被子植物双受精过程
被子植物的双受精过程及其生物学意义

被子植物的双受精过程及其生物学
意义
被子植物的双受精过程是指花药中的两个生殖细胞(即花粉管核和胚乳)在花内相互受精的过程。
在双受精过程中,先有一个花粉管核细胞受精,然后另一个胚乳细胞受精,最后形成一个双受精的雄性三倍体细胞。
该双受精过程在被子植物中十分重要,因为它有助于保持种群遗传多样性,并防止基因突变的出现以及对环境适应性的改变。
双受精过程使得植物能够有效地提高繁殖效率,有助于植物的繁衍、发展和适应环境变化。
双受精还可以防止基因过度流失或突变,保护植物的基因多样性,提高植物的生存能力和繁殖能力,有利于植物的长期发展。
植物学乙及完整答案

2013年植物学乙试题一、问答题:1.试述双受精的过程及生物学意义。
花粉管进入胚囊时,先端破裂,两个精子由花粉管进入胚囊。
其中一个精于与卵细胞结合,形成二倍体的合子,将来发育成胚;另一个精子与极核结合形成三倍体的初生胚乳核,这种两个精于分别与卵和极核结合的现象,称为双受精,双受精是进化过程中被子植物所特有的现象。
双受精的意义:受精作用实质上是雌、雄配子的相互同化过程,通过单倍体的雌配子一卵细胞与单倍体的雄配子一精子的结合,形成了一个二倍体的合子,由合子发育成新一代的植物体,恢复了各种植物体原有的染色体数目,保持了物种的稳定性:并且由于雌,雄配子间存在遗传差异,精、卵融合将父母本具有差异的遗传物质组合在一起,通过受精形成的合子及由它发育形成的新个体具有父母本的遗传特性,同时具有较强的生活力和适应性。
由于雌、雄配子本身相互之间的遗传差异(由减数分裂过程中所发生的遗传基因交换、重组所决定的),因而在所形成的后代中就可能形成一些新的变异,极大地丰富了后代的遗传性和变异性,为生物进化提供了选择的可能性和必然性。
双受精作用具有特殊的生物学意义。
因为双受精不仅使合子或由合子发育成的胚具有父母双方丰富的遗传特性,而且作为胚发育中的营养来源的三倍体胚乳,也是通过受精而来的,因而也带有父母双方的遗传特性。
这就使产生的后代具有更深的父母遗传特性,以及更强的生活力和适应性。
因此,被子植物的双受精,是植物界有性生殖过程中最进化最高级的形式,加上其它各种形态构造上的进化适应,使它们成为地球上适应性最强、构造最完善、种类最多、分布最广、在植物界中占绝对优势的类群。
2.列表说明花药的发育及花粉粒形成过程。
3.列表说明胚珠发育及胚囊形成过程4.指出高等植物各门首先出现的进化特征及其意义所在。
苔藓:明显的世代交替现象。
配子体占优势;孢子体不发达并“寄生”在配子体上,不能独立生活。
蕨类:明显的世代交替现象。
孢子体远比配子体(原叶体)发达,孢子体和配子体都能独立生活。
被子植物的双受精作用课件

实验结果与分析
1 2
结果展示
通过显微镜观察并记录实验结果,包括花粉发育 情况、雌蕊的受精情况等。可以使用图片和图表 等形式展示结果。
数据分析
对实验数据进行统计分析,比较不同处理组之间 的差异,并使用表格和图表等形式呈现数据。
3
结果解读
根据实验结果,分析双受精作用的机制和影响因 素,并探讨其对被子植物繁殖的意义。
胚乳提供营养
极核与精核结合形成的胚乳,为胚胎 发育提供营养物质。
03 精子与卵细胞的特点
精子的特点
精子体积较小,通常 只有卵细胞的1/20, 但数量众多。
精子头部含有细胞核 ,携带一半的遗传物 质。
精子具有鞭毛,能够 自由游动,寻找合适 的卵细胞。
卵细胞的特点
卵细胞是单倍体细胞,体积较 大,是精子体积的几十倍。
。
对植物多样性的意义
பைடு நூலகம்
增加植物种类多样性
双受精作用使得被子植物在遗传、生理和形态等方面产生了多样 性,形成了丰富的植物种类。
