黄嘌呤氧化还原酶钼中心与配体的相互作用
乳中黄嘌呤氧化还原酶的研究进展

形式的统称 , 主要作用于嘌呤代谢过程。 因为它能 从乳中大量地提取纯化 ,0 年来已被广泛研究 。 10
直以来认 为 X R主要在生物体 的嘌呤代谢 中 O 起作用, 最近的研究发现 X R 在脂肪合成 、 O 消化 道抑菌、 先天免疫 、 心血管疾病和缺血再灌注的损
仅包括黄嘌呤和次黄嘌呤,还有广泛的醛类和杂 环分子。 在氧化过程中底物的电子传递给 X R的 O 钼位点, 而后再传递到 N D或者分子氧 , F D A 在 A 位 点 ,AD被 还 原成 N D N A H,分子 氧生 成 过氧 化 氢 和超 氧 阴离子 。X R有 两 种形 式 ,还原 型 O ( H) 氧化 型 ( O)并且 这 两种 形式 的分 子可 XD 和 X , 通过巯基 的氧化形成可逆转化 ,也可由蛋白酶水 解造成不可逆的转化。这两种酶的催化活力不尽
H n Lqa g,Y n uy a iin agG ou,Wa gY eig,G oY j n uyn u ui e
( ol eo nm lH sady& V t nr MeiieH n gi l rlU i ri ,hnzo 1C lg fA ia ubnr e ee a dcn , ea A rut a nv syZ eghu i r y n c u e t
维普资讯
22 1
《 乳业科学与技术》
20 年 第 5 06 期
( 总第 10 2 期)
乳中黄嘌呤氧化还原酶的研究进展
韩立强 ,杨 国宇 ,王 月影 ,郭豫杰
( 河南农业大学牧 医工程学院, 1 河南郑州 40 0 ;2 50 2 河南省农业生物技术重点开放实验室)
高尿酸血症治疗药物黄嘌呤氧化酶抑制剂的研究进展

高尿酸血症治疗药物黄嘌呤氧化酶抑制剂的研究进展陆海波;鲁传华【摘要】Hyperuricemia is a metabolic disease resulting from multiple factors,and xanthine oxidase (XO) is an important target for hyperuricemia therapy.Due to the determination of crystal structure of the enzyme and application of CADD and HTS,there emerge a large number of xanthine oxidase inhibitors which have good activity for lowering uric acid in recent ing traditional Chinese medicine to treat hyperuricemia is prevalent in China.It is an important method to modificate natural product to find new compounds. In this article,we review the compounds which have excellent inhibitory activity of xanthine oxidase based on the pathological mechanism.%高尿酸血症是由多种原因引起的代谢性疾病,黄嘌呤氧化酶(XO)是药物治疗高尿酸血症的重要靶点。
随着黄嘌呤氧化酶晶体结构的解析、计算机辅助药物设计和高通量筛选等技术的运用,近年涌现了许多具有良好的 XO 抑制活性的化合物。
中药也是我国治疗高尿酸血症的一种主要方式,对天然化合物进行结构修饰也是寻找新化合物的重要方法。
新生儿起病的钼辅因子缺乏症3例病例报告
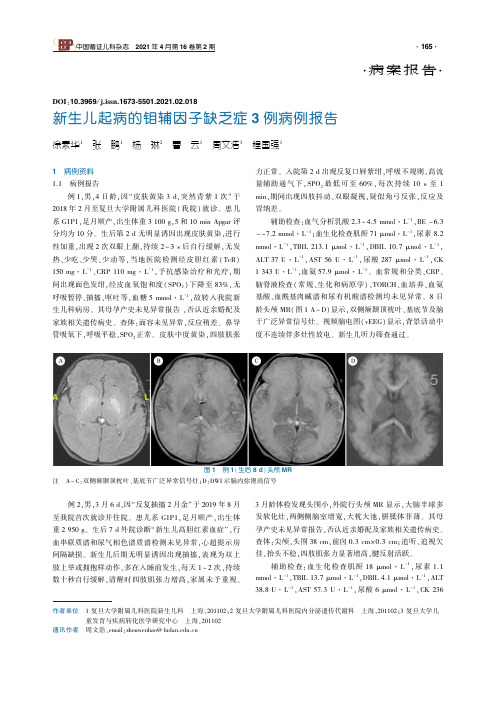
