2019年色氨酸操纵子.ppt
原核表达调控与色氨酸操纵子优秀课件

1、不依赖于ρ因子的终止子(强终止子)
• 结构特点:RNA具有一个发夹结构(富含GC) 和随后polyU片段。
• 作用机制:RNA pol + 发夹结构 → 转录暂停→ 后随杂合分子不稳定的poly(U-dA) → RNA容易 从模板上脱落→RNA-DNA-RNA pol 解聚 → 转 录终止
-10
+1
+10
stream
start point
downstream
一、转录的起始
• 关键:全酶能以很高的亲和性结合在启 动子promoter ;
• 启动子:RNA聚合酶识别、结合并起始 转录的一段DNA序列。
转录的起始
• 核心酶在σ因子帮助下特异性结合到DNA上; • RNA pol与启动子-35 box 结合,形成封闭型起
RNA polymerase
holoenzyme and
DNA
a promoter
Incorporating the first few Nt
二、原核生物转录的延伸
• RNA pol沿着模板链移动,RNA链不断延伸, 并保持三元复合物的结构;
• 转录泡中的DNA螺旋前开、后合,RNA延长, 不断脱离DNA;
• RNA的转录包括promotion,elongation, termination 三过程;
• 从promoter到terminator称为transcriptional unit; • 原核生物中的转录单位多为 polycistron ; • 转录起始点记为+1,其上游记为负值,下游记为正值。
原核表达调控与色氨酸操纵子

开发高效原核表达系统
继续优化原核表达系统,提高外源基因在原核细 胞中的表达效率和稳定性,降低生产成本,推动 基因工程药物和疫苗的广泛应用。
深入解析代谢途径调控
深入研究色氨酸等代谢途径的调控机制,揭示代 谢物与基因表达的相互作用关系,为代谢工程和 合成生物学提供新的理论和方法。
04
研究方法和实验结果
介绍本研究所采用的研究方法、实验设计和实验结果 ,以及对实验数据的分析和解读。
05
研究意义和展望
总结本研究的意义和贡献,以及对未来研究方向的展 望和建议。
02
原核表达调控概述
原核生物基因表达特点
80%
转录与翻译偶联
原核生物的转录和翻译过程在时 间和空间上紧密偶联,转录未完 成时翻译已经开始。
催化RNA合成的酶,其活性受到多种因子的调控。
其他调控因子
03
如小分子代谢物、环境因素(温度、pH值)等也能影响原核生
物的基因表达。
03
色氨酸操纵子结构与功能
色氨酸操纵子结构组成
阻遏蛋白
结合到操纵基因上,阻止RNA聚合酶的结合 和转录的起始。
结构基因
编码色氨酸生物合成所需的酶。
操纵基因
与阻遏蛋白结合的区域,控制转录的起始。
转化与筛选
将重组质粒转化入宿主细胞, 通过选择性培养基筛选阳性克 隆。
诱导表达
在含有合适诱导剂的培养基中 培养阳性克隆,诱导目标蛋白 的表达。
表达产物检测
通过SDS-PAGE、Western blot等方法检测目标蛋白的表 达情况。
数据收集和处理方法
色氨酸操纵子
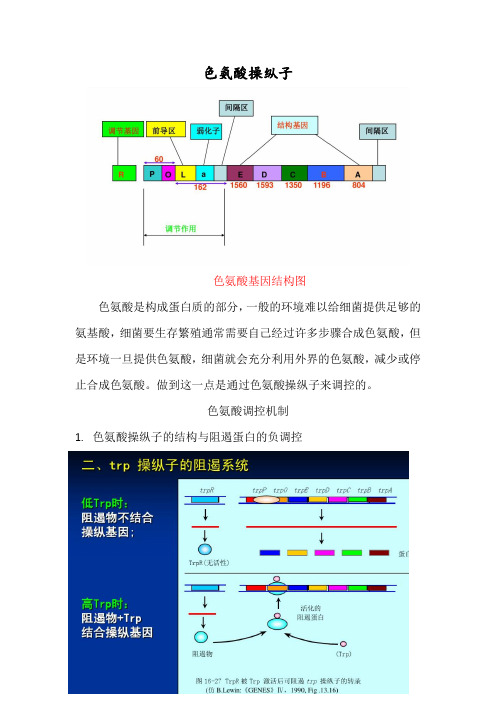
色氨酸操纵子
色氨酸基因结构图
色氨酸是构成蛋白质的部分,一般的环境难以给细菌提供足够的氨基酸,细菌要生存繁殖通常需要自己经过许多步骤合成色氨酸,但是环境一旦提供色氨酸,细菌就会充分利用外界的色氨酸,减少或停止合成色氨酸。
做到这一点是通过色氨酸操纵子来调控的。
色氨酸调控机制
1.色氨酸操纵子的结构与阻遏蛋白的负调控
如图所示:在调控色氨酸合成的结构基因上游有一个操纵基因trpR ●在低色氨酸浓度时,trpR控制的阻遏蛋白无活性,下游的结构基
因可正常转录翻译。
●在高色氨酸浓度时,trpR控制的阻遏蛋白具有活性。
