近亲繁殖和杂种优势
合集下载
近亲繁殖和杂种优势—杂种优势(普通遗传学课件)

** 1 13
掖单19 15.5
** 1 14
豫玉23 15.2 各省 对照种 15
** 1
8
** 1 13
二、突出优点
(二)稳产 据国家、省区试结果,地点、年份间变异系数最
小,属高产稳产类型。
二、突出优点
(三)抗病虫能力强
据国家指定单位鉴定:兼抗7种主要病虫害,是国
内为数不多的多抗品种。
郑单958对几种主要病(虫)害的接种鉴定结果(1999年)
说明杂种基因型的高度杂合性是形成杂种优势的 根源。
× 优势高:产量高,品质好
→
三、优势大小与双亲基因型的纯合程度密切相 关
杂交亲本越纯,杂种优势越明显。 杂种优势一般体现在群体的优势表现。双亲基因 型纯度高→F1群体具有整齐一致的异质性→优势明显。 如:一般玉米杂种优势表现为: 单交种> 双交种> 品种间杂交种
2023/9/3
一、衰退现象的含义
衰退现象:F2与F1相比较,生长势、生活力、抗逆性 和产量等方面都显著地表现下降的现象。
原因:F2群体中由于基因的分离和重组,必然会出 现性状的分离和重组。
小麦F2群体
玉米杂种优势随自交代数的增加而不断下降 (Poehlman,1优势的计算方法
(二)超亲优势 超亲优势:F1超过最优亲本的百分率。
H
F1-HP HP
100%
分子部分即杂种均值与纯种亲本均值的差值,称 为杂种优势量,若>0为正杂种优势,﹤0为负杂种优 势。
三、杂种优势的计算方法
(三)对照优势 对照优势:F1超过生产对照品种的百分率。
H
F1-CK CK
《遗传学》
杂种优势利用的发展简史与现状
一、杂种优势利用的发展简史
第九章-近亲繁殖与杂种优势

A1A1<A1A2>A2A2 杂合座位越多优势越强
又称:等位基因互作说 杂合性假说
A1B1C1 D1E1 × A2B2C2 D2E2 A1B1C1 D1E1 A2B2C2 D2E2
纯合状态 1个单位 杂合状态 2个单位
F1 A1B1C1 D1E1 A2B2C2 D2E2
2+2+2+2+2=10
根据这一假说,等位基因间没有显隐性关系。 杂合等位基因间的相互作用显然大于纯合等位 基因间的相互作用
3基因杂合 2杂1纯
1杂2纯 3基因纯合 (8 种)
四. 回交的遗传效应
回交(back cross)是指杂交后代与两亲本之一的再次交配。
甲×乙
↓ F1×乙
↓ 回交一代BC1×乙
↓ 回交二代BC2×乙
↓ 回交三代BC3×乙
↓
···
轮回亲本: 用来连续回交的亲本。
1. 基因代换
2.
连续回交,核内基因逐渐被轮回亲本取代。
(需年年配制杂种,较为费时费力) ■ 杂种优势的固定
利用杂种优势时,注意点:
■ 保持亲本的纯合性和典型性; ■ 选择自身表现好,配合力高的优良亲本及
其强优势组合; ■ 掌握杂交制种和亲本保种技术。
上位性是杂种优势的主要遗传基础
Proposed by R.A.Fisher in 1949, K. Mather in 1955, R.W.Allard in 1962, S.B.Yu in 1997.
华金平,张启发 显性、上位性、超显性在杂种优势中共同存在。 PNAS,Genetice(2002)
F2及其以后的世代 纯合体 多基因假说
近亲繁殖在育种上的应用
■ 保持品种纯度和相对的稳定(纯系) ■ 作物常采用自交、回交或兄妹交 (因授粉方式和育种方法不同而不同)
又称:等位基因互作说 杂合性假说
A1B1C1 D1E1 × A2B2C2 D2E2 A1B1C1 D1E1 A2B2C2 D2E2
纯合状态 1个单位 杂合状态 2个单位
F1 A1B1C1 D1E1 A2B2C2 D2E2
2+2+2+2+2=10
根据这一假说,等位基因间没有显隐性关系。 杂合等位基因间的相互作用显然大于纯合等位 基因间的相互作用
3基因杂合 2杂1纯
1杂2纯 3基因纯合 (8 种)
四. 回交的遗传效应
回交(back cross)是指杂交后代与两亲本之一的再次交配。
甲×乙
↓ F1×乙
↓ 回交一代BC1×乙
↓ 回交二代BC2×乙
↓ 回交三代BC3×乙
↓
···
轮回亲本: 用来连续回交的亲本。
1. 基因代换
2.