作者单位㊀1复旦大学附属儿科医院新生儿科㊀上海,201102;2复旦大学附属儿科医院内分泌遗传代谢科㊀上海,201102;3复旦大学儿童发育与疾病转化医学研究中心㊀上海,201102通讯作者㊀周文浩,email:zhouwenhao@fudan.edu.cn㊃病案报告㊃DOI:10.3969/j.issn.1673⁃5501.2021.02.018新生儿起病的钼辅因子缺乏症3例病例报告徐素华1㊀张㊀鹏1㊀杨㊀琳2㊀曹㊀云1㊀周文浩3㊀程国强11㊀病例资料1.1㊀病例报告㊀㊀例1,男,4日龄,因 皮肤黄染3d,突然青紫1次 于2018年2月至复旦大学附属儿科医院(我院)就诊㊂患儿系G1P1,足月顺产,出生体重3100g,5和10minApgar评分均为10分㊂生后第2d无明显诱因出现皮肤黄染,进行性加重,出现2次双眼上翻,持续2 3s后自行缓解,无发热㊁少吃㊁少哭㊁少动等,当地医院检测经皮胆红素(TcB)150mg㊃L-1㊁CRP110mg㊃L-1,予抗感染治疗和光疗,期间出现面色发绀,经皮血氧饱和度(SPO2)下降至83%,无呼吸暂停㊁抽搐㊁呕吐等,血糖5mmol㊃L-1,故转入我院新生儿科病房㊂其母孕产史未见异常报告,否认近亲婚配及家族相关遗传病史㊂查体:面容未见异常,反应稍差㊂鼻导管吸氧下,呼吸平稳,SPO2正常㊂皮肤中度黄染,四肢肌张力正常㊂入院第2d出现反复口唇紫绀,呼吸不规则,高流量辅助通气下,SPO2最低可至60%,每次持续10s至1min,期间出现四肢抖动㊁双眼凝视,疑似角弓反张,反应及胃纳差㊂㊀㊀辅助检查:血气分析乳酸2.3 4.5mmol㊃L-1,BE-6.3 -7.2mmol㊃L-1;血生化检查肌酐71μmol㊃L-1,尿素8.2mmol㊃L-1,TBIL213.1μmol㊃L-1,DBIL10.7μmol㊃L-1,ALT37U㊃L-1,AST56U㊃L-1,尿酸287μmol㊃L-1,CK1343U㊃L-1,血氨57.9μmol㊃L-1㊂血常规和分类㊁CRP㊁脑脊液检查(常规㊁生化和病原学)㊁TORCH㊁血培养㊁血氨基酸㊁血酰基肉碱谱和尿有机酸谱检测均未见异常㊂8日龄头颅MR(图1A D)显示,双侧额颞顶枕叶㊁基底节及脑干广泛异常信号灶㊂视频脑电图(vEEG)显示,背景活动中度不连续伴多灶性放电㊂新生儿听力筛查通过㊂图1㊀例1(生后8d)头颅MR注㊀A C:双侧额颞顶枕叶㊁基底节广泛异常信号灶;D:DWI示脑内弥漫高信号㊀㊀例2,男,3月6d,因 反复抽搐2月余 于2019年8月至我院首次就诊并住院㊂患儿系G1P1,足月顺产,出生体重2950g㊂生后7d外院诊断 新生儿高胆红素血症 ,行血串联质谱和尿气相色谱质谱检测未见异常,心超提示房间隔缺损㊂新生儿后期无明显诱因出现抽搐,表现为双上肢上举或拥抱样动作,多在入睡前发生,每天1 2次,持续数十秒自行缓解,清醒时四肢肌张力增高,家属未予重视㊂3月龄体检发现头围小,外院行头颅MR显示,大脑半球多发软化灶,两侧侧脑室增宽,大枕大池,胼胝体菲薄㊂其母孕产史未见异常报告,否认近亲婚配及家族相关遗传病史㊂查体:尖颅,头围38cm,前囟0.3cmˑ0.3cm;追听㊁追视欠佳,抬头不稳,四肢肌张力显著增高,腱反射活跃㊂㊀㊀辅助检查:血生化检查肌酐18μmol㊃L-1,尿素1.1mmol㊃L-1,TBIL13.7μmol㊃L-1,DBIL4.1μmol㊃L-1,ALT38.8U㊃L-1,AST57.3U㊃L-1,尿酸6μmol㊃L-1,CK236U㊃L-1,血氨82μmol㊃L-1㊂TORCH检查CMV⁃IgM㊁IgG阳性;血CMV⁃DNA阴性㊂血常规㊁CD系列㊁免疫球蛋白等未见异常㊂血氨基酸㊁酰基肉碱谱检测,甲硫氨酸㊁精氨酸略升高;尿有机酸谱检测,2⁃酮戊二酸㊁柠檬酸㊁乌头酸显著升高㊂听力及眼底检查未见异常㊂视觉诱发电位,有双侧波形可引出,分化差㊂vEEG显示,醒睡大量广泛及多灶性棘波㊁尖慢波㊁多棘慢波发放,右侧颞区及双侧后头部显著,睡眠时显著,家长指认抽动事件不伴同步异常放电㊂㊀㊀例3,男,10月龄,因 生后发育落后 于2015年7月至我院神经内科门诊就诊㊂患儿生后即出现发育落后,6月龄抬头,不能独坐㊂8月龄因 肠道感染 后出现抽搐,大发作,经治疗(具体不详)后,抽搐未再发作,但双眼向上凝视㊂外院血串联质谱检测未见异常;EEG:两侧适量尖波㊁尖慢波㊁棘慢波发放伴阵发,两前额部明显;头颅MR:双侧基底节㊁丘脑㊁脑白质异常信号;考虑 线粒体脑病(Leigh s)待排除 ,故至我院就诊㊂患儿系G1P1,足月顺产,出生体重3600g,其母孕产史未见异常报告,否认近亲婚配及家族相关遗传病史㊂查体:反应差,前额突出,双眼向上凝视,无眼球震颤,四肢肌张力增高明显,病理征阴性㊂1.2㊀基因检测㊀在取得患儿父母的知情同意后对3例患儿及其父母行基因检测,例1行新生儿panel检测,例2和3均行家系WES测序,根据我院高通量测序数据分析流程[1]进行分析,并行Sanger测序验证㊂表1显示,3例患儿均经基因诊断为钼辅因子缺乏症(MoCD,MIM:252150),例1和2为A型,例3为B型㊂例1检测到MOCS1基因的2个杂合突变c.45delG(p.A17Qfs∗79)和c.199C>T(p.R67W),c.199C>T为人类基因突变数据库(HGMD)已收录的致病突变位点,错义突变,来自患儿母亲;c.45delG为新发现的突变,移码突变,来自患儿父亲㊂例2检测到MOCS1基因的2个杂合突变c.203delinsCGGCA(p.Q68Pfs∗122)和c.646⁃2A>G,均为新发现的突变位点㊂c.203delinsCGGCA为移码突变,来自患儿母亲;c.646⁃2A>G为剪切突变,来自患儿父亲㊂例3检测到1个MOCS2基因的纯合无义突变,位于1号外显子的c.16C>T(p.Q6X),是HGMD已收录的致病突变位点㊂1.3㊀治疗和随访㊀例1予禁食㊁补液㊁机械通气㊁止惊㊁改善循环等治疗后,病情好转,自主呼吸平稳,无青紫㊁惊厥发作,喂养耐受㊂生后15d随访血尿酸49μmol㊃L-1,余指标未见异常㊂出院后门诊随访,基因明确诊断后家属放弃治疗㊂患儿1月龄起出现四肢肌张力增高,频繁四肢抽动,喂养困难,生长发育迟缓,随访清醒脑电图提示多量尖波㊁尖慢波发放㊂后反复发热,极度消瘦,18月龄死亡㊂㊀㊀例2在基因明确诊断后予康复干预,丙戊酸钠㊁托吡酯㊁ACTH等抗癫治疗㊂病程中表现为全面性发育迟滞,6月龄出现角弓反张,8月龄出现频繁成串痉挛发作,11月龄复查头颅MR(图2A D):脑发育不良,脑内多发软化灶形成;vEEG:背景变慢,醒睡多量多灶尖波㊁尖慢波㊁棘慢波㊁多棘慢波发放,后头部高度失律,清醒期多次孤立痉挛发作和1次强直发作㊂12月龄复查心超:左室后壁与室间隔增厚,心功能正常;予口服普萘洛尔㊂13月龄出现发育倒退,不能抬头㊁追视,末次随访22月龄时仍有频繁痉挛样发作㊂图2㊀例2(11月龄)头颅MR注㊀A㊁B:T2WI示皮层下白质斑片状高信号影;双侧基底节区可见小卵圆形脑脊液信号影;双侧脑室及三脑室扩张㊂C㊁D:胼胝体菲薄,大枕大池㊀㊀例3予康复干预及抗癫治疗,末次随访为5岁龄,全面发育迟滞,仍有小发作,喂养困难㊂2㊀讨论㊀㊀钼辅因子(MoCo)的生物合成途径高度保守㊂从遗传学上,可将MoCD分为3种类型(图3A)㊂A型MoCD(MIM:603707)因MOCS1基因变异导致MoCo合成的中间代谢产物环吡喃蝶呤单磷酸(cPMP)缺乏所致,占MoCD大多数;B型MoCD(MIM:603708)因编码含硫基因的钼蝶呤(MPT)合成酶亚基的MOCS2基因变异,或编码MOCS3硫酸化酶的MOCS3基因变异,导致MPT合成障碍所致;C型MoCD(MIM:603930)由编码催化钼与硫基团结合的桥尾蛋图3㊀钼辅因子(MoCo)合成途径(A)及钼辅因子缺乏症(MoCD)引起代谢异常(B)注㊀GTP:三磷酸鸟苷;cPMP:环吡喃蝶呤单磷酸;MPT:钼蝶呤;SOX:亚硫酸钠氧化酶;XOR:黄嘌呤氧化还原酶白的GPHN基因变异所致,目前仅有2例GPHN致病变异引起MoCD的报告[2,3]㊂㊀㊀截止2020年6月,HGMD已收录的MoCD相关基因突变类型如下㊂MOCS1基因突变:共37种,包括错义或无义突变21种㊁剪切突变6种㊁小片段缺失7种㊁小片段插入2种㊁大片段缺失1种;MOCS2基因突变:共16种,其中错义或无义突变14种㊁小片段插入1种㊁大片段缺失1种;MOCS3基因突变:仅1种错义突变;GPHN基因突变:共19种,包括大片段缺失14种㊁错义突变4种㊁剪切突变1种,其中与MoCD有关的致病变异仅2种㊂值得注意的是,GPHN的半合子缺失已在多种神经发育障碍中被描述,包括孤独症谱系障碍㊁精神分裂症和癫等[4]㊂本文例1和2中检测到MOCS1基因的2个新发移码变异c.45delG和c.203delinsCGGCA,导致蛋白质编码提前终止;例2检测到的c.646⁃2A>G为经典剪切位点;2例患儿表型均与MoCD相符,且父母均为携带者㊂㊀㊀体内MoCo依赖酶包括:亚硫酸钠氧化酶(SOX)㊁黄嘌呤氧化还原酶(XOR)㊁线粒体偕胺肟还原组分和醛氧化酶㊂SOX参与蛋氨酸和半胱氨酸等硫化氨基酸降解途径(图3B),其活性降低可导致蛋氨酸㊁半胱胺酸㊁亚硫酸盐㊁硫代硫酸盐㊁牛磺酸和S⁃磺酸半胱氨酸等代谢产物堆积,其中亚硫酸盐和S⁃磺酸半胱氨酸水平升高可导致神经系统进行性损害[5]㊂高效液相色谱法对检测尿液中亚硫酸盐和S⁃磺酸半胱氨酸有较高敏感度