能与trpO特
异性结合,阻遏结构基因的转录。
从而阻遏体内的色氨酸合成。
2.衰减子的作用
当色氨酸达到一定程度,但没有高到能够活化阻遏蛋白使其起阻遏作用的程度时,产生色氨酸合成酶类的量已经明显降低,靠着衰减子来调控。
如图所示:在高色氨酸时,trp mRNA在第一个trp E基因开始转录之前即停止生长。
低色氨酸时,mRNA正常转录。
这是因为在色氨酸操纵元trp O与第一个结构基因trp E 之间有一段前导序列。
高色氨酸时转录就会停止在这里。
如图所示:
在低浓度色氨酸条件下,2-3形成发卡结构,不含有U区域,不会形成终止子结构,不会停止转录,继续转录翻译形成色氨酸在高浓度色氨酸条件下,3-4会形成发卡结构,含有U区域,形成终止子结构,停止转录,阻遏色氨酸的合成。
第2节 色氨酸操纵子

内容提要: 色氨酸操纵子的结构 色氨酸操纵子的阻遏系统 色氨酸操纵子的弱化机制
一、色氨酸操纵子的结构
调控基因
结构基因
trpR
催化分枝酸转变为色氨酸的酶
分支酸 → 邻氨基苯甲酸 → 磷酸核糖基 → CDRP → 吲哚甘油-磷酸 → 色氨酸 邻氨基苯甲酸
邻氨基苯甲酸合成酶
RNA聚合酶 结构基因
5’
前导肽
23
核1 糖体
2 43
4
UUUU…U…UUU……
trp 密码子 序列3、4不能形成衰减子结构
2.当色氨酸浓度低时
High Trp Low Trp
弱化机制
高Trp时: Trp-tRNATrp 存在
核糖体通过片段1(2个Trp密码子) 封闭片段2
片段3,4形成发夹结构 类似于不依赖ρ因子的转录终止序列
Leader peptide
夹结构 / 富含 C G
U 的单链末端 C G
Aaaaaa C G
Met Lys Aly Ile Phe Val Leu Lys Gly Trp Trp Arg Thr Ser
A
GC
CG
A
CG
UU
AA
图 16-28 trp 操纵子含有 5 个结构基因和 1 个控制区。控制区由启动子、操纵基因、前导顺序和衰减子 构成。前导区编码 14 个氨基酸,其中有 2 个是色氨酸。(仿 B.Lewin:《GENES》Ⅵ,1997, Fig .12.38)
四、原核生物转录的整体调控模式
由成群的操纵子组成的基因转录调控网络称为调 节子。通过组成调节子调控网络,对若干操纵子 及若干蛋白质的合成进行协同调控,从而达到整 体调控的目的。
色氨酸操纵子与负控阻遏系统

被称为适应性表达基因,或被称为奢侈基因
基因的表达调控方式
➢ 基因水平的调控
➢ 转录水平的调控
➢ 转录产物加工的调控
➢ 翻译水平的调控以及翻译后的加工等
原核基因表达调控分类
根据调控机制: 负转录调控 调节基因编码阻遏蛋白,阻止结构基因转 录 分为负控诱导,负控阻遏 正转录调控 调节基因编码激活蛋白,促进结构基因转 录。 分为正控诱导,正控阻遏
鼠伤寒沙门氏菌中已陆续发现不少操纵子都有弱化
致转录终止。当色氨酸浓度较低时,TRAP失活,转录可以继
现象。 弱化子(attenuator)是指原核生物操纵子中能显著 减弱甚至终止转录作用的一段核苷酸序列,该区域
续,结构基因得以表达。另外枯草杆菌对未负荷色氨酸的 tRNATrp也很敏感,后者大量堆积,会诱导合成抗TRAP 蛋白 (anti -PRAP,AT)。AT与Trp激活的PRAP结合,可以取消其 转录终止活性。trpG表达也受PRAP调控,活化的TRAP与和
空 白 演 示 水平可提高6倍。研究发现,当mRNA开始合成后,
除非培养基中完全不含色氨酸,否则转录总是在这 个区域终止,产生一个仅有140个核苷酸的RNA分
养基中Trp 浓度很低时,负载有Trp 的tRNATrp也就少,这样 翻译通过两个相邻色氨酸密码子的速度就会很慢,当4区被转 录完成时,核糖体滞留1区,这时的前导区结构是2 - 3配对, 不形成3 - 4配对的终止结构,所以转录可继续进行。反之,核