连续回交,核内基因逐渐被轮回亲本取代。
(需年年配制杂种,较为费时费力) ■ 杂种优势的固定
利用杂种优势时,注意点:
■ 保持亲本的纯合性和典型性; ■ 选择自身表现好,配合力高的优良亲本及
其强优势组合; ■ 掌握杂交制种和亲本保种技术。
上位性是杂种优势的主要遗传基础
Proposed by R.A.Fisher in 1949, K. Mather in 1955, R.W.Allard in 1962, S.B.Yu in 1997.
华金平,张启发 显性、上位性、超显性在杂种优势中共同存在。 PNAS,Genetice(2002)
F2及其以后的世代 纯合体 多基因假说
近亲繁殖在育种上的应用
■ 保持品种纯度和相对的稳定(纯系) ■ 作物常采用自交、回交或兄妹交 (因授粉方式和育种方法不同而不同)
14 近亲繁殖和杂种优势

❖ 纯系学说的基本内容:在自花授粉植物的天然混 杂群中,可分离出许多基因型纯合的纯系,因此 在一个混杂群体中选择是有效的;但是,在纯系 内个体所表现的差异只是环境的影响,是不能遗 传的,所以在纯系内选择是无效的。
❖ 注意:纯系是相对的而不是绝对的。
2020/4/24
14.3 近交与杂交的遗传效应
❖ 近交增加纯合子频率,杂交增加杂合子频率。 ❖ 近交降低群体均值,杂交提高群体均值。 ❖ 近交使群体分化,杂交使群体一致。 ❖ 近交加选择能加大群体间基因频率的差异,从而
❖ 杂种优势的表示方法HF1(%)HPF1 100%
用绝对值表示:即F1超过其双亲平均数的部分
HF1 PF1 P
用相对值表示:即F1超过双亲平均数的百分率
HF1(%)HPF1 100%
2020/4/24
14.4.2 杂种优势的表现
按性状表现的性质分类:营养型、生殖型、适应型 杂种优势表现的特点:
Chapter14 近亲繁殖与杂种优势
基本要求 14.1 概念 14.2 纯系学说 14.3 近交与杂交的遗传效应 14.4 杂种优势 14.5 近交与杂交在育种上的应用 复习思考题
2020/4/24
基本要求
➢掌握近交与杂交的基本概念及近交与杂交的遗传效应 ➢了解近交衰退现象及防止近交衰退的方法 ➢了解杂种优势理论及杂种优势表现的特点 ➢了解近交与杂交在育种上的用途
❖ 杂交(cross breeding):指两个或更多个品种之 间、或同一品种的不同品系之间个体的交配;就某 一性状而言,指不同纯合子间的异型交配。
2020/4/24
14.2 纯系学说(pure line theory)
❖ 纯系(pure line)是指从一个基因型纯合个体 自交产生的后代群体,其基因型也是纯一的。
❖ 注意:纯系是相对的而不是绝对的。
2020/4/24
14.3 近交与杂交的遗传效应
❖ 近交增加纯合子频率,杂交增加杂合子频率。 ❖ 近交降低群体均值,杂交提高群体均值。 ❖ 近交使群体分化,杂交使群体一致。 ❖ 近交加选择能加大群体间基因频率的差异,从而
❖ 杂种优势的表示方法HF1(%)HPF1 100%
用绝对值表示:即F1超过其双亲平均数的部分
HF1 PF1 P
用相对值表示:即F1超过双亲平均数的百分率
HF1(%)HPF1 100%
2020/4/24
14.4.2 杂种优势的表现
按性状表现的性质分类:营养型、生殖型、适应型 杂种优势表现的特点:
Chapter14 近亲繁殖与杂种优势
基本要求 14.1 概念 14.2 纯系学说 14.3 近交与杂交的遗传效应 14.4 杂种优势 14.5 近交与杂交在育种上的应用 复习思考题
2020/4/24
基本要求
➢掌握近交与杂交的基本概念及近交与杂交的遗传效应 ➢了解近交衰退现象及防止近交衰退的方法 ➢了解杂种优势理论及杂种优势表现的特点 ➢了解近交与杂交在育种上的用途
❖ 杂交(cross breeding):指两个或更多个品种之 间、或同一品种的不同品系之间个体的交配;就某 一性状而言,指不同纯合子间的异型交配。