[6],可协助MoCD的诊断㊂XOR参与嘌呤代谢,其活性异常可导致尿酸生成减少㊁黄嘌呤水平升高(图3B)㊂因MoCD临床症状与影像学表现与亚硫酸氧化酶缺乏症(MIM:272300)十分相似,故血㊁尿液中尿酸下降㊁黄嘌呤水平升高,有助于两类疾病的鉴别㊂因对该类疾病的了解不够,在基因明确诊断以前,虽然本文例1和2病程中血尿酸有显著降低,但并未进一步检测尿亚硫酸盐㊁S⁃磺酸半胱氨酸㊁黄嘌呤等代谢产物㊂㊀㊀MoCD根据临床表现分为早发型和晚发型,以早发型居多,该病起病年龄中位数为生后第1d,但确诊年龄中位数为4.5月龄[7],此时患儿已出现进行性神经系统恶化㊂MoCD典型表现为新生儿难治性惊厥发作㊁喂养困难㊁过度惊跳反应㊁肌张力异常,需要与严重缺氧缺血性脑病(HIE)㊁氨基酸代谢异常等疾病相鉴别㊂随后出现进行性锥体和锥体外系功能障碍的临床症状㊂如不接受治疗,患儿仅存活1周至数月㊂病程超过1月龄后,患儿可出现严重的精神运动发育迟缓㊁生长迟缓和晚发性晶状体脱位,生活质量极差[7⁃9]㊂部分病例可有眼裂延长㊁厚嘴唇㊁宽人中㊁小鼻等面部畸形及小头畸形㊂本文例1在新生儿早期出现反复惊厥发作,疑似角弓反张,后期出现进食困难㊁极度消瘦㊁全面发育迟滞,最后死于恶病质㊂例2存在小头畸形,于新生儿晚期出现反复抽搐,后逐渐进展为婴儿痉挛症及全面发育迟滞㊂例3存在前额突出的面部畸形,生后发育较同性别同龄儿落后,8月龄因肠道感染后出现难治性癫,后逐渐出现全面发育迟滞和喂养困难㊂残留MoCo酶活性的部分轻型表型患儿,发病时间较晚,通常在其他疾病或感染后出现症状,如精神运动迟缓㊁锥体外系和锥体症状[10]㊂㊀㊀MoCD患儿头颅MR异常表现似乎按时间顺序发展,疾病早期可见全面性脑梗死所致脑水肿和扩散加权受限改变,基底神经节㊁丘脑等受累也较早可见,最终进展为严重的囊性脑软化;还可观察到胼胝体发育不良㊁皮质下囊肿㊁小脑脑萎缩㊁脑干发育不良等其他异常[11⁃14]㊂本文中例1生后8d头颅MR病变以弥漫性脑水肿为主,例2在3月龄及12月龄随访时头颅MR可见脑内多发软化灶形成㊂MoCD早期影像学发现易与严重HIE混淆,尤其是当患儿存在明确围生期窒息史时㊂越来越多的证据表明,部分病例在胎儿期头颅影像学已发生改变,目前确诊的病例中最早见于孕26周,胎儿B超见巨枕大池和第六脑室[8,15]㊂㊀㊀cPMP是MoCo最直接㊁最稳定的前体,目前研究已表明,A型MoCD患儿采用cPMP替代治疗安全且有效[16],在脑病发生前即予以替代治疗,患儿长期神经系统发育良好,但不能逆转已经产生的脑损伤㊂然而,cPMP目前仍处于临床试验性阶段㊂其他类型的MoCD尚缺乏特异性治疗,以对症支持治疗为主㊂㊀㊀综上所述,MoCD是一种罕见的遗传代谢病,主要临床症状为顽固性新生儿惊厥发作㊁喂养困难㊁肌张力异常㊁发育迟缓㊁面部畸形,其临床症状和影像学表现易与HIE㊁氨基酸代谢异常等混淆;血中尿酸水平下降,尿液中亚硫酸盐㊁S⁃磺酸半胱氨酸㊁黄嘌呤等水平升高,可协助诊断;及时基因诊断对确诊及产前诊断至关重要㊂参考文献1 黎籽秀 刘博 徐凌丽 等.高通量测序数据分析和临床诊断流程的解读.中国循证儿科杂志 2015 10 1 19⁃24.2 REISSJ LENZU AQUAVIVA⁃BOURDAINC etal.AGPHNpointmutationleadingtomolybdenumcofactordeficiency.ClinGenet 2011 80 6 598⁃599.3 REISSJ GROSS⁃HARDTS CHRISTENSENE etal.Amutationinthegenefortheneurotransmitterreceptor⁃clusteringproteingephyrincausesanovelformofmolybdenumcofactordeficiency.AmJHumGenet 2001 68 1 208⁃213.4 LIONELAC VAAGSAK SATOD etal.RareexonicdeletionsimplicatethesynapticorganizerGephyrinGPHN inriskforautism schizophreniaandseizures.HumMolGenet 2013 22 10 2055⁃2066.5 ZHANGX VINCENTAS HALLIWELLB etal.Amechanismofsulfiteneurotoxicity