不受葡萄糖或cAMP-CAP的调控。
空白演示
在此输入您的封面副标题
弱化子
弱化作用
是在研究大肠杆菌的色氨酸操纵子表达弱化现象 中发现的。在trp mRNA 5,端trp正基因的起始密 码前有一个长162 bp的DNA序列称为前导区,其
第六章-色氨酸(trp)操纵子3

The trp operator is a palindromic
DNA sequence
trp操纵子转录的调控是通过Trp阻遏物实现的,它结 合于trp操纵基因序列 ,但Trp阻遏物的DNA结合活性 直接受色氨酸调控,色氨酸结合Trp阻遏物,并起着 一个效应分子的作用(也称之辅阻遏物)。
在有高浓度色氨酸存在时,Trp阻遏物-色氨酸复合物 形成一个同源二聚体,并且紧密结合于trp操纵基因 序列,因此可以阻止转录。然而当色氨酸水平低时, 缺少色氨酸的Trp阻遏物以一种非活性形式存在,不 能结合DNA。在这样的条件下,trp操纵子被RNA聚 合酶转录,同时色氨酸生物合成途径被激活。
而当培养基中色氨酸浓度高时,核糖体可顺利 通过两个相邻的色氨酸密码子,在4区被转录 之前,核糖体就到达2区,这样使2-3不能配对, 3-4区可以自由配对形成茎-环状终止子结构, 转录停止,trp操纵子中的结构基因被关闭而 不再合成色氨酸(图6-24)。所以,弱化子对 RNA聚合酶的影响依赖于前导肽翻译中核糖体 所处的位置。
转录弱化作用
转录的弱化理论认为mRNA转录的终止是通过前导 肽基因的翻译来调节的。因为在前导肽基因中有两 个相邻的色氨酸密码子,所以这个前导肽的翻译必 定对tRNATrp的浓度敏感。
当培养基中色氨酸的浓度很低时,负载有色氨酸的 tRNATrp也就少,这样翻译通过两个相邻色氨酸密 码子的速度就会很慢,当4区被转录完成时,核糖 体 才 进 行 到 1 区 ( 或 停 留 在 两 个 相 邻 的 trp 密 码 子 处),这时的前导区结构是2-3配对,不形成3-4配 对的终止结构,所以转录可继续进行,直到将trp 操纵子中的结构基因全部转录。
trpE基因是第一个被翻译的基因,和trpE紧邻的是启 动子区和操纵区。另外,前导区和弱化子区分别定名为 trpL和trpa(不是trpA)。
16.3.3 色氨酸操纵子

基础生物化学Basic Biochemistry色氨酸(Trp)操纵子色氨酸操纵子是用来编码生成色氨酸的重要元件之一。
研究表明当有足够的Trp时,操纵子自动关闭,细菌直接利用外界的Trp。
缺乏Trp时,Trp操纵子被打开,5个结构基因表达,产生3个酶催化分支酸合成为Trp。
1、阻遏蛋白的负调控合成Trp的酶,需要5 个基因E、D、C、B和A的共同编码。
这5个基因的表达受上游启动子p和操纵基因o的调控。
在远离p-o-结构基因群的位置有一个调控基因TrpR,它能低水平的表达阻遏蛋白R’。
R’并无活性,当提供足够的Trp时,Trp与R’结合使其构象改变而成为有活性形式R,R与O特异性结合,阻遏结构基因的转录。
2、Trp操纵子的衰减调控⏹前导序列编码了一个14个氨基酸的前导肽;前导肽的第10、11位是相邻的两个Trp密码子。
⏹先导序列后半段含有4个彼此互补的区域(1、2、3、4),在被转录生成mRNA时相互间能形成发夹结构。
原核生物转录和翻译几乎同时进行,当转录起始后,RNA聚合酶沿DNA转录合成mRNA,同时核糖体结合在mRNA上开始翻译。
UUUU (34)UUUU 3’34核糖体前导肽前导mRNA当色氨酸浓度高时转录衰减机制125’trp 密码子衰减子结构就是终止子可使转录前导DNAUUUU 3’RNA 聚合酶终止UUUU (34)2423UUUU ……核糖体前导肽前导mRNA15’trp 密码子结构基因前导DNA RNA 聚合酶当色氨酸浓度低时Trp 合成酶系相关结构基因被转录序列3、4不能形成衰减子结构乳糖操纵子和色氨酸操纵子的比较Lac操纵子负责营养碳源的分解,只有当需要消耗乳糖时,才通过诱导物使阻遏蛋白失活而开放,是可诱导的负调控基因;此外还存在CAP的正调控。
