分子生物学第三章
分子生物学:第3章RNA的转录习题和答案

第三章RNA的转录一、名词解释1.转录2.模板链(反义链)3.非模板链(编码链)4.不对称转录5.启动子6.转录单位7.内含子8.外显子9.sigma因子10.RNA编辑11.核酶12.gRNA 13.GU-AG规则14.转录后加工15.核内不均一RNA 16.RNA复制二、填空题1.由逆转录酶所催化的核酸合成是以_______为模板,以_______为底物,产物是_______。
2.RNA生物合成中,RNA聚合酶的活性需要_______模板,原料是_______、_______、_______、_______。
3.大肠杆菌RNA聚合酶为多亚基酶,亚基组成_______,称为_______酶,其中_______亚基组成称为核心酶,功能_______;σ亚基的功能_______。
4.用于RNA生物合成的DNA模板链称为_______或_______。
5.RNA聚合酶沿DNA模板_______方向移动,RNA合成方向_______。
6.真核生物RNA聚合酶共三种_______、_______、_______,它们分别催化_______、_______和_______的生物合成。
7.某DNA双螺旋中,单链5’… ATCGCTCGA … 3’为有意义链,若转录mRNA,其中碱其排列顺序为5’… _______… 3’。
8.能形成DNA--RNA杂交分子的生物合成过程有_______、_______。
形成的分子基础是_______。
9.DNA复制中,_______链的合成是_______的,合成的方向和复制叉移动方向相同;_______链的合成是_______的,合成的方向与复制叉方向相反。
10.一条单链DNA(+)的碱基组成A2l%、G29%,复制后,RNA聚合酶催化转录的产物的碱基组成是_______。
11.RNA聚合酶中能识别DNA模板上特定起始信号序列的亚基是_______ ,该序列部位称_______。
分子生物学 第3章 DNA复制
DNA helicase (DNA解旋酶)
利用ATP供能,解开DNA双链, 可随复制叉 的伸展向前移动
大肠杆菌中解旋酶的种类
种 类
DnaA DnaB DnaC
功 能
辨认起始点,并结合到复制起始部位 解开DNA双链 运送和协同DnaB
single-stranded binding protein (SSB, 单链结合蛋白)
是一类调节DNA分子的超螺旋水平,可改变DNA拓扑性 质的酶。对DNA分子的作用是既能水解、又能连接磷酸 二酯键。 • 拓扑异构酶 I: 切开DNA双链中的一股,使DNA在解链旋 转中不打结,DNA变为松弛状态再封闭切口。 同转录有 关 • 拓扑异构酶 II: 能切断DNA双链,使螺旋松弛。在ATP参 与下,松弛的DNA进入负超螺旋,再连接断端。同复制
3´→5´外切酶活性: 切除错配的核苷酸
5'
3' C T T C A G G A G A A G T C C G G C G 5'
3'
DNA ligase
连接DNA链3-OH末端和相邻DNA链5-P末端,形成磷 反应需要ATP。
酸二酯键,从而把两段相邻的DNA链连接成完整的链。
二、 DNA复制的过程
E. Coli DNA在15N-标记的营养液中生
长多代,使DNA双链充分标记
将15N-标记
细胞在
14N中
细胞在
14N中复
细胞在
14N中复
的E.Coli 加入14N 培 养液中
万有引力
复制1 次
制第2次
制第3次
单林娜 制作
11
DNA半保留复制的生物学意义:
DNA的半保留复制表明DNA在代谢上的稳定性,
分子生物学第三章试题及答案