2020/4/24
14.2 纯系学说(pure line theory)
❖ 纯系(pure line)是指从一个基因型纯合个体 自交产生的后代群体,其基因型也是纯一的。
第9章近亲繁殖和杂种优势

纯合体在遗传上是稳定的,杂合体在遗传上是 不稳定的。
自交使得基因型不断得到纯合,从而使遗传性 状的表现稳定一致。
自交和近交对于农作物品种的保纯和物种的稳 定性都具有重要的意义。
四、回交及其遗传效应 回交在遗传学研究和育种上都是常用的方法。
利用隐性亲本和F1回交,可以测定F1产生的 配子的种类和比例。
近交和杂交的比较:
近交可以使原本是杂交繁殖的生物增加纯合性 (homozygosity),提高遗传稳定性,但是 往往伴随严重的近交衰退现象(inbreeding depression)。
杂交(outbreeding),可以使原本自交或 近交的生物增加杂合性(heterozygosity), 产生杂种优势(heterosis)。
回交与自交的区别 ❖回交和自交都能导致后代基因型的纯合 ❖自交后代中纯合基因型的种类多种多样 ❖而回交后代中基因型严格受轮回亲本的制约
回交与自交的区别
❖ 假设3对基因,自交后代应该有23=8种纯合基因型 AABBCC AABBcc AAbbCC AAbbcc aaBBCC aaBBcc aabbCC aabbcc
同一基因座上的两个等位基因,如果分别来自没有亲缘 关系的两个祖先,即使功能上是相同的,核苷酸序列却 不一定完全相同。 只有同一祖先某一基因座上的某个等位基因的拷贝,其 核苷酸序列才是完全相同的。
❖两个有亲缘关系的个体有可能从共同祖先 那里得到同一个等位基因。
❖这两个个体交配,又有可能把同一个等位 基因传递给同一个子女。
❖ 杂合体自交子代中,纯合体的两个等位基 因的核苷酸序列是完全相同的,其比例是 1/2,也就是近交系数等于1/2。
近交系数的计算:
➢ ♀ A1A2、♂A3A4
➢ 子代的基因型为
自交使得基因型不断得到纯合,从而使遗传性 状的表现稳定一致。
自交和近交对于农作物品种的保纯和物种的稳 定性都具有重要的意义。
四、回交及其遗传效应 回交在遗传学研究和育种上都是常用的方法。
利用隐性亲本和F1回交,可以测定F1产生的 配子的种类和比例。
近交和杂交的比较:
近交可以使原本是杂交繁殖的生物增加纯合性 (homozygosity),提高遗传稳定性,但是 往往伴随严重的近交衰退现象(inbreeding depression)。
杂交(outbreeding),可以使原本自交或 近交的生物增加杂合性(heterozygosity), 产生杂种优势(heterosis)。
回交与自交的区别 ❖回交和自交都能导致后代基因型的纯合 ❖自交后代中纯合基因型的种类多种多样 ❖而回交后代中基因型严格受轮回亲本的制约
回交与自交的区别
❖ 假设3对基因,自交后代应该有23=8种纯合基因型 AABBCC AABBcc AAbbCC AAbbcc aaBBCC aaBBcc aabbCC aabbcc
同一基因座上的两个等位基因,如果分别来自没有亲缘 关系的两个祖先,即使功能上是相同的,核苷酸序列却 不一定完全相同。 只有同一祖先某一基因座上的某个等位基因的拷贝,其 核苷酸序列才是完全相同的。
❖两个有亲缘关系的个体有可能从共同祖先 那里得到同一个等位基因。
❖这两个个体交配,又有可能把同一个等位 基因传递给同一个子女。
❖ 杂合体自交子代中,纯合体的两个等位基 因的核苷酸序列是完全相同的,其比例是 1/2,也就是近交系数等于1/2。
近交系数的计算:
➢ ♀ A1A2、♂A3A4
➢ 子代的基因型为
第八章 近亲繁殖与杂种优势

血缘系数为0:个体 和 间近期世代内没有共同祖先 血缘系数为 :个体x和y间近期世代内没有共同祖先
(2)近亲系数和血缘系数的计算 )
