1010型离子溅射仪喷镀金膜,用日立公司S-3400N扫描电镜观察并摄影.1.2.2 DNA提取剪取海参样品肌肉组织约30mg,用双蒸水洗净,灭菌吸水纸吸干水分,按照海洋动物组织DNA提取试剂盒流程提取基因组DNA,溶解于50μL TE,4℃保存.1.2.3 聚合酶链式反应(PCR)参照Kerr[7]的引物(16Sar:5’-CGCCTGTTTATCAAAAACAT-3’,16Sbr:5’-CTCCGGTTTGAACTCA GATCA-3’),以基因组DNA为模板,扩增海参样品线粒体16SrRNA基因约530个碱基片段.反应体系:总反应体积50μL,包括10×PCR缓冲液5μL(内含 Mg2+50mmol/L)、1μL dNTPs(10mmol/L)、2μL每种引物(10μmoL/L)、40ng DNA模板、0.5μl Taq酶(2U/μL),灭菌的双蒸水补充至50μL.PCR反应程序为:95℃变性30s,然后95℃30s、50℃退火30s、72℃1min,经40个循环,72℃延伸4min.1.2.4 PCR产物凝胶成像,纯化,测序PCR产物用1.2%琼脂糖凝胶电泳,Goldview染色,凝胶成像系统观察、照相.从电泳结果看,引物均能扩增出纯度较高的惟一的目的片段.PCR产物经试剂盒纯化后,送广州英骏生物公司双向测序.1.2.5 数据处理双向测序后用DNASTAR和Clustal X软件进行拼接和比对,为了明确本实验所用样品与其它刺参的同源性差异和系统发生关系,从GenBank中选取5个种类的刺参(见表1)与本实验结果序列比较,用MEGA软件分析它们的遗传距离,从GenBank中选取海胆Arbaxia lixula为外群,用PHYLIP软件构建了系统树.表1 源自GenBank中的5种刺参及1种海胆的基本信息属名Genus种名Species缩写Abbreviation登录号Accession No仿刺参属Apostichopus 仿刺参Apostichopus japonicus A.japo AY852278刺参属Stichopus 花刺参Stichopus hermanni S.herm FJ589203刺参属Stichopus 绿刺参Stichopus chloronotus S.chlo FJ589204梅花参属Thelenota 梅花参Thelenota ananas T.ananas FJ589205梅花参属Thelenota 巨梅花参Thelenota anax T.anaxFJ589206 Arbaxia Arbaxia lixula Ar.lixu X803962 结果与分析2.1 3种仿刺参骨片的扫描电镜结果3种仿刺参背脊部骨片见图1.其中在新鲜的仿刺参和干仿刺参中观察到1种骨片,即退化的桌形体.桌形体底盘近圆形,直径在55μm左右,周缘平滑或呈波状,底盘分布有若干个不规则的小孔.桌形体塔部变低或消失(图1,C),顶部中央有1大孔(图1,A),或没有(图1,B).在冷冻仿刺参中观察到2种骨片,桌形体和花纹样体.桌形体底盘近圆形,直径在80μm左右,周缘呈波齿状,底盘分布有数十个不规则的小孔.桌形体塔部较高,约为70μm,有4~5个立柱和2~3个横梁,各立柱上有若干个大棘.桌形体顶端不规则,但不愈合为单尖,常有3~5个尖(图1,D)或多个尖(图1,E).花纹样体骨长在75μm左右,主枝干粗长,长约60 μm,宽约10μm,一般分枝1~2次,末端形成瘤状结构(图1,F).比较发现,新鲜仿刺参和干品仿刺参骨片结构基本相似,此结果也与廖玉麟[6]的描述基本一致,可以初步判定为同一种,即仿刺参.但与冷冻仿刺参骨片形态和结构存在明显差异.主要表现在:(1)桌形体大小不同,冷冻仿刺参桌形体明显大于前两者;(2)桌形体结构不同,前两者桌形体底盘小孔明显少于冷冻仿刺参,前两者桌形体塔部退化,甚至顶端消失,后者塔部高,顶端分布多个尖刺;(3)冷冻仿刺参存在花纹样体,而前两者没有.因此,可以初步判定此类冷冻仿刺参非真正的仿刺参,为假冒产品.图1 3种仿刺参样品骨片扫描电镜图A.鲜仿刺参桌形体;B.干仿刺参桌形体;C.干仿刺参桌形体;D.冷冻仿刺参桌形体;E.冷冻仿刺参桌形体;F.冷冻仿刺参花纹样体2.2 3种仿刺参16SrRNA基因片段序列比对结果从三种海参扩增的DNA片段大小在530bp左右,经比对及剪切引物,鲜仿刺参和干仿刺参序列的碱基长度为530bp,冷冻仿刺参为528bp.相似性比较发现,鲜仿刺参和干仿刺参两个个体的序列完全一致,序列已递交GenBank,收录序列号为FJ589207.仿刺参的16SrRNA基因序列在GenBank中已有记录(表1),和李颖[8]报道的序列结果相比较发现,它们仅在第329个碱基处出现基因转换(G-A),同源性达到99.8%.而与冷冻仿刺参的序列比对发现,不仅长度不同,而且存在14个插入缺失位点,48个碱基转换位点以及36个碱基颠换位点,同源性仅为81.5%.