directinhibitionofglutamatedehydrogenase.JBiolChem 2004 279 4143035⁃43045.6 BELAIDIAA ARJUNES SANTAMARIA⁃ARAUJOJA etal.Molybdenumcofactordeficiency anewHPLCmethodforfastquantificationofs⁃sulfocysteineinurineandserum.JIMDRep 2012 5 35⁃43.7 MECHLERK MOUNTFORDWK HOFFMANNGF etal.Ultra⁃orphandiseases aquantitativeanalysisofthenaturalhistoryofmolybdenumcofactordeficiency.GenetMed 2015 17 12 965⁃970.8 ALONZOMM CAZORLAE CANOVASE etal.Molybdenumcofactordeficiency Megacisternamagnaintwoconsecutivepregnanciesandreviewoftheliterature.ApplClinGenet 2020 13 49⁃55.9 ARICANP GENCPINARP KIRBIYIKO etal.TheclinicalandmolecularcharacteristicsofmolybdenumcofactordeficiencyduetoMOCS2mutations.PediatrNeurol 2019 99 55⁃59.10 SCELSAB GASPERINIS RIGHINIA etal.MildphenotypeinMolybdenumcofactordeficiency Anewpatientandreviewoftheliterature.MolGenetGenomicMed 2019 76 e657.11 NAGAPPAM BINDUPS TALYAB etal.ChildNeurology Molybdenumcofactordeficiency.Neurology 2015 85 23 e175⁃e178.12 HIGUCHIR SUGIMOTOT TAMURAA etal.Earlyfeaturesinneuroimagingoftwosiblingswithmolybdenumcofactordeficiency.Pediatrics 2014 133 1 e267⁃e271.13 SASSJO GUNDUZA ARAUJORFC etal.Functionaldeficienciesofsulfiteoxidase Differentialdiagnosesinneonatespresentingwithintractableseizuresandcysticencephalomalacia.BrainDev 2010 32 7 544⁃549.14 KIKUCHIK HAMANOS MOCHIZUKIH etal.Molybdenumcofactordeficiencymimicscerebralpalsydifferentiatingfactorsfordiagnosis.PediatrNeurol 2012 472 147⁃149.15 LUBOUTC DERKST MEINERSL etal.MolybdenumcofactordeficiencytypeA PrenatalmonitoringusingMRI.EurJPaediatrNeurol 2018 22 3 536⁃540.16 SCHWAHNBC VANSPRONSENFJ BELAIDIAA etal.EfficacyandsafetyofcyclicpyranopterinmonophosphatesubstitutioninseveremolybdenumcofactordeficiencytypeA aprospectivecohortstudy.Lancet 2015 386 10007 1955⁃1963.(收稿日期:2020⁃06⁃30㊀修回日期:2021⁃04⁃01)(本文编辑:孙晋枫)。
钼提高植物抗逆性研究进展