通经( 通经(path): 在一个相关的网络系统中,连接原因变量和 结果变量 的 ) 在一个相关的网络系统中, 每一条箭头为一个通经 每一条箭头为一个通经 通经系数( 通经系数(path coefficient) :原因变量对结果变量的影响系数 ) 原因变量对结果变量的影响系数 通经分析原理:随机交配群体中,每一条通经的通经系数为 通经分析原理:随机交配群体中,每一条通经的通经系数为1/2 ·两个结果变量间的相关系数等于连接它们的所有通经的 两个结果变量间的相关系数等于连接它们的所有通经的 通经系数的乘积
基因型和表现型。 首次提出基因型和表现型 首次提出基因型和表现型。
(三)、 纯系学说的意义 三、
• 理论意义: 理论意义: – 区分了遗传变异和不遗传的变异,指出选择遗传变异 区分了遗传变异和不遗传的变异, 遗传变异 的变异 的重要性,并对遗传、环境及其与个体发育、 的重要性,并对遗传、环境及其与个体发育、性状表 现关系研究起了很大推动作用。 现关系研究起了很大推动作用。 • 实践意义: 实践意义: – 直接指导自花授粉植物的育种,即:可以在混杂群体 直接指导自花授粉植物的育种, (如地方品种群体 内进行单株选择得到不同的纯系;但 如地方品种群体)内进行单株选择得到不同的纯系 如地方品种群体 内进行单株选择得到不同的纯系; 是在纯系中继续选择是无效的。 是在纯系中继续选择是无效的。
轻粒种子 63.2 75.2 54.6 63.6 74.4 69.1 66.7
重粒种子 64.9 70.9 56.9 63.6 73.0 67.7 66.2
第五章 近亲繁殖和杂种优势(P99)
(2)近亲系数和血缘系数的计算 )
通经( 通经(path): 在一个相关的网络系统中,连接原因变量和 结果变量 的 ) 在一个相关的网络系统中, 每一条箭头为一个通经 每一条箭头为一个通经 通经系数( 通经系数(path coefficient) :原因变量对结果变量的影响系数 ) 原因变量对结果变量的影响系数 通经分析原理:随机交配群体中,每一条通经的通经系数为 通经分析原理:随机交配群体中,每一条通经的通经系数为1/2 ·两个结果变量间的相关系数等于连接它们的所有通经的 两个结果变量间的相关系数等于连接它们的所有通经的 通经系数的乘积
基因型和表现型。 首次提出基因型和表现型 首次提出基因型和表现型。
(三)、 纯系学说的意义 三、
• 理论意义: 理论意义: – 区分了遗传变异和不遗传的变异,指出选择遗传变异 区分了遗传变异和不遗传的变异, 遗传变异 的变异 的重要性,并对遗传、环境及其与个体发育、 的重要性,并对遗传、环境及其与个体发育、性状表 现关系研究起了很大推动作用。 现关系研究起了很大推动作用。 • 实践意义: 实践意义: – 直接指导自花授粉植物的育种,即:可以在混杂群体 直接指导自花授粉植物的育种, (如地方品种群体 内进行单株选择得到不同的纯系;但 如地方品种群体)内进行单株选择得到不同的纯系 如地方品种群体 内进行单株选择得到不同的纯系; 是在纯系中继续选择是无效的。 是在纯系中继续选择是无效的。
轻粒种子 63.2 75.2 54.6 63.6 74.4 69.1 66.7
重粒种子 64.9 70.9 56.9 63.6 73.0 67.7 66.2
第五章 近亲繁殖和杂种优势(P99)
07第七章 近亲繁殖和杂种优势讲解

父母与儿女之间、同胞兄弟姐妹之间基因 相同的可能性为1/2,彼此之间称为一级亲属; 祖父母与孙子(女)之间、外祖父母与外孙子(女) 之间、叔(伯、姑)与侄儿(女)之间、舅(姨)与外 甥儿(女)之间基因相同的可能性为1/4,彼此 之间称为二级亲属;堂兄弟姐妹之间、表兄弟 姐妹之间基因相同的可能性为1/8,彼此之间 称为三级亲属,依次类推。
菜豆两个株系按粒重选择和种植的结果( P339之表12-4)
小粒株系
大粒株系
收获年份 当选植株种子 后代植株种子 当选植株种子 后代植株种子
平均重(mg) 平均重(mg) 平均重(mg) 平均重(mg)