因此,可以进一步判定干品为仿刺参,冷冻产品非仿刺参.2.3 不同刺参种间的遗传距离比较为比较本研究3种仿刺参产品与其他刺参种间的遗传距离及进化关系,从GenBank中检索到5种刺参科海参的16SrRNA基因部分序列共7种刺参材料.利用MEGA 4.1软件的双参数法,以转换加颠换计算相对遗传距离,见表2.从遗传距离分析结果可以看到,梅花参与冷冻仿刺参的遗传距离最大,为0.256,干仿刺参与仿刺参遗传距离最小,为0.002,远低于不同种间的距离,因此可以判断它们为同一物种,即仿刺参,这一结果也说明,16SrRNA的遗传稳定性相当高.而冷冻仿刺参与仿刺参的遗传距离为0.220,大于0.1,甚至大于其它种间的遗传距离,因此可以说明它们为两个种,也判定该冷冻仿刺参产品为假冒仿刺参.表2 基于Kimura双参数模型的7种刺参遗传距离注:Ap.japo1为鲜仿刺参和干仿刺参样品序列,Ap.japo2为冷冻仿刺参样品序列序列(1)(2)(3)(4)(5)(6)Sequence(1)S.herm(2)S.chlo 0.146(3)Ap.japo2 0.1920.203(4)Ap.japo 0.244 0.223 0.220(5)Ap.japo1 0.244 0.223 0.220 0.002(6)T.ananas 0.238 0.235 0.256 0.176 0.178(7)T.anax 0.230 0.244 0.235 0.181 0.184 0.1102.4 不同刺参种间的亲缘关系分析以海胆Arbaxia lixula为外群,用PHYLIP软件构建了NJ和MP系统树,结点的数值是Bootstrap 1000次重复抽样检验的结果.结果表明,两者的拓扑结构完全一致(图2).结果说明了这7种刺参惟一的系统分类关系.结果表明,干仿刺参和仿刺参聚为一支(支持率为100%),表明它们的亲缘关系最近,同属于一个属,即仿刺参属,而目前知道的这个属只有一个种,即仿刺参,因此我们可以判定该干仿刺参产品为仿刺参.冷冻仿刺参与刺参属的两个种(花刺参和绿刺参)关系较为密切(支持率为95%),明显疏远了仿刺参,因此我们可以断定该冷冻产品不是仿刺参.而它究竟是什么种尚无定论,改种目前在国内外尚无记载,我们通过Genbank搜索同源序列比较发现,它与Isostichopus fuscus(序列号AY 153499)的相似性为94%,与Isostichopus badionotus(序列号EU 822435)的相似性为92%,因此我们可以初步认为,该种可能属于刺参科的Isostichopus 属.图2 NJ和MP系统树节点上的数字分别代表NJ法和MP法1000次重复的自展值百分比3 讨论海参是我国的传统美食,骨片是位于海参真皮表层的海参内骨骼,是进行海参分类及快速鉴别的重要特征[9].李贇[10]等研究了十五种海参干品背脊部骨片的形态学,认为骨片组成特征相当稳定,可作为海参快速鉴定的理想指标.线粒体DNA 结构简单,在细胞中含量丰富,其中,核糖体RNA基因进化速度最慢,高度保守,常用于种或种以上水平的检测.国内外已有学者利用16SrRNA基因来研究海参的系统水平发生[7][8][11][12].本研究利用扫描电子显微镜和PCR技术,对新鲜仿刺参样品和市场上的仿刺参产品背脊骨片进行了细微的结构观察,分别扩增和测序了它们的16SrRNA基因序列,形态学特征和分子生物学证据都表明,干仿刺参产品为仿刺参,冷冻仿刺参产品为假冒仿刺参,商家采取以次充好,乱贴标签,蒙蔽消费者,从中牟取利益.本研究也说明,这两种技术结合在一起可以作为快速可靠的方法来鉴定市场中的海参商品,以维护消费者的合法权益.同时,我们也呼吁有关部门,对于食品中的假冒伪劣产品给予严厉打击,强调食品安全,建立健康,稳定,和谐的食品市场.参考文献:【相关文献】[1]樊绘曾.海参:海中人参,关于海参及其成分保健医疗功能的研究与开发[J]中国海洋药物,2001(4):37-44.[2]Chen J X.Present status and prospects of sea cucumber industry in China[C]∥ Lovatelli A,Conand C,Purcell,S,Uthicke S,Hamel J F,Mercier A(Eds.),Advancesin sea cucumber aquaculture and management.FAO,Rome,2004,pp:25-38.[3]Zhang J,Wang H,Cai Z.The application of DGGE and AFLP-derived SCAR for discrimination between Atlantic salmon(Salmo salar)and rainbow trout(Oncorhynchus mykiss)[J]Food Control,2007,18:672-676.[4]Rea S,Storani G,Mascaro N,Stocchi R,Loschi A R.Species identi?cation in anchovy pastes from the market by PCR-RFLP technique[J]Food Control,2009,20:515-520.