分子生物学第三章试题及答案一、选择题(每题2分,共20分)1. DNA双螺旋结构是由谁提出的?A. 沃森和克里克B. 罗莎琳·富兰克林C. 詹姆斯·沃森D. 弗朗西斯·克里克答案:A2. 下列哪项不是DNA复制的特点?A. 半保留复制B. 双向复制C. 单向复制D. 需要引物答案:C3. 真核生物的转录主要发生在哪个细胞器中?A. 细胞核B. 线粒体C. 高尔基体D. 内质网答案:A4. 在蛋白质合成过程中,mRNA上的密码子与tRNA上的反密码子配对,下列哪一项配对是错误的?A. UUU - AAAB. AUG - UUAC. CUU - GAAD. GUU - CAC答案:B5. 下列哪种酶在DNA复制过程中起作用?A. DNA聚合酶B. RNA聚合酶C. 逆转录酶D. 限制性内切酶答案:A6. 基因表达调控中,转录因子的作用是什么?A. 促进DNA复制B. 促进mRNA的剪接C. 促进转录的起始D. 促进翻译的起始答案:C7. 下列哪种RNA不参与蛋白质的合成?A. mRNAB. tRNAC. rRNAD. snRNA答案:D8. 真核生物的基因表达调控中,增强子的作用是什么?A. 促进转录的起始B. 抑制转录的起始C. 促进mRNA的剪接D. 促进翻译的起始答案:A9. 在PCR技术中,变性步骤的目的是?A. 使DNA双链分离B. 使DNA单链结合引物C. 使DNA单链结合DNA聚合酶D. 使DNA单链结合tRNA答案:A10. 下列哪种分子生物学技术用于检测特定DNA序列的存在?A. PCRB. 测序C. 电泳D. 限制性内切酶分析答案:A二、填空题(每题2分,共20分)1. DNA聚合酶的主要功能是______。
答案:合成DNA链2. 在转录过程中,RNA聚合酶识别的启动子是______。
答案:DNA上的特异序列3. 真核生物的mRNA在出核之前需要进行______。
《分子生物学教学》第三章可移动的遗传因子

探讨可移动遗传因子在基因工程、基因治疗、生物 育种等领域的应用前景,以及相关的伦理和安全问 题。
02
可移动遗传因子的类型和特性
转座子的类型和特性
80%
插入序列(IS)
是细菌中最简单的转座子,能够 编码自身转座所需的酶,并能在 基因组中随机插入。
100%
转座噬菌体(Tn)
是一种复杂的转座子,带有与噬 菌体相关的基因,能够在细菌之 间水平转移。
分子生物学教学第三章可移动 的遗传因子
目
CONTENCT
录
• 引言 • 可移动遗传因子的类型和特性 • 可移动遗传因子的机制 • 可移动遗传因子的生物学意义 • 研究方法和实验技术 • 实际应用和未来展望
01
引言
目的和背景
阐述可移动遗传因子的概念
本章旨在介绍可移动遗传因子的概念,包括其定义、分类、功能 以及在生物学领域的重要性。
转录调控
可移动遗传因子如转座子和逆转 录病毒可通过插入或删除基因序 列,影响转录因子的结合和基因 表达的调控。
表观遗传学调控
某些可移动遗传因子能够影响染 色质结构和组蛋白修饰,从而参 与表观遗传学调控,改变基因的 表达模式。
在基因组进化和多样性中的作用
基因重组
可移动遗传因子通过介导基因重组事 件,促进基因组的重排和多样性产生 。
对核酸和蛋白质序列进行比对,找出同源序列和保守区域,并进行功能
注释。
02
基因表达与调控分析
利用高通量测序技术分析基因表达谱,研究基因表达的时空特异性和调
控机制。
03
生物信息学数据库与工具
利用生物信息学数据库(如GenBank、UniProt等)和在线分析工具
分子生物学第三章核酸的结构与功能

分子生物学第三章核酸的结构与功能核酸是生物体内重要的生物大分子,在维持遗传信息传递、调控基因表达和蛋白质合成等生物学过程中起着重要的作用。
本文将介绍核酸的结构和功能,包括DNA和RNA的结构、功能以及细胞中的DNA重复序列和嵌合DNA的现象。
核酸是由核苷酸单元组成的大分子。
核苷酸由一糖分子(核糖或脱氧核糖),一个含有一键磷酸基的磷酸基团和一个含有碱基的碱基组成。
DNA的碱基包括腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)和胞嘧啶(C),而RNA的碱基包括腺嘌呤(A)、鸟嘌呤(G)、尿嘧啶(U)和胞嘧啶(C)。