第42卷 第6期2023年 11月华中农业大学学报Journal of Huazhong Agricultural UniversityVol.42 No.6Nov. 2023,50~58钼提高植物抗逆性研究进展秦晓明,赵优优,武松伟,胡承孝,孙学成华中农业大学新型肥料湖北省工程实验室/微量元素研究中心,武汉430070摘要 钼(Mo )作为植物必需的微量元素,在促进植物生长发育和增强植物抗逆性方面发挥着关键作用。
植物对钼的吸收转运主要受到钼酸盐转运蛋白基因MOT1和MOT2调控,钼进入植物体内以含钼酶形式参与植物生长代谢,其中对植物抗逆性方面的调控主要表现为:钼通过含钼酶硝酸还原酶、醛氧化酶、黄嘌呤脱氢酶影响植物体内的光合碳氮代谢、激素合成和活性氧代谢进而调控植物抗寒性;钼通过硝酸还原酶和醛氧化酶介导的信号转导过程调控根系发育、养分水分利用及抗旱基因表达,进一步影响脂质合成与代谢调控植物抗旱性;最新研究还发现钼在植物适应盐胁迫、缓解重金属胁迫方面也具有重要作用。
这些研究结果为通过钼营养调控提升植物的抗逆性提供了新思路。
关键词 钼; 钼酶; 转运蛋白; 抗寒; 抗旱; 抗盐; 重金属抗性中图分类号 S143.7+1 文献标识码 A 文章编号 1000-2421(2023)06-0050-09农业生产会受到多种环境胁迫(如寒冷、干旱、盐碱、重金属等)的影响,如何通过营养调控提高植物的抗胁迫能力一直是科学家们关注的热点[1]。
大量元素磷、钾提高作物抗逆性的效应及机制较为明确,而微量元素与作物抗逆性的关系报道较少。
钼是植物体必需的微量元素,它在植物体的生理功能主要通过含钼酶来实现。
较早的研究发现低温处理下施钼增加了硝酸还原酶和黄嘌呤脱氢酶的活性,进而提高植物的低温耐受性[2]。
近年来,越来越多的研究证实钼不仅可提高植物抗寒性,还能提高植物抗旱、抗盐胁迫及抗重金属胁迫的能力。
本文以钼的吸收和转运、含钼酶调控的代谢过程为主线综述钼提高植物抗逆性的生理及分子机制,旨在为通过钼营养调控提升植物的抗逆性提供理论依据。
钼的生物学作用及钼缺乏对生物体的影响_韩冰

钼的生物学作用及钼缺乏对生物体的影响韩冰,张菁华(菏泽医学专科学校,山东 菏泽 274030))关键词: 钼;生物学作用;钼/缺乏;影响中图分类号: Q581;O614.61+2 文献标识码:A 文章编号:1008-4118(2009)01-0073-02 钼广泛存在于水、土壤以及各种动植物体内。
1953年,DeRenzo 等研究发现,大鼠饲料中添加不同水平的钼对其体内黄嘌呤氧化酶活性产生显著影响。
1954年,Mahcer 等的实验结果表明,兔的肝脏中存在一种含钼的酶即醛氧化酶。
后来Cohen 等进一步研究发现,钼还是亚硫酸盐氧化酶的金属组分,从而证实了钼是动物体内必需的微量元素[1]。
现已证实,钼是黄嘌呤氧化酶、醛氧化酶和亚硫酸盐氧化酶等的组成成分。
1 钼的生物学作用1.1 参与人体内糖和脂肪的代谢,促进发育[2] 糖和脂肪属于营养物质,在生物体的生长发育中发挥重要作用。
钼参与糖和脂肪的代谢,进而参与人体物质代谢和能量代谢,促进发育。
1.2 能增强氟在体内的储留 动物实验表明,钼能增加氟在大鼠体内的贮留,增强其作用效果[3]。
1.3 参与黄嘌呤氧化酶、醛氧化酶和亚硫酸盐氧化酶等的组成[4] 钼可参与硫、铁、铜之间的相互反应。
钼是黄嘌呤氧化酶、醛氧化酶和亚硫酸盐氧化酶发挥生物活性的必需因子。
黄嘌呤氧化酶主要参与体内的电子传递过程,负责向细胞色素C 转运电子,参与体内嘌呤化合物的氧化代谢以及尿酸的形成。
此外,调节体内铁的代谢,促进肝铁蛋白中铁的释放,促使铁进入血浆,催化Fe 2+转化为Fe 3+并与β1球蛋白结合成运铁蛋白供机体利用。
醛氧化酶参与细胞内的电子传递过程和体内醛的氧化,使醛氧化成羧酸,消除有毒醛类物质对人体的危害。
亚硫酸盐氧化酶在机体消化道内微生物区系中发挥重要作用[1]。
1.4 抑制肿瘤发生 钼是抗氧化剂,有阻断过氧化物生成的作用,从而可防止氧化损伤。
钼离子能促进细胞内氧化还原过程,特别是具有抑制亚硝基的致癌作用。
黄嘌呤氧化还原酶抑制剂的构效关系研究进展

黄嘌呤氧化还原酶抑制剂的构效关系研究进展
苏彦雷;蒋建勤;汪俊松;王明时
【期刊名称】《药学进展》
【年(卷),期】2009(33)8
【摘要】介绍了近年来黄嘌呤氧化还原酶及其抑制剂的研究进展.从构效关系的角度出发,概述与黄嘌呤氧化还原酶底物结构类似的抑制剂和非嘌呤类抑制剂的特点,为新的黄嘌呤氧化还原酶抑制剂的研发提供参考.