[5]Comi G,Iacumin L,Rantsiou K,Cantoni C,Cocolin L.Molecular methods for the differentiation of species used in production of cod-fish can detect commercial frauds[J]Food Control,2005(16):37-42.[6]廖玉麟.中国动物志,棘皮动物门,海参纲[M]北京:科学出版社,1997.[7]Kerr A M,Kim J.Molecular Phylogeny of Coral-Reef Sea Cucumbers (Holothuriidae:Aspidochirotida)Based on 16S Mitochondrial Ribosomal DNA Sequence [J]Mar Biotechnol,2005(7):53-60.[8]李颖,刘萍,孙慧玲,等.仿刺参(Apostichopus japonicus)mt DNA三个基因片段的序列分析[J]海洋与湖沼,2006,37:143-153.[9]廖玉麟.我国的海参[J]生物学通报,2001,35(9):124.[10]李贇,费来华,陈家鑫,等.十五种海参骨片的形态学研究[J]中国海洋大学学报,2008(38):211-216.[11]Arndt A,Marquez C,Lambert P,et al.Molecular phylogeny of eastern pacific sea cucumbers(Echinodermata:Holothuroidea)based on mitochondrial DNA sequence[J]Mol Phylogen Evol,1996,6(3):425-437.[12]费来华,李贇,陈家鑫.10种海参16SrDNA序列多样性及其亲缘关系分析[J]中国水产科学,2008(15):755-765.。
中科院遗传所研究成果
目
录
一.实验室概况 ............................................................................................................................. 1 二.科研工作进展和成果 ............................................................................................................. 6 高等植物表观遗传学研究(曹晓风课题组) ......................................................................... 6 植物比较基因组学研究(陈明生课题组) ........................................................................... 11 植物对非生物胁迫应答调控的分子机制(陈受宜课题组) ............................................... 14 植物分子细胞遗传(程祝宽课题组) ................................................................................... 18 植物基因表达调控(储成才课题组) ................................................................................... 23 基因表达调控和植物生物技术(方荣祥课题组) ............................................................... 28 RNA 沉默和植物抗病机制(郭惠珊课题组) ...................................................................... 38 植物转录调控网络研究(焦雨铃课题组) ........................................................................... 43 茉莉酸的生理功能及作用机理研究(李传友课题组) ....................................................... 46 水稻理想株型基因的克隆与功能研究(李家洋课题组) ................................................... 