促进生态系统的稳定性
植物多样性的增加有助于提高生态系统的稳定性,维护生态平衡, 确保生物圈的正常运转。
提供生态服务
植物多样性在提供食物、栖息地、调节气候等方面发挥着重要作用 ,对人类和其他生物的生存和发展具有重要意义。
被子植物的双受精作 用课件
目录
CONTENTS
• 双受精作用概述 • 双受精过程 • 精子与卵细胞的特点 • 双受精作用的生物学意义 • 双受精作用的实验研究
01 双受精作用概述
定义与特点
定义
双受精作用是指被子植物的雄配 子(花粉中的精子)与雌配子( 胚囊中的卵细胞和极核)融合形 成合子的过程。
被子植物的双受精

二、被子植物的双受精花粉管里的两个精子分别与胚囊里的卵细胞和极核融合,形成受精卵和受精极核的过程,叫做双受精。
双受精是被子植物特有的受精方式。
经过开花、传粉,花粉落到雌蕊柱头上,在柱头分泌物的刺激下,花粉即开始萌发。
这时,花粉粒的内壁自萌发孔突出并逐渐伸长,形成花粉管。
花粉管穿入柱头,沿花柱向胚珠方向生长。
在生长过程中,花粉粒内的物质也全部移入花粉管顶端。
有3个细胞的花粉粒(1个营养细胞和2个精子)都进入花粉管,有2个细胞的花粉粒,生殖细胞进入花粉管后,立即分裂成两个精子。
花粉管通过花柱,到达胚珠以后,从珠孔伸进珠心的胚囊,这时,花粉管的前端破裂,两个精子移动出来,一个精子与卵细胞融合为受精卵(2n);一个精子与极核(2n)融合为受精极核(3n),完成双受精,进入新个体发育阶段。
(大孢子母细胞)减数分裂有丝分裂雌蕊:胚珠中胚囊母细胞(2N)————→大孢子—————→胚囊(N)[卵细胞(1个)、助细胞(2个)(退化消失)、反足细胞(3个)(退化消失)、极核(2个)]珠被(2N)————————————————→种皮(2N)胚珠→卵细胞(N)+精子(N)—→受精卵(2N)————→胚(2N)极核(N,2个)+精子(N)—→受精极核(3N)—→胚乳(3N)请解释:1.大孢子分裂成胚囊为何要经历3次有丝分裂?2.何为助细胞与反足细胞?3.被子植物双受精的意义。
1。
大孢子,全称是功能大孢子(更准确的说法应该是单核胚囊)经过3次有丝分裂形成八核胚囊,即有八个细胞:卵细胞(1个)、助细胞(2个)反足细胞(3个)、极核(2个)3。
一方面,通过单倍体的精子和卵子的结合,形成了一个二倍体的合子,由合子发育成新一代的植物体,恢复了各种植物原有的染色体数目,保持了物种的稳定性;并且将父母本具有差异的遗传物质组合在一起,形成具有双重遗传性的合子,即加强了后代个体的生活力和适应性,为合子发育为新一代植株可能出现某些新性状,新变异提供了基础;另一方面,由受精的中央细胞发育成三倍体的胚乳,同样兼有父母本的遗传性,作为新一代的植物胚期的养料,可以使子代的生活力更强,适应性更广。
植物的受精过程(9)

单位名称
沙雅县第二中学
填写时间
2021年7月25日
学科
生物
年级/册
七年级〔上〕
教材版本
人教版
课题名称
植物的受精过程
难点名称
子房的结构及植物的受精过程
难点分析
从知识角度分析为什么难
在花的结构中,雄蕊的花粉粒可以产生精子;雌蕊的子房内有胚珠,卵细胞在胚珠内形成。通过开花与传粉,完成受精。被子植物的受精过程是:一个精子与卵细胞结合形成受精卵,另一个精子与极核结合形成受精极核,受精卵继续发育成胚,受精极核发育成为胚乳。这就是绿色开花植物特有的双受精现象。被子植物双受精的方式比拟特殊,对初中生来说比拟难理解,不容易掌握。
6、受精过程:
花粉落到柱头上以后,在柱头上黏液的刺激下开始萌发,长出花粉管。花粉管穿过花柱,进入子房,一直到达胚珠。花粉管中的精子随着花粉管的伸长而向下移动,最终进入胚珠内部。胚珠里面有卵细胞,它跟来自花粉管的精子结合,形成受精卵。