DNA(去氧核糖核酸)是双链结构,由两条互补的单链以互补碱基配对(A和T,G和C)的方式相互连接而成。
这种双链结构被称为双螺旋结构,其中的两个链通过氢键相互链接。
DNA在细胞中起着存储遗传信息的作用,是遗传物质的主要组成部分。
DNA通过转录过程产生RNA分子,进而通过翻译过程合成蛋白质。
RNA(核糖核酸)有多种类型,包括信使RNA(mRNA)、核糖体RNA (rRNA)和转运RNA(tRNA)。
mRNA是由DNA转录得到的,其中的密码子序列编码蛋白质的氨基酸序列。
rRNA是核糖体的组成部分,参与蛋白质合成的过程。
tRNA将氨基酸带入核糖体与mRNA相匹配的密码子上,完成蛋白质合成的过程。
在细胞中,存在许多DNA重复序列。
其中,基因是密集编码蛋白质的DNA序列,它们在核酸的遗传信息传递和基因表达中起着重要作用。
除了基因,还存在大量的非编码DNA序列,如内含子和调控序列,它们对基因表达的调控起着重要作用。
此外,DNA重复序列还包括微卫星、线粒体DNA和细胞质DNA等。
总之,核酸是生物大分子,在维持遗传信息传递和调控基因表达等生物学过程中起着重要作用。
DNA和RNA具有不同的结构和功能,包括存储遗传信息、编码蛋白质序列、调控基因表达和蛋白质合成等。
此外,细胞中存在多种形式的DNA重复序列和嵌合DNA现象,对维持细胞功能和遗传多样性具有重要作用。
分子生物学第三章蛋白质大分子结构与功能

49
4.蛋白质四级构造
一个蛋白质由几条多肽链组成1个活性单 位。亚基的相互关系,空间排布,亚基 间通过非共价键聚合成的特定构象。单 一亚基无活性,只有聚合后才有生物活 性。如血红蛋白。
50
51
蛋白质预测网站
Compute pI/WM ://expasy.hcuge.ch Predictprotein :// embl-heidelberg.de/predictprotein/ SOPMA :// ibcp.fr/predict.html Unpredict ://www /
43
44
45
46
构造域
• 多肽链在超二级构造根底上进一步卷 曲折叠成严密的近似球状的构造。对 较小蛋白质分子,构造域往往就是三 级构造,即这些蛋白质是单构造域。
• 许多蛋白质是多构造域。
47
构造域
48
3.蛋白质三级构造
• 多肽链的某些区域氨基酸形成二级构造: α螺旋、β-折叠、 β-转角、无规那么卷曲等构 象单元, 然后相邻二级构造集装成超二级构 造, 进而折叠绕曲成构造域, 由2个或2个以上 的构造域组装成三级构造。
30
二 硫 键
31
一级构造确定的原那么
• 测定蛋白质中氨基酸组成 • 蛋白质N端和C端的测定 • 2种以上方法水解蛋白质,得到一系列
肽段 • 别离提纯所得肽,测其序列 • 从有重叠构造的肽序列中推断蛋白质
的全部氨基酸的排列顺序
32
〔二〕蛋白质的空间构造
• 肽平面:肽腱中的4个原子以及相邻的 2个α-碳原子处在同一平面,使肽链具 有一定的稳定性
20
氨基酸与异硫氰酸苯酯的反响
• AA的氨基可与异硫氰酸苯酯(PITC)反 响,生成苯氨基硫甲酰氨基酸(PTC-AA)。 所得PTC-AA经乙酸乙酯抽提→层析鉴 定→ 确定N端氨基酸的种类。 “多肽 顺序自动分析仪〞据此原理。
分子生物学第三章RNA转录

分⼦⽣物学第三章RNA转录第三章 RNA 转录(RNA transcription)3.1. Basic concept3.2. Trancription survey3.3. Promoter in Eukaryotes and Prokaryotes3.4. Transcription Termination3.5. Pre-RNA processing in Eukaryotes3.1. 基本概念(P64) Basic concept●基因表达的第⼀步●以D. S. DNA 中的⼀条单链作为转录的模板某⼀基因只以⼀条单链DNA 为模板进⾏转录(不对称转录)●在依赖DNA 的RNA 聚合酶的作⽤下●按A U ,C G 配对的原则,合成RNA 分⼦●模板单链 DNA 的极性⽅向为3’ → 5’, ⽽⾮模板单链DNA 的极性⽅向与RNA 链相同,均为5’ → 3’.