50 植物对病原微生物的识别及信号转导(邱金龙课题组) ................................................... 53 植物天然产物代谢(王国栋课题组) ................................................................................... 56 生物信息学和系统生物学(王秀杰课题组) ....................................................................... 59 与植物重要农艺性状相关基因的结构和功能研究(夏桂先课题组) ............................... 62 植物胁迫信号传导的分子机制(谢旗课题组) ................................................................... 66 北方粳稻耐逆性的分子设计和新品种选育(姚善国课题组) ........................................... 70 乙烯信号传递与植物胁迫和生长发育反应(张劲松课题组) ........................................... 73 植物细胞壁形成及其生物学功能研究(周奕华课题组) ................................................... 77 水稻分化发育和抗病性的功能基因组研究(朱立煌课题组) ........................................... 82 植物遗传工程研究(朱祯课题组) ....................................................................................... 88 细胞分裂素信号转导和植物细胞的程序性死亡(左建儒课题组) ................................... 92 承担课题及当年经费到位情况 ............................................................................................... 95 三.人员情况 ............................................................................................................................. 119 四.学术交流 ............................................................................................................................. 129 五.运行管理 ............................................................................................................................. 141 六.2010 年学术年会纪要 ........................................................................................................ 145
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
表达量约是青刺参的 2倍, 而虾青素和 小眼畸形转录因子在白化刺参中相对表达量则低于青刺 参, 分别为青刺参的0 . 3 8 倍和0 . 4 7 倍, 相对表达量趋势和所得结果一致. 该研究为进一步了
解刺参 体 色分化及 其优 良性 状 的形成机 制提 供 了基础 资料 .
关键 词 :仿 刺 参;体 色遗传 ;抑制 消减 杂 交;荧 光定量 P C R
部为白色的刺参称之为白化刺参. 白化刺 参 以其生长 决 、 抗 逆性 强等优 势被 为
是 优 良 的刺 参 品种 .野 生 刺 参 白化 是 一 种 比较 少 见 的现 象 ,其 与 普 通 刺参 的 外观 区别 主 要 表 现 为
白化 刺参 和 青刺参 各 2 0 个 个体 于 2 0 1 2 年 3月 采集 于 辽 宁 省锦 州 市 某 刺参 育 苗 场 ,湿 重分 别 为 ( 9 5 + 1 2 ) g和( 8 5 + 1 2 ) g ,活体低温 带 回实验 室 .
中表达的情况,白化刺参中 2 种基l J 大 相对表达量均 低于普通刺参.为识别 白化 刺参优 良性状 的分 子
生 物 学 基 础 ,笔 者构 建 了白化 设计育种提供参考资料.