课堂练习
〔难点稳固〕
1.一朵花中,能产生精子和卵细胞的结构分别是〔 〕
知识讲解
〔难点突破〕
一、传பைடு நூலகம்:
定义: 花粉从花药落到雌蕊柱头上的过程,叫做传粉。
介绍传粉的方式
方式:1、自花传粉:一朵花的雄蕊产生的花粉粒,落在同一朵花雌蕊的柱头上。
2、异花传粉:一朵花的雄蕊产生的花粉粒,落在另一朵花雌蕊的柱头上。
二、受精:
1、花粉粒吸水后,从萌发孔伸出花粉管,并产生两个精子,这是花粉粒发育的标志
从学生角度分析为什么难
植物的受精过程学生很难观察,切初一抽象逻辑思维较弱,理解困难,很难想象植物的受精过程。
难点教学方法
通过动态模拟图让学生直观的看到植物的受精过程
被子植物的有性生殖过程和胚、胚乳发育过程简图
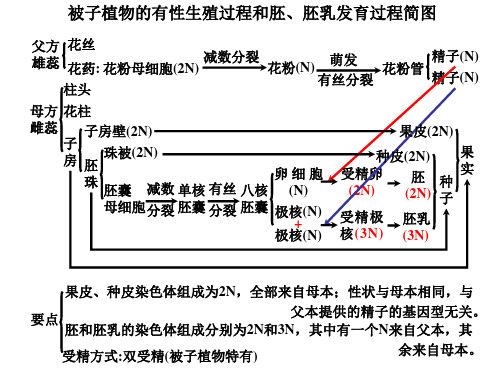
果皮、种皮染色体组成为2N,全部来自母本;性状与母本相同,与 父本提供的精子的基因型无关。 要点 胚和胚乳的染色体组成分别为2N和3N,其中有一个N来自父本,其 余来自母本。 受精方式:双受精(被子植物特有)
被子植物的有性生殖过程和胚、胚乳发育过程简图
父方 花丝 精子(N) 雄蕊 花药: 花粉母细胞(2N) 减数分裂 花粉(N) 萌发 花粉管 精子(N) 有丝分裂 柱头
母方 花柱 雌蕊 子房壁(2N) 子 珠被(2N) 房 胚 珠
果皮(2N) 果 种皮(2N) 实 受精卵 胚 种 (2N) (2N) 子 受精极 胚乳 核 (3N) (3N)
被子植物的双受精作用

双受精作用在植物进化过程中发挥了重要作用,通过遗传物质的重 组和变异,推动了植物的适应性和多样性的发展。
受精过程的生物学意义
有利于扩大植物的地理分布范围
双受精作用产生的胚乳富含营养物质,为植物种子的萌发和早期生长提供了充足的养分, 有利于植物在各种环境中的适应性生长。
有利于植物的繁殖和种群增长
精细胞在花粉管内的运动
精细胞在花粉管内通过鞭毛运动向卵细胞移动。
识别与结合
精细胞识别雌性生殖细胞的特异性受体,与卵细胞结合形成合子。
雄性生殖细胞的发育与分化
减数分裂
雄性生殖细胞经历减数分裂,产生四 个精细胞。
细胞壁的形成
精细胞形成细胞壁,准备进入花粉管 。
雌性生殖细胞的发育与分化
卵细胞的形成
雌性生殖细胞经过多次分裂形成卵细胞。
特点
双受精作用是被子植物有性生殖过程 中特有的现象,其结果是形成受精卵 和受精极核,分别发育成胚和胚乳。
受精过程的重要性
遗传物质的传递
双受精作用实现了遗传物质的横向传递,使后代具有父母本的遗 传特性,从而保持了物种的遗传稳定性。
植物体的发育
受精卵和受精极核分别发育成胚和胚乳,为植物体的发育提供了必 要的物质基础,保证了植物体的正常生长和发育。
被子植物的基因组通常由多个染色体组成,这些染色体携带 了植物生长、发育和繁殖所需的遗传信息。
染色体
染色体是基因组的载体,由DNA和蛋白质组成,它们以特定 的方式排列和组织,以确保遗传信息的正确传递。
基因重组与遗传信息的传递
基因重组
在双受精过程中,雌雄配子通过融合形成合子,导致基因重组的发生。基因重组有助于增加基因组的多样性和复 杂性,促进植物的适应性和进化。
植物学第八章第四节开花、传粉、受精

异花传粉
Lloyd & Webb认为,雌雄异熟 的类型很多,按两性功能出现的时 间顺序分为雌性先熟,如油菜、甜 菜的两性花为雌蕊先熟;和雄性先 熟、向日葵、梨、苹果的两性花为 雄蕊先熟。按雌雄异熟表现在花内 或花间分为花内雌雄异熟和花间雌 雄异熟,如玉米的雄花序比雌花序 先成熟。根据雌雄异熟的分离程度 分为完全雌雄异熟、不完全雌雄异 熟。按照株内雌雄成熟的同步性分 为同步雌雄异熟),可见于胡萝卜、 花蔺 等植物中;以及异步雌雄异熟, 发现于核桃科、蔾科、瑞香科、桦 木科和榛属等植物中。