● RNA 的转录包括promotion, elongation, termination 三个阶段●从启动⼦(promoter )到终⽌⼦(terminator )的DNA序列称为转录单位(transcriptional unit )●原核⽣物中的转录单位多为 polycistron in operon真核⽣物中的转录单位多为monocistron, No operon●转录原点记为+1,其上游记为负值,下游记为正值● RNA 的主要种类及功能:mRNA ——携带编码多肽的遗传信息tRNA ——将核苷酸信息转化为aa 信息转运aa 进⼊核糖体rRNA ——参与多肽合成3.2.RNA 转录概况3.2.1转录的基本过程1. 模板识别:RNApol 与启动⼦相互识别并结合的过程(形成封闭的⼆元复合物)启动⼦(promoter ):DNA 分⼦上结合RNApol 并形成转录起始复合物的区域,通常也包括促进这⼀过程的调节蛋⽩结合位点rich A/T ,易发⽣DNA 呼吸现象形成单链区2转录起始:启动⼦区解链,转录起始(封闭的⼆元复合物开放的⼆元复合物三元复合物)通常在这⼀过程中RNApol 移动较慢,且易发⽣脱落——流产式起始 ——决定启动⼦的强弱3延伸:延伸过程中的延宕现象(Eukaryotes ):Euk genome G/C 分布不均匀σ脱离全酶(Pro )/RNApol 脱离转录起始复合物(Euk )4终⽌:在终⽌⼦(terminator )处停⽌转录3.2.2 RNApolymerase1 RNA polymerase in Prokaryotes (以E.coli 为例)1)构成核⼼酶(core enzyme):2αββ’DNA3’----TACTCAT----5’ RNA 5’----AUGAGUA----3’5’---ATGAGTA----3’ Non-template (sense strand)template (antisense strand)全酶(holoenzyme)2αββ’σα:核⼼酶组建因⼦/ 启动⼦识别β:RNA合成的活性中⼼β’:与β共同构成活性中⼼σ:识别启动⼦,增加酶与DNA的亲和⼒σ因⼦可减少RNApol与⾮启动⼦DNA序列的亲和⼒,⽽增加RNApol与启动⼦的亲和⼒,⼀旦转录起始,σ因⼦将脱离RNApol再次引导新的RNApol进⾏转录ρ:参与转录终⽌2)Rifamycin(利福霉素)及Streptolydigin(利链菌素)对Pro转录的影响Rif可结合β,阻⽌NTP的进⼊I位点(Initiation site )(⼀旦形成三元复合物Rif不再起抑制作⽤);利链菌素结合β的延伸位点(Elongation site),抑制延伸。
分子生物学基础第三章遗传与变异 第一节原核生物的遗传规律

第一节 原核生物的遗传规律
(3)Fˊ因子与性导 F+与Hfr两种菌株可以相互转换,也就是说F因子既可 以插入到染色体中去,形成Hfr菌株,有时又可通过有规 则的交换和剪切,从染色体上完整地游离下来形成F+菌株, 但是偶尔也会出现不规则的环出,形成的F因子携带了相 邻细菌染色体的基因(图3-6)。这种带有插入细菌基因 的环状F因子称为Fˊ因子(Fˊ-facter)。
高频重组 后来Cavalli和Hayes先后在菌株A中发现 了一种高频重组菌株Hfr。它们能跟F―的菌株杂交,并能 得到频率很高的重组细菌,频率要比一般的F+×F―高出上 千倍。经研究证明,Hfr和F+不同之处是Hfr中的F因子整 合在细菌的染色体上(图3-4B),但一般的F+中的F因子 是存在于细胞质中的质粒。
第一节 原核生物的遗传规律
图3-7 P22噬菌体的普遍性转导示意图
第一节 原核生物的遗传规律