【总页数】10页(P350-359)
【作者】苏彦雷;蒋建勤;汪俊松;王明时
【作者单位】中国药科大学天然药物化学教研室,江苏,南京,210009;中国药科大学天然药物化学教研室,江苏,南京,210009;中国药科大学天然药物化学教研室,江苏,南京,210009;中国药科大学天然药物化学教研室,江苏,南京,210009
【正文语种】中文
【中图分类】R589.7;R914.2
【相关文献】
1.乳中黄嘌呤氧化还原酶的研究进展 [J], 韩立强;杨国宇;王月影;郭豫杰
2.高尿酸血症治疗药物黄嘌呤氧化酶抑制剂的研究进展 [J], 陆海波;鲁传华
3.天然产物来源的黄嘌呤氧化酶抑制剂研究进展 [J], 谭明亮;陈刚
4.药用植物中黄嘌呤氧化酶抑制剂的研究进展 [J], 张晨辉;谢雄雄;曾金祥;谢晶;李敏;钟国跃
5.黄嘌呤氧化还原酶抑制剂研究进展 [J], 朱深银;周远大;杜冠华
因版权原因,仅展示原文概要,查看原文内容请购买。
黄嘌呤氧化还原酶的结构、功能和作用

黄嘌呤氧化还原酶的结构、功能和作用李丽书;陈献华;邵叶波;刘璇;徐平【期刊名称】《细胞生物学杂志》【年(卷),期】2004(26)4【摘要】黄嘌呤氧化还原酶(XOR)参与人体内的嘌呤代谢,并且是这一代谢过程的限速酶。
其终产物是活性氧(包括OH·、H2O2和O2-)和尿酸。
这两种产物参与体内多种生理活动。
从XOR基因的结构、XOR蛋白的分子结构和基本功能、控制XOR活性的多个环节以及XOR的两种催化产物活性氧和尿酸在生理和病理情况下的功能及机制进行了总结,以期对XOR的发现、研究历史及现状和有待解决的问题有一个系统的了解。
【总页数】4页(P381-384)【关键词】黄嘌呤氧化还原酶;尿酸;生理;嘌呤代谢;活性氧【作者】李丽书;陈献华;邵叶波;刘璇;徐平【作者单位】复旦大学生命科学学院基因生理学实验室脑研究中心;复旦大学上海医学院【正文语种】中文【中图分类】Q554【相关文献】1.黄嘌呤氧化还原酶钼中心与配体的相互作用 [J], 王传铭;张旭东;王栋;谢昆2.1,3-丙二醇氧化还原酶结构与功能关系的探讨 [J], 林锦霞;张光亚;方柏山3.精索静脉曲张病人血中一氧化氮合成酶及黄嘌呤氧化酶的活性:氧化氮及过氧化氮对精子功能不良所起的作用 [J],4.捻转血矛线虫NADH:泛醌氧化还原酶结构域包含蛋白基因的克隆表达及功能分析 [J], 吴玲燕;王玉俭;温玉玲;严若峰;徐立新;宋小凯;李祥瑞5.含有WW结构域的氧化还原酶的基因在骨肉瘤组织中表达的临床意义及其对骨肉瘤的调控作用 [J], 许峰;王帝;李哲;元耀博;李隆卿;张卫红;闻嘉;谢祎;李家平;李甲振;张岩;卢新昌;张翼;刘永奎因版权原因,仅展示原文概要,查看原文内容请购买。
黄嘌呤氧化酶基因

黄嘌呤氧化酶基因
黄嘌呤氧化酶基因是人类基因组中的一个关键基因,也是代谢相关基因中的重要成员之一。
它位于染色体7上,由16个外显子和15个内含子组成,编码一种催化黄嘌呤代谢的酶——黄嘌呤氧化酶(XO)。
黄嘌呤氧化酶是一种半胱氨酸依赖性的氧化还原酶,主要参与嘌呤代谢途径中的次要分支途径。
该酶的催化作用是将黄嘌呤和次黄嘌呤分别氧化为尿酸和异黄嘌呤,产生的尿酸和异黄嘌呤可以通过肾脏排泄出体外。
由于黄嘌呤氧化酶在嘌呤代谢途径中具有重要作用,因此突变或缺陷的黄嘌呤氧化酶基因会导致许多代谢性疾病,如高尿酸血症、痛风等。
此外,黄嘌呤氧化酶还参与一些非代谢性过程,如肿瘤细胞的增殖、心血管疾病等,因此在某些疾病的治疗中也具有一定的临床意义。
黄嘌呤氧化酶基因的突变和多态性已经被广泛研究。
目前发现的突变包括点突变、插入、缺失等,这些突变都可能导致酶活性的降低或者丧失。
另外,该基因中还存在一些常见的单核苷酸多态性,如rs17038412和rs2231142等。
这些多态性可以影响基因表达水平和酶活性,并与一些疾病的发生风险相关。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
硫 /
吣 \ 一 点. 本文根据P D B 数据库中现存的晶体结构和国内 外学者多年来在此酶的催化过程、抑制机制等方面开展的实验和理论研究,对其
钼中心与底物和抑制剂之 间的相互作用形式进行了综述。
关键词 :黄嘌呤氧化还原酶 ;抑制剂 ;底物 ;钼辅 因子 ;分子模拟
中 图 分 类 号 :Q7 文 献 标 识 码 :A 文章 编 号 :1 0 0 8 — 9 1 2 8( 2 0 1 3) 0 2 — 0 0 4 5 — 0 7
的相 互 作 用 形式 方 面 的 研究 进 展 展 开综 述 ,以期 帮 助人们了解其催化过程及酶活抑制的分子机理.
1 钼碟 呤中心 的结构与功能
.’