一般可分为2类: ①配子体型自交不亲和性 即花粉在柱头上萌发后可侵入柱头,并能在花柱组织中延伸一段,此后就受到抑制。花粉管与雌性因 素的抑制关系发生在单倍体配子体(即卵细胞与精细胞)之间。常见于豆科、茄科和禾本科的一些植物。 这种抑制关系的发生可以在花柱组织内,也可以在花粉管与胚囊组织之间;有的甚至是花粉管释放的精子 已达胚囊内,但仍不能与卵细胞结合。
②孢子体型自交不亲和性 即花粉落在柱头上不能正常发芽,或发芽后在柱头乳突细胞上缠绕而无法侵入柱头。由于这种不亲和 关系发生在花粉管与柱头乳突细胞的孢子体之间,花粉的行为决定于二倍体亲本的基因型,因而称为孢子 体型自交不亲和性,多见于十字花科和菊科植物。
自花不孕 自花不孕是指花粉粒落 到同一朵花或同一植株的柱 头上不能结实的现象。自花 不孕有两种情况:一种是花 粉粒落到自花的柱头上,根 本不能萌发,如向日葵、荞 麦、黑麦等;另一种是自花 的花粉粒虽能萌发,但花粉 管生长缓慢,一般没有异花 的花粉管生长快,最终不能 自体受精,如玉米、番茄等 (进行玉米等自交系的培育, 必须在人工传粉后套袋隔 离)。此外,某些兰科植物 的花粉对自花的柱头有毒害 作用,常引起柱头凋萎,以 致花粉管不能生长。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
二、卵和合子
卵细胞核对孚尔根反应与精子不同, 刚形成的卵核呈孚尔根正反应, 成熟的卵核则是负反应或弱的正反应。
胡适宜和朱至清对几种植物的卵细 胞和中央细胞核研究的结果,证明不 同植物有差异
棉花、向日葵和扁豆
年幼的卵细胞核和胚囊中央细胞核 都表现强的孚尔根反应,
随着发育而逐渐降低对孚尔根反应 的强度。
二、糖蛋白在配子识别中的作用
1 精、卵融合前期有关的糖蛋白
被子植物的受精识别从广义而言可看作是 多位点的。首先是从花粉与柱头表面的接 触开始。对于自交不亲和性植物,其等位 基因编码一系列S位点糖蛋白。通过这些S 糖蛋白的作用,植物最终表现为自交不亲 和性状。对于同种亲和性植物,其花粉与 柱头之间也存在识别,并与花粉壁、柱头 表面及花柱引导组织内的糖蛋白有关。
在柱头的乳突(papillae)的表 面,存在一层亲水蛋白质的薄膜。 一方面有粘着花粉或水合花粉的 作用,同时也是不亲和系统的识 别反应的位置。
烟草柱头扫描电镜照片
湿型柱头:柱头在传粉时有溢出物, 它为花粉萌发提供必须的基质。溢出 物含少量游离的糖,还含脂类和酚的 化合物。
干型柱头:柱头在传粉时是不产生分 泌物的。这种柱头由于在表面存在蛋 白质薄膜的亲水性,能通过其下层的 角质层的中断处汲水,辅助粘着花粉 和使花粉获得萌发必须的水分。
花粉萌发 花粉落在柱质相互作 用。
二者如果是亲和的,产生“认可” 反应,花粉粒由周围吸收水分,内 壁由萌发孔伸出为萌发管。
二、花粉管在雌蕊组织中生长
1 花粉管进入柱头和花柱
从柱头组织的细胞壁之间或通过细胞壁 向下生长的
开放型花柱:有中空的管道,管道内
与中央细胞融合的精子细胞质参与融合。
大麦
3、与卵及与中央细胞融合的精子细胞质都 参与融合。矮牵牛
三 、配子质膜的融合
精细胞被释放在卵与中央细胞之间的区域。 包围它们的花粉管内质膜崩解,两个裸露
的精细胞同时分别与卵和中央细胞接触。 精细胞与雌性细胞质膜间的距离从20~ 70μm不等。 膜的融合开始在膜的一个位点,然后在多 个位点融合,融合的膜逐渐崩解,直至仅 观察到纤细的膜轮廓。