trp操纵子负责Trp的合成,平时开放,调节基因的产物使其关闭,是可阻遏的负调控;此外还存在翻译与转录耦联的衰减子调控手段。
第三节 色氨酸操纵子

制作:王庆容
一、结构及调控模式 1.结构 2.粗调节
3.细微调控
二、弱化子与前导肽 1.弱化子attenuator :
位于trp mRNA 5‘端trpE基因的起始密码前,调 节已启动转录的trp操纵子是否继续下去的特定区域。
2.前导肽Leader peptide:
当trp浓度较高时, trp操纵子转录产生一个 140bp的RNA分子的前导序列,它编码的一个14个 AA的短肽。
3.mRNA 前导区的序列分析 4.转录弱化作用 三、trp操纵子弱化机制的实验依据 四.小结
R
P
O
L
a
E
D
C
B
A
RNA pol. mRNA
R'
R
P
O
L
a
E
D
C
B
A
mRNA
R'
色氨酸
色氨酸合酶 α和β亚基
色氨酸操纵子各组份详图
组 成
R P O
L a
间 隔 序 列
E
D
C
B
A
碱 基 数 功 能
未知 辅 阻 遏 蛋 白
60
162
1560
邻氨 基苯 甲酸 合酶
1593
ห้องสมุดไป่ตู้1350
1196
804
粗 开 关
微调控 前导肽
邻氨基苯 吲哚甘 甲酸磷酸 油磷酸 核糖转移 合酶 酶
色氨酸合酶α 和β亚基
R
RNA pol.
间隔区 调节基因 前导区 弱化子 结构基因 间隔区
60 R P O L a 162 E 1560 D 1593 C 1350 B 1196 A 804
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
蛋白 TrpR(无活性)
高Trp时: 阻遏物+Trp 结合操纵基因
阻遏物
活化的 阻遏蛋白
(Trp)
图 16-27 TrpR 被 Trp 激活后可阻遏 trp 操纵子的转录 (仿 B.Lewin:《GENES》Ⅳ,1990, Fig .13.16)
6
色氨酸的调节
转录衰减
调节区
trpR RNA聚P合酶O
2
3
UUUU……
形成发夹结构能力强弱:
3
4
序列1/2>序列2/3>序列3/4
14
前导DNA
转录衰减机制
前导mRNA
1 5’
核糖体
RNA聚合酶
UUUU 3’
衰减子结构
就是终止子
4 可使转录 终止
3
2
34
UUUU 3U’UUU……
前导肽
trp 密码子
1.当色氨酸浓度高时
15
前导DNA 前导mRNA
Trp合成酶系相关 结构基因被转录
10
邻氨基苯 吲哚甘油 色氨酸合成酶 甲酸合成酶 磷酸合成酶
TrpE terpD trpC trpB trpA t
t’
启动子 操纵基因
前导序列 衰减子
pppN26AUGAAAGCAAUUUUCGUACUGAAGGUUGGUGGCGCACUUCCUGAN43A UUUUUUUU 富含 G-C 的发 G C
4
特点:
(1) trpR和trpABCDE不连锁;
(2) 操纵基因(操作子)和启动子部分重叠 (3) 有衰减子(attenuator)/弱化子 (4) 启动子和结构基因不直接相连,二者被
前导序列(Leader)所隔开
5
二、trp 操纵子的阻遏系统
低Trp时: 阻遏物不结合 操纵基因;
trpR
trpP trpO trpE trpD trpC trpB trpA
RNA聚合酶停止转录,产生衰减子转录产物 转录、翻译偶联,产生前导肽
19
低Trp时: Trp-tRNATrp 没有供应