轻粒种子 重粒种子 轻粒种子 重粒种子 轻粒种子 重粒种子 轻粒种子 重粒种子
1902
300 400 358 348 600 700 632 649
举例:以一对基因为例,即AA×aa F1 100%都是杂合体 F2 有1/2杂合体和1/2纯合体(根据
分离比例而得) F3 有1/4杂合体和3/4纯合体
Fr+1有(1/2)r杂合 体,1-(1/2)r 纯合体。 详见下表 。
当连续自交多代时,后代将逐渐趋于 纯合,每自交一代,杂合体所占比例即减 少一半,并逐渐接近于 0 , 而不会完全 消失。纯合体增加的速度,则与所涉及的 基因对数及自交代数有关。 继续
1903
250 420 402 410 550 800 752 709
1904
310 430 314 326 500 870 546 569
1905
270 390 383 392 430 730 636 636
1906
300 460 379 399 460 840 744 730
1907 平均
第十二章 近亲繁殖与杂种优势
纯合率=29791/32768x100%=90.91% 杂合率=1-90.91%=9.09% 5 r 5 纯合率x=[1-1/2 ] n x100%=[(2 -1)]/2 X100%=90.91%
例:设一大小植株的基因型为AABbCcDd,已知各对基因是独立遗传的;如果 例:设一大小植株的基因型为AABbCcDd,已知各对基因是独立遗传的;如果 连续自交5 试问其后代群体中具有1对、2对、3 连续自交5代,试问其后代群体中具有1对、2对、3对基因的纯合率各是多少? r n [1+(2 -1 )] n=3 r=5
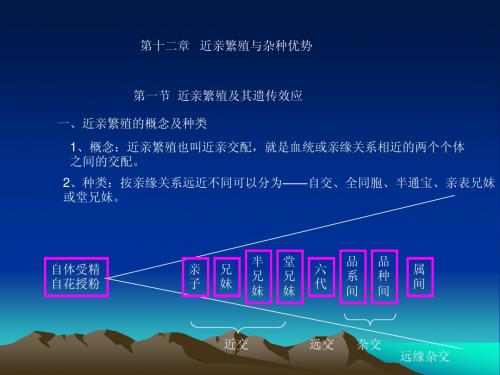
第十二章 近亲繁殖与杂种优势
第一节 近亲繁殖及其遗传效应 一、近亲繁殖的概念及种类 1、概念:近亲繁殖也叫近亲交配,就是血统或亲缘关系相近的两个个体 之间的交配。 2、种类:按亲缘关系远近不同可以分为——自交、全同胞、半通宝、亲表兄妹 或堂兄妹。
自体受精 自花授粉
亲 子
兄 妹
半 兄 妹
堂 兄 妹
六 代
③、群体总的遗传方差 σ 2 2 2 2 σ =2F σ G+(1-F) G =(1+F) σ G
群体总的遗传方差随近交系数上升而增大。
4、近交衰退(inbreeding depression) 近交衰退(inbreeding depression):指近交使生物个体的生活力、生产性能、 群体均值都降低的现象。
RsD:个体S和D之间的亲缘系数; FA: 共同祖先的近交系数; Fs: 个体S的近交系数; FD: 个体D的近交系数; N: 个体S和D分别到共同祖先的代数之和。
如果A、S、D都不是近交个体,则FA=FS=FD=0 RsD=∑(1/2) ②、亲兄妹 N
(1+1) (1+1) RsD=(1/2) +(1/2) =50%
基础六 近亲繁殖繁殖与杂种优势

模块三 异花授粉植物的自交不亲和性
异花授粉植物为了确保异花授粉,在进化过程中形成了不同的自 花授粉障碍,即自交不亲和性。 自交不亲和性:植物雌雄蕊发育正常,但自花授粉不能结实,这 种特性称为自交不亲和性。有广义和狭义之分。 广义的自交不亲和性是指形态结构上的自交不可能性。 1.雄蕊先熟 在这种情况下,雄蕊在同一朵花中的柱头成熟之前先 成熟,从而避免了花粉粒传到柱头上授粉受精,从而导致不育。 2.雌蕊先熟 在这种情况下,柱头比雄蕊成熟早,同样导致不育。 3.自花不育 在没有昆虫传粉的条件下,同一植株上的雌性和雄性 器官的自然排列阻止自花授粉。 狭义的自交不亲和性是遗传学和生理学的机制所致的自交不亲和 性。如许多种类的苹果、梨、杏、李子、甜菜、油菜等都属于此 类。