表皮细胞能分泌粘性物质,花粉管沿着 管道的内表面,在粘液中向下生长。
闭合型花柱 :没有中空的花柱道, 有引导组织 。细胞内含丰富的细
胞器,具较高的活性,花粉管在 充满基质的间隙中生长。
花粉管在花柱中生长
花粉管生长部位:
花粉管尖端5μm左右有生长点,在光 学显微镜下呈较透明的半球形,其中 含有高尔基体、核糖体等多种细胞器, 丰富的RNA,各种酶类和多糖、油滴 等。这些特征与形成新的管壁有关。
花粉管生长的营养来源:
(1)花粉本身的贮藏物 (2)柱头和花柱组织中
2 花粉管到达胚珠和进入胚囊
花粉管穿过珠孔后,径直朝向助细胞 丝状器从而进入胚囊中。花粉管穿入 两个助细胞中的一个后,于花粉管的 顶端形成一个孔,从孔中释放出花粉 管的内容物。
精细胞、一个营养核和少量细胞质
三、双受精
精子*卵———合子———胚 精子*两个极核—初生胚乳核—胚乳
2 、 精细胞表膜的糖分子和糖蛋白
植物精细胞表膜存在多种糖蛋白,其 中可能包括某些凝集素受体。
第四节 精子、卵及合子中DNA的含量
植物受精作用是植物世代交替的转折点, 它涉及配子体世代向孢子体世代的转变, 并伴随发生DNA倍性从单倍转变为二倍。
一、精子
精子核均呈孚尔根正反应,DNA特异性 荧光染料染色也均表现强的荧光。
第九章 受 精
第一节 双受精过程
受精即雌、雄配子的融合
配子配合前期(progamic phase) 配子配合(syngamy)
一、花粉萌发
花粉被传送到雌蕊的柱头上, 柱头具有特殊的表面,适于“捕捉” 和承受花粉,并为花粉萌发时提供 必须的水分。
柱头通常有大的表面,表皮细 胞变为乳突状或毛状,适于附着花 粉。
配子融合的时期十分短促,在白花丹 似乎整个时期历时不到1 min,推测游
离钙可能对促进膜的融合起一定作用。
第三节 配子识别
细胞间的识别(recognition)是生物 界普遍存在的现象之一。它是细胞分 辨“自己”与“异己”的一种能力, 是细胞表面在分子水平上选择性的相 互识别与辨别。
配子融合时两个精细胞与卵和中央细 胞的结合是随机的,还是预定性的呢?
1、在配子融合时,两个精子的细胞质 都不参与融合。陆地棉
在这种融合模式中尚有一个未能阐明的 问题,即核从精细胞独自转移的完成,精 细胞必须维持凝胶状态,阻止精细胞质流 动至卵细胞及接着发生细胞质的闭合。然 而,既使在受精时每分钟采样的详细观察 也未得到显示膜闭合的证据。
2、与卵融合的精子的细胞质被排斥,而
被子植物的双受精
A.花粉管进入胚囊 B.花粉管释放出内含物 C.两个 精子分别转移至卵和中央细胞 D.精核转移示意图a.精 子与卵接近,b.精核在转移中,c.精核转移到卵细胞中
第二节 配子的融合
花粉管进入胚囊是经由退化的一个助细胞 一般认为细胞内高水平的游离钙或松弛的 结合钙,是导致助细胞退化的主要因素。 另一方面,花粉管成分、渗透压或pH的改 变,所有这些因素构成的微环境,对促进 花粉管的释放和停止生长起作用。
两个精细胞在花粉管中下行时,它们 一直维持连结。
雌性生殖单位结构特点,阐明了在受 精的靶区缺少细胞壁,从而认识到雌、 雄配子之间能通过膜的融合而为为一 体。
二、配子融合时雄配子细胞质的参与
虽然能确定融合前的精子是具细胞质 的,但对它们与雌性配子融合时,细 胞质是否也同核一起进入雌配子,至 今未有一致的结论。
一、被子植物配子识别的假说
1、随机假说(chance hypothesis) 精细胞与卵细胞或中央细胞的结合是
随机决定的。 2、特异受体假说(specific receptor
hypothesis) 雌雄配了的融合是由其细胞表面特异
识别因子决定的。
Faure (1999) 提出三种模型 :
精-卵识别的三种模型