。
截至 2 0 1 2 年7 月 ,P D B ( P r o t e i n D a t a B a n k )  ̄已经 收录- i " 3 5 个实验测定的X O R 结构( 表1 ) . 从这些解析的 数据 可 以看 出f 2 l 3 】 ,X O R 催 化 位 点包 含 三个 氧 化还 原 中 l : , ( r e d o x c e n t r e ) :一个 钼 蝶 呤辅 因子 、两 个 F e 2 S 2 中心 和 一 个 核 黄 素 腺 嘌 呤 二 核 苷 酸 辅 因子 ( F A D ) , 三个 部 分 几 乎呈 线 性 排列 ( 如 图1 A ) . 其 中钼 蝶 呤 中心 是X O R 催化关键位点 ,使体 内的黄嘌呤成为尿酸 , 在 此 过 程 中钼 中心首 先 失 去2 个 电子 ( 由六 价成 为 四 价) ,经铁硫 中心传 递 ,然后通 过F A D 将 电子传给 N A D 或氧分子 ,将其转化成N A D H 或 .
第l 1 卷 第 2 期 2 0 1 3 年4 月
红 河学院学报
J o u r n a l o f I t o n g h e U n i v e r s i t y
V o 1 . 1 1 N o . 2 A p r . 2 01 3
黄嘌 呤氧化还原 酶钼 中心与配体个辅 因子的空间位置关 系( 以P D B : 3 U N C为例) B :钼蝶 岭辅 因子 的简化模型
在理论模拟 中,钼辅因子 中的蝶呤部分常以不 合的硫原子及其相连的两个碳原子( 图1 B ) . 催 化关键 部位 是 一 个 以钼 为 中心 原子 的 四角锥 形 配 位结 构 , 同形式 简化 以减少计算量 ,其 中吡喃环和吡嗪环在 其具体配位方式人们还存在不同看法理解. 相对统一 模拟中可以提供足够的信息 ,相 比之下蝶呤部分远 的理 解是 :四角锥 顶 端是 氧原子 ( Mo = O 。 D ) ,底部 四边 离钼原 子的第二个环 可以略去 ,对计算可靠性影 响 形的三个端点分别是两个 与蝶岭亚基结合的硫原子 S - M o ) 和一个羟基( M o — O H ) . 而第四个顶点( x ) 可能有 不大[ 4 1 . 最简化 的方式是只保 留两个与钼原子配位结 (
作为第6 族( vI B 族 ,铬 族 ) 的过 渡 金 属 ,钼 f Mo l y b d e n u m, Mo ) 是 第 二 过 渡 系 中唯 一 一 种 对 地 球 上几乎所有生命都不可或缺的元 素[ 1 J . 在哺乳动物体 内 ,钼 主要 以 钼蝶 呤 ( mo l y b d o p t e r i n ,MP T ) 的形 式 存 在 于 黄 嘌呤 氧化 还 原 酶( x a n t h i n e o x i d o r e d u c t a s e , X O R ) 等三类金属酶 中,并 以其高的可变化合价参与 到 氧化 还原 过程 之 中. X O R 是广泛分布于各种生物体 内调控核酸代谢 过程的一种重要酶类 ,主要功能是催化嘌呤代谢 的 最 后 两 步 ,即次 黄 嘌 呤 氧化 为 黄 嘌 呤 和黄 嘌 呤进 一 步 氧化 为 尿 酸. 这 些生 化 反应 在 氮 循环 和 能量 代 谢过 程 中具 有重要作用 ,故有关此酶的结构 、功能和活 性 调控 研 究一 直 是 生物 医学 界关 注 的热 点 . 本 文试 从 结构生物学 和计算化学的角度对此酶的催化关键位 点( 钼蝶呤 中心) 与其相应配体( 底物 、抑制剂等) 之间
收稿 日期 :2 0 1 2 — 0 8 — 0 6 基金项 目:红河学 院中青年 学术带头人后备人才基金( 2 0 1 0 P Y O 1 0 4 ) ;红河学 院科研 基金项 目( 项 目编号 I O X J Y1 1 3 )
第一作者 :王传铭 ( 1 9 7 1 一),男 ,河南人 , 副教授 , 硕 士 ,研究方 向:生物信 息学 、分 子模 拟.
红河学院学报
2 0 1 3 . 2 / 生物学体育农学研究
硫( Mo = S ∞ ) 和氧 ( M0 = O q ) 两 种 情 况. Mo = S 。 方式 是 有 活 性的结构 ,也是 目前公认 的X O R 家族 的模式特征 , 被广泛应用在绝大多数理论计算中[ 5 - 7 1 .  ̄M o = O 明 结构 没有 活性 ,并 且在 此 种结 构 中用 S 印 替 换O 印 后x OR 的
王传铭 ,张旭 东 ,王 栋 ,谢 昆
l
( 1 . 红 河 学院云 南省 高校 农作物 优质 高效栽培 与安 全控 制 实验 室 云 南蒙 自 6 6 1 1 0 0 ;
2 . 云南大学生命科学学院 昆明 6 5 0 0 9 1 )
,
摘 要 :黄嘌呤氧化还原酶是一种分布广泛 的核 酸代谢调控酶类 ,过渡金属钼和蝶呤 等辅 因子构成其主要的催化活性位