1 材 料 与方法
1 . 1 材料 1 . 1 . 1实验样本
1 . 1 . 2 主 要 试 剂
全 身或局部体表呈 白色.已有研究表明 白化 刺参 繁殖 的后代仍具有 白化特征, 说 明刺参 白化具有 定的遗传性,因此从基因层面研究刺参 白化, 有 利于我们更好 地了解 刺参 白化的机理, 从而更好
一
实 验 用 R NAi s o P l u s ,P C R — S e l e c t c D NA S u b t r a c t i o n Ki t 。 M— ML V c D NA S y n t h e s i s Ki t , 单链
收稿 日期 :2 0 1 3 — 0 4 — 0 8 . 宁 波大 学学 报 ( 理 工版 )网址 : h t t p : / / j o u r n a l l g n b u . e d u c n / 基金项 目: 浙汀 省科技 厅海 水养殖 创新 团1 人 , 项目 ( 2 0 1 2 R1 0 0 2 5 — 0 9 ) 第~ 作者 : 朱 琳 ( 1 9 8 6 一) ,男,山东 高密 人, 在 读 硕士研 究生 ,主要研 究方 : 海 洋生 物技术 . E - m a i l : z 1 2 7 8 3 6 8 7 @1 2 6 . c o m 通信 作者 :李成 华 ( 1 9 7 8 一 ),男, t l l 东济 宁人 . 博 十, 副研 究 员,主要研 究方 向 : 海 洋生物 技 术. E - m a i l : l i c h e n g h u a @n b u . e d u c n
基于抑制消减杂交识别 白化刺参和青刺参差异表达基 因
朱 琳,郭春 阳,王 振,李成华
( 宁波 大学 海洋学 院, 浙江 宁波 3 1 5 2 1 1 )
摘 要 :为探 寻 刺参体 色遗传 的 内在 分子 基础 ,运 用 C l o n t e c h S S H 试 剂盒分 别构 建 了白化刺 参和 青 了体 色遗传 差 异性表 达基 因 2 6 个: GO分析 显 示这 些差异基 因分别 隶属 于核糖 体基 因、能量代谢 、细胞 结构 、免疫 调控 、白化 性 状直接 相 关基 因及 未知基 因.荧光 定量 P C R 结 果显示 : AD P 一 核糖 激化 因子在 白化 刺参 中相 对
c D NA 合 成 试剂 盒( P r i me S c r i p t 1 s t S t r a n d c D NA
地对 刺参 的生 长进 行人 _ T干预 ,增加 刺参 的品质 .
有研究表明,黑色素细胞或黑色素生成发生 障碍是导致 白化的直接原因.目前 已经展 开对 刺 参白化在基因层面上研究, 赵鹤凌¨ 2 J 比较了虾青素 及小眼畸形相关转录因子在 白化刺参和普通刺参
S y n t h e s i s K i t ) , p MD 1 8 . T载体和 P C R扩增试剂均 购 自大连宝生生物公司; P C R 产物 回收试剂盒为 Q I A G E N公司产品, D H 5  ̄ t 感受态细胞 由实验室 自 行制备, 其他试剂均为分析纯.
中图分 类号 : Q 7 8 5 文献 标志 码 : A 文 章编号 : 1 0 0 1 — 5 l 3 2( 2 0 1 4)0 1 — 0 0 0 8 — 0 5
刺参 又称仿 刺参 ( A p o s t i c h o p u s j a p o n i c u s ,
S e l e n k a ) , 属棘皮动物f J , 海参纲, 楣手 目 , 刺参科, 仿刺参属, 是我国重要的水产养殖 品种…. 刺参主 要分布于我国辽宁 、山东 、 河北等省浅海沿岸, 具 有补肾 、 益精 、养血之功能, 营养价值极高. 依据 其体表颜色的差异, 人们通常把棕色 、 青色或褐色 的刺参称之为青刺参, 体表颜色局 部为白色或全
第2 7 卷第i 期, 2 0 1 4 年1 月
Vo l - 2 7 No . 1 . J a n 2 0l 4
宁 波 大 学 学 报 (理 工 版 )
J O UR NAL O F N I N GB O U NI VE R S I T Y( NS E E)
首 届 中国高 校优 秀科 技期 刊奖 浙汀 省优 秀科 技期 刊一 等奖