(二)杂种优势的利用 杂种优势在生产上已被广泛利用。 在作物方面,因其繁殖和授粉方式不同,利用杂种优势 的方法有所差异。 对无性繁殖作物,如甘薯、马铃薯等,只要选择那些具 有杂种优势的优异单株进行无性繁殖,就可将其杂种优 势固定。 而对于有性繁殖作物,由于杂种优势一般只能利用F1(F2 以后,杂种优势逐渐下降)。因此,必须年年制种。工作 量非常大,比如玉米制种,必须进行去雄、授粉。为了 提高制种效率,在一些作物上(如水稻)采用了三系配 套制种。就是培育雄性不育系,这就省去了去雄这一环 节。
(2+1+1+2+1=7)×(1+2+2+1+2=8)→(2+2+2+2+2=10) 用显性假说解释杂种优势示意图
(二)超显性假说
超显性假说的基本观点是:杂种优势产生的原因是 双亲基因型的异质结合所引起的等位基因间的相互作用。 等位基因间没有显隐性关系,异质的等位基因相互作用 大于同质等位基因的作用,即a1a2>alal或a2a2。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
第九章近亲繁殖和杂种优势
一、近亲繁殖及其遗传效应
1、近亲繁殖的概念
异交:亲缘关系较远的个体间随机相互交配
近交:亲缘关系相近的个体间交配按亲缘关系远近程度,可分为:
全同胞、半同胞、亲表兄妹
植物还有:自交、回交
自交是最极端的方式
自交
↓
回交(父女或母子交配)
↓
全同胞交配(同父母的兄妹交配)
↓
半同胞交配(同父或同母的兄妹交配)
2、杂种优势理论
(1)显性假说
双亲的显性基因全部聚集在杂种中产生互补作用。不能有效解释数量性状杂种优势的遗传机理
(2)超显性假说(等位基因异质结合假说)
杂种优势来源于双亲基因型的异质结合所引起的等位基因间的互作。未考虑不同基因位点之间相互作用即上位性效应
(3)上位性假说
杂种优势的遗传实质上在于数量性状基因各个遗传效应的综合作用:加性、显性、上位性、GE互作效应
适应型:杂种对外界不良环境适应能力较强
杂种优势表现特点:
(1)不是某一二个性状单独表现突出,而是许多性状综合表现突出。如产量
(2)杂种优势的大小,取决于双亲性状间的相对差异和相互补充。亲本选配
(3)杂种优势的大小与双亲基因型的高度纯合有关
单交种、双交种、品种间
(4)杂种优势大小与环境条件有关
F1表现杂种优势,F2出现衰退现象,因为F2会出现性状分离
1
(1 ---------)n
2r
因此,回交在基因型纯合进度上大于自交,一般回交5-6代即可
二、纯系学说
纯系学说:约翰生1909年提出。他以自花授粉的菜豆天然混杂群体为试验材料,按豆粒的轻重分别播种,从中选出19个单株。这19个单株的后代,即19个株系,在平均粒重上彼此具有明显的差异,而且是能够稳定遗传的。他又在19个株系中分别选择最轻和最重的两类种子分别种植,如此连续进行6年,发现在各年份里同一株系内轻粒和重粒的后代平均粒重几乎没有差异
纯系学说:
(1)在自花授粉植物的天然混杂群体中,可分离出许多基因型纯合的纯系。在一个混杂群体中选择是有效的
(2)在纯系内个体所表现的差异,那只是环境的影响,是不能遗传的。所以,在纯系内继续选择是无效的
局限性
纯系:从一个基因型纯合个体自交产生的后代(自交系)
三、杂种优势
1、杂种优势表现
杂种优势:两个遗传组成不同的亲本杂交产生的杂种F1在生长势、生活力、繁殖力、抗逆性、产量、品质等上优越于双亲的现象。杂种优势所涉及的性状大都为数量性状
(1)纯合基因型严格受轮回亲本的控制,1种纯合基因型;
自交是2n种纯合基因型
(2)纯合率同样用:
1
纯合体频率x= (1 -------)n
2r
但是,纯合率包含的内容不同
自交后代每一种基因型的纯合率为:
11
(1 ------------)n×(------)n
2r2
回交后代最后只有一种接近轮回亲本的纯合基因型,纯合率为:
F10Aa(1/2)00
↓
F211 AA 2Aa 1aa(1/2)11-(1/2)1
↓
F32 4AA 2 AA 4Aa 2aa 4aa (1/2)21-(1/2)2
. ..