核糖体翻译停止在片段1 (2个Trp密码子)
片段2,3 形成发夹结构
转录不终止
RNA聚合酶继续转录
20
细菌通过弱化作用弥补阻遏作用的不足,因为阻 遏作用只能使转录不起始,对于已经起始的转录, 只能通过弱化作用使之中途停下来。阻遏作用的信 号是细胞内色氨酸的多少;弱化作用的信号则是细 胞内载有色氨酸的tRNA的多少。它通过前导肽的翻 译来控制转录的进行,在细菌细胞内这两种作用相 辅相成,体现着生物体内周密的调控作用。
吲哚-3-甘油 磷酸合成酶
色氨酸合成酶
β链
α链
60,000 O L 间隔区 trpE
60,000 trpD
4 5,000 50,000 29,000
trpC
trpB
trpA
P
P:启动子;O:操作子; L:前导序列; E.coli trpO 的结构及其产物所催化的色氨酸合成反应
3
乳糖操纵子的基本结构与调控原理
11
2、前导序列:在trp mRNA5‘端trpE基因的起始密码 前一个长162bp的mRNA片段。
12
13
调节区
trpR
PO
前导序列
前导mRNA
1
2
结构基因
衰减子区域
3
4
UUUU……
trp 密码子 终止密码子
1
2 第141a0a、前1导1密肽码编子码UU为区Ut:Ur包…p密含…U码序U子U列U1……衰减子结构
RNA聚合酶 结构基因
5’
前导肽
23
核1 糖体
2 43
4
UUUU…U…UUU……
trp 密码子 序列3、4不能形成衰减子结构
2.当色氨酸浓度低时
16
High Trp Low Trp
17
弱化机制
18
高Trp时: Trp-tRNATrp 存在
核糖体通过片段1(2个Trp密码子) 封闭片段2
片段3,4形成发夹结构 类似于不依赖ρ因子的转录终止序列
21
色氨酸操纵子的调控机制
阻遏调控 弱化(衰减调控) ●前导序列L含有四段特殊序列,序列1前有一段前
导肽。其10-11位是连续的色氨酸; ●序列2-3或3-4可形成茎环结构,3-4之间的茎环结
构是一个转录终止子结构(衰减子) ●Trp缺乏时,核糖体的翻译停滞在11-12位色氨酸密
码处,序列2-3形成茎环结构,3-4就不能形成终止 子,结构基因转录继续进行。反之,停止。
RNA聚合酶
Trp 低时
Trp 高时 Trp
结构基因
mRNA
色氨酸操纵子
7
三、trp 操纵子的弱化机制 衰减子(attenuator)/弱化子 前导序列(leader sequence)
8
1、衰减子/弱化子:DNA中可导致转录过早终止的 一段核甘酸序列(123-150区)。
123~150
9
研究引起终止的mRNA碱基序列,发现该区mRNA通过自 我配对可以形成茎-环结构,有典型的终止子特点。
Leader peptide
夹结构 / 富含 C G
U 的单链末端 C G
Aaaaaa C G
Met Lys Ala Ile Phe Val Leu Lys Gly Trp Trp Arg Thr Ser
A
GC
CG
A
CG
UU
AA
图 16-28 trp 操纵子含有 5 个结构基因和 1 个控制区。控制区由启动子、操纵基因、前导顺序和衰减子构 成。前导区编码 14 个氨基酸,其中有 2 个是色氨酸。(仿 B.Lewin:《GENES》Ⅵ,1997, Fig .12.38)
22
重点
Байду номын сангаас 色氨酸操纵子的阻遏调控机理 色氨酸操纵子的衰减(弱化)调控机理
23
第四节 色氨酸操纵子(Trp operon)
内容提要: 色氨酸操纵子的结构 色氨酸操纵子的阻遏调控机制 色氨酸操纵子的弱化调控机制
1
一、色氨酸操纵子的结构
调控基因
结构基因
trpR
催化从分支酸到合成色氨酸所需的酶
2
分支酸 → N-氨基苯甲酸 →→→→→→吲哚-3-甘油-磷酸 → 色氨酸
邻氨基苯甲酸合成酶