特异性转导 我们现在介绍另一类噬菌体,它们所进 行的转导是特异性转导(specialized transduction), 或局限性转导(restricted transduction),这类噬菌 体只转移细菌染色体的特定部分。λ噬菌体是特异转导者 (transducer)的一个很好例子。大肠杆菌的一个溶源菌 株K12(λ)可由紫外光诱导,用来进行转导。唯一成功的 转导为gal+基因座位。根据实验知道,λ总是附着在供体 的gal+基因座位的邻近位置,特异性转导供体的gal+基因 给受体(图3-8)。
第一节 原核生物的遗传规律
图3-9 细菌转化的机制 A:转化菌形成的过程; B:转化中遗传重组的机制
第一节 原核生物的遗传规律
转化时供体细菌DNA断裂成小片段,这些片段平均长 度约为20,000个核苷酸对,外源DNA片段进入受体后可以 和受体染色体形成部分二倍体,有可能发生重组,从而使 受体细胞发生稳定性的遗传转化。转化过程包括几个连续 的阶段:①供体双链DNA 分子和受体细胞表面受体部位进 行可逆性结合;②供体DNA片段被吸入受体细胞,并要防 止被受体DNA酶破坏;③供体DNA进入受体后,立即从双链 DNA转变成单链DNA,其中一条单链被降解;④未被降解的 单链DNA部分地或整个地插入受体细胞的DNA链中与同源区 段形成杂合的DNA分子;⑤杂合DNA经复制、分离以后,形 成一个受体亲代类型的DNA和一个供体与受体DNA结合的杂 种双链DNA,从而导致基因重组形成各种类型的转化子 (transformant)。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
M. Meselson and F. W. Stahl, Sciences 44:675, 1958.
OK !
3’
5’
5’
3’
3’
How ?
5’
a)
Okazaki fragment E.coli [t-] 2s, 7s, 15s, 60s
1968 Reiji Okazaki
20℃ in dT-H3 30s pulse-chase
• Replisome
The multi protein (30±) structure that assembles at replicating fork to undertake synthesis of DNA
DNA replication at phase S of cell cycle
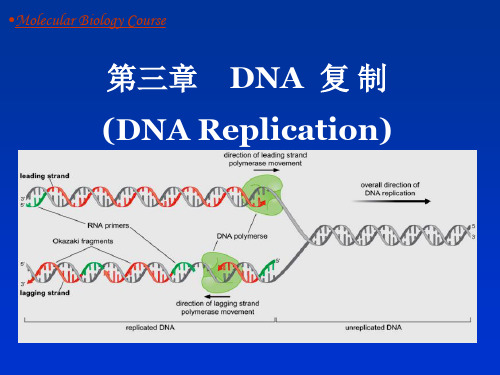
DNA复制
亲代双链DNA分子在DNA聚合酶的作用下,分
别以每单链 DNA分子为模板,聚合与自身碱基
可以互补配对的游离的dNTP,合成出两条与亲代
DNA分子完全相同的子代DNA分子的过程。
• Replicon
A unit of the genome in which DNA contain a region from origin to terminator
3’
ppp
RNA聚合酶
RNA聚合酶
合成DNA子链 3’ 5’
ppp
RNA聚合酶 DNA聚合酶 DNA聚合酶 Usain Bolt DNA聚合酶
3’
5’
切除RNA引物,填补DNA子链
5’ 3’
3’
5’
Conclusion • RNApol (RNA polymerase) [ Rif
完成对先导链引物的合成 实现DNA复制的转录激活起始
G ppp OH
A T
C G OH 3’
+
5’ ppp
G 5’ ppp
A
T
C G
ppp
OH 3’
在0.2M Nacl 的生理环境中,磷酸基团间的强电负性,使dNTP难以聚合到 DNA的5’端,而且双链DNA的5’端碱基配对困难 需要其他机制以解脱
碱基发生错配后的校正……
A
ppp T C G
A T C G
60’’
7’’
2’’
Prok. 400Nt/sec 10S (1kb)
15’’
40S
DNA replication in Okazaki fragment 1kb
3’
?