Fr+1r(1/2)r1-(1/2)r
设有n对异质基因,自交r代
1
则纯合体频率x= (1 --------)n
2r
某一种纯合基因型的频率:
如果杂种的性状表现超过双亲的均值,这种优势称为中亲优势(平均优势)
F1-MP
中亲优势(%)=---------------×100%
MP
在实际育种中常使用超亲优势,即超越优亲的杂种优势
F1-HP
超亲优势(%)=----------×10旺
生殖型:生殖器官发育较盛
11
(1 -----------)n╳-----
2r2n
n对基因均杂合的个体的频率:
[1+(2r-1)]n的展开为Fr+1群体中各种个体数的分布
如:n=3 r=5
[1+(25-1)]3=1+93+2883+29791
(2)杂合体通过自交导致等位基因纯合,淘汰有害隐性纯合体
(3)杂合体通过自交,获取不同纯合基因型。利用该点选育自交系
(3)多倍体法
(4)平衡致死法
↓
表兄妹交配
↓
品种内交配
↓
品种间交配
↓
远缘杂交(种间及其以上)
近亲繁殖的程度,用近交系数(F)来度量:指个体的某个基因位点上两个等位基因来源于共同祖先某个基因的概率
F=0 – 1
2、自交的遗传效应
(1)杂合体通过自交,使后代群体中的遗传组成迅速趋于纯合化
自交代数AA×aa杂合体Aa纯合体AA+aa
↓
3、回交的遗传效应
回交(back cross):杂种后代与其两个亲本之一再次交配
如A×B→F1
F1×B→BC1,BC1×B→BC2……
F1×A→BC1,BC1×A→BC2……
BC1表示回交一代
BC2表示回交二代,其余类推
甲×乙甲:非轮回亲本
↓乙:轮回亲本(♂)
F1×乙
↓
BC1×乙
↓
BC2×乙
↓
……
回交也能使回交后代基因型纯合,但是:
四、近亲繁殖与杂种优势的利用
1、近亲繁殖在育种上的利用(自学)
2、杂种优势利用
(1)无性繁殖作物直接利用(甘薯)
(2)人工去雄,直接利用(玉米)
(3)雄性不育系的利用(水稻)
(4)化学杀雄(小麦)
注意不同利用途径与各植物之间的关系,不同利用途径的优缺点和限制因素
3、杂种优势固定
(1)无性繁殖法
(2)无融合生殖法
一、近亲繁殖及其遗传效应
1、近亲繁殖的概念
异交:亲缘关系较远的个体间随机相互交配
近交:亲缘关系相近的个体间交配按亲缘关系远近程度,可分为:
全同胞、半同胞、亲表兄妹
植物还有:自交、回交
自交是最极端的方式
自交
↓
回交(父女或母子交配)
↓
全同胞交配(同父母的兄妹交配)
↓
半同胞交配(同父或同母的兄妹交配)
2、杂种优势理论
(1)显性假说
双亲的显性基因全部聚集在杂种中产生互补作用。不能有效解释数量性状杂种优势的遗传机理
(2)超显性假说(等位基因异质结合假说)
杂种优势来源于双亲基因型的异质结合所引起的等位基因间的互作。未考虑不同基因位点之间相互作用即上位性效应
(3)上位性假说
杂种优势的遗传实质上在于数量性状基因各个遗传效应的综合作用:加性、显性、上位性、GE互作效应
适应型:杂种对外界不良环境适应能力较强
杂种优势表现特点:
(1)不是某一二个性状单独表现突出,而是许多性状综合表现突出。如产量
(2)杂种优势的大小,取决于双亲性状间的相对差异和相互补充。亲本选配
(3)杂种优势的大小与双亲基因型的高度纯合有关
单交种、双交种、品种间
(4)杂种优势大小与环境条件有关
F1表现杂种优势,F2出现衰退现象,因为F2会出现性状分离
1
(1 ---------)n
2r
因此,回交在基因型纯合进度上大于自交,一般回交5-6代即可
二、纯系学说