DNA replication in Okazaki fragment 1kb
3’
5’
5’
3’
(semi-discontinuous replication ! )
oriC
RNA pol 发动primer
真核生物(酵母)复制起点
AT rich
AT rich
• “呼吸现象”
DNA复制原点处氢键迅速断裂与再生,
导致两条DNA链不断解链与聚合,
形成瞬间的单泡状结构的过程。
在富含AT的区域内尤为明显
• replication origin 两侧基因的转导频率高
a c e g e c c a a a o o o o b b b b d d d f f h 复制的不同步性 断裂的随机性
Most rDNA are located
4a 3c 2e 1g 4b 3d 2f 1h
replication origin
• 复制的多模式
单起点、单方向
多起点、单方向
单起点、双方向
多起点、双方向
复制多模式的证据
1963 Cairns
慢停突变(发动突变型)
(slow-stop mutation ) 快停突变 ( fast-stop mutation) (延伸突变型) 当温度升高 当温度升高
T CA 5’ PPP C
OH
+
ppp
OH
进化中保留的 选择与适应的
化学与功能的
T CA C
5’ PPP
OH 3’
渊源
+ ppi
如果DNA的延伸方向是 3’ → 5’
G ppp OH
A T
C G OH 3’
+
5’ ppp
G A 5’ ppp
T
C G OH 3’
游离dNTP具有ppp
因能量的需要, DNA的5’端必须带有PPP
后随链按 Okazaki 片段不连续复制
3.4. DNA复制模式
(DNA replication model)
• primer
M13+
• Rifampin 是 E.coli RNA polymerase 的抑制剂!?
RF
S.S. DNA virus
+
M13
E.coli [Rif s ]
DNA polymerase不能发 无M13 RF 动子链 DNA 的复制起始 ! E.coli
5’
3’
5’
At least one strand of DNA replication in Okazaki fragment 1kb
b) 半不连续复制
(semi-discontinuous replication) 证据
• dump fragment in DNA
dUTP : dTTP = 1 :300 in cell
labeled the intact primers on the Okazaki fragments with [32p]GTP. DNA/RNA primer
destroyed DNA with DNase, leaving only the labeled primers.
genotype
RNaseH DNase
3.1. 基 本 概 念 ( Basic Concept )
What is the gene? Basic property 传宗接代 Replication
种瓜得瓜 Expression
大千世界 Mutation
DNA Double Helix model
1953 J .Watson and F. Crick
Rifampin
Rifampin
[Rif S ] + M13
E.coli [ Rif R ]
Rifampin M13
有M13 RF
有M13 RF
Conclusion
• M13 RF 的形成需要 RNA polymerase 发动 合成一段RNA分子作为 primer • RF启动后,RNA引物已经形成,Rifampin 的 抑制无效
S
]
• dnaG (primase) [Rif
R
]
完成对后随链引物的合成
较先导链的启动落后一个Okazaki片断 • 完成 10 ± Nt RNA引物合成后. DNApolII 进行 DNA 链的延伸 • DNApol I 对RNA引物切除并聚合填补
U can’t be in DNA ! dut gene
少数dUTP
A U ---A------T----
dUTPase
----G -------C---G U
突变频率 =
1/1200 !?
---A------T----
---A-----U--尿嘧啶-N-糖基 酶
---A----- --Ungase U ung
RNA polymerase [ Rif S ]
origin
10 Nt RNA பைடு நூலகம்rimer for leading Strand
dnaG
primase [ Rif
R
] for lagging Strand
Carter Nesta
DNA复制的启动与延伸
合成RNA引物
多种酶的接力赛跑
DNA模板链 5’
13mer RNA primer labeled with [32p]GTP
a and e; b and f ; c and g; d and h;
+ +
+ +
●
重新起始方式(de novo initiation) 或 复制叉式(replication fork)
starting point
→ RNA primer → transcription activation
复制的准确性 (修复,校正) 研究试材的特殊性 (温度敏感型ts,突变抑制体系Su) DNA复制速度 (E.coli 105 bp/min ,高速解旋 112 km/h ?)
缺乏统一的模式 (D.S. DNA, S.S. DNA, Linear DNA….)
3.2. 复制起点与方向
(replication origin & direction )
A U denature
A U
dump fragment ~ 1200base ~ Okazaki fragment
●
ung – dut –
dump fragment longer dump fragment shorter
●
leading strand (in dUMP F.) lagging strand (in O. F.) 直接证据?
pulse-labeling in dT-H3
stop in KCN 0℃
transfer to dT then continue D.S. DNA S.S. DNA
Density gradient of sucrose Measure H3-T
H3-T
pulse-labeling