纯系学说:约翰生1909年提出。他以自花授粉的菜豆天然混杂群体为试验材料,按豆粒的轻重分别播种,从中选出19个单株。这19个单株的后代,即19个株系,在平均粒重上彼此具有明显的差异,而且是能够稳定遗传的。他又在19个株系中分别选择最轻和最重的两类种子分别种植,如此连续进行6年,发现在各年份里同一株系内轻粒和重粒的后代平均粒重几乎没有差异
纯系学说:
(1)在自花授粉植物的天然混杂群体中,可分离出许多基因型纯合的纯系。在一个混杂群体中选择是有效的
(2)在纯系内个体所表现的差异,那只是环境的影响,是不能遗传的。所以,在纯系内继续选择是无效的
局限性
纯系:从一个基因型纯合个体自交产生的后代(自交系)
三、杂种优势
1、杂种优势表现
杂种优势:两个遗传组成不同的亲本杂交产生的杂种F1在生长势、生活力、繁殖力、抗逆性、产量、品质等上优越于双亲的现象。杂种优势所涉及的性状大都为数量性状
(1)纯合基因型严格受轮回亲本的控制,1种纯合基因型;
自交是2n种纯合基因型
(2)纯合率同样用:
1
纯合体频率x= (1 -------)n
2r
但是,纯合率包含的内容不同
自交后代每一种基因型的纯合率为:
11
(1 ------------)n×(------)n
2r2
回交后代最后只有一种接近轮回亲本的纯合基因型,纯合率为:
F10Aa(1/2)00
↓
F211 AA 2Aa 1aa(1/2)11-(1/2)1
↓
F32 4AA 2 AA 4Aa 2aa 4aa (1/2)21-(1/2)2
. ..
Fr+1r(1/2)r1-(1/2)r
设有n对异质基因,自交r代
1
则纯合体频率x= (1 --------)n
2r
某一种纯合基因型的频率:
如果杂种的性状表现超过双亲的均值,这种优势称为中亲优势(平均优势)
F1-MP
中亲优势(%)=---------------×100%
MP
在实际育种中常使用超亲优势,即超越优亲的杂种优势
F1-HP
超亲优势(%)=----------×10旺
生殖型:生殖器官发育较盛
11
(1 -----------)n╳-----
2r2n
n对基因均杂合的个体的频率:
[1+(2r-1)]n的展开为Fr+1群体中各种个体数的分布
如:n=3 r=5
[1+(25-1)]3=1+93+2883+29791
(2)杂合体通过自交导致等位基因纯合,淘汰有害隐性纯合体
(3)杂合体通过自交,获取不同纯合基因型。利用该点选育自交系
(3)多倍体法
(4)平衡致死法
↓
表兄妹交配
↓
品种内交配
↓
品种间交配
↓
远缘杂交(种间及其以上)
近亲繁殖的程度,用近交系数(F)来度量:指个体的某个基因位点上两个等位基因来源于共同祖先某个基因的概率
F=0 – 1
2、自交的遗传效应
(1)杂合体通过自交,使后代群体中的遗传组成迅速趋于纯合化
自交代数AA×aa杂合体Aa纯合体AA+aa
↓
3、回交的遗传效应
回交(back cross):杂种后代与其两个亲本之一再次交配
如A×B→F1
F1×B→BC1,BC1×B→BC2……
F1×A→BC1,BC1×A→BC2……
BC1表示回交一代
BC2表示回交二代,其余类推
甲×乙甲:非轮回亲本
↓乙:轮回亲本(♂)
F1×乙
↓
BC1×乙
↓
BC2×乙
↓
……
回交也能使回交后代基因型纯合,但是:
四、近亲繁殖与杂种优势的利用
1、近亲繁殖在育种上的利用(自学)
2、杂种优势利用
(1)无性繁殖作物直接利用(甘薯)
(2)人工去雄,直接利用(玉米)
(3)雄性不育系的利用(水稻)
(4)化学杀雄(小麦)
注意不同利用途径与各植物之间的关系,不同利用途径的优缺点和限制因素
3、杂种优势固定
(1)无性繁殖法
(2)无融合生殖法