第十三章 B淋巴细胞介导的体液免疫应答
B细胞介导的免疫应答

第十三章B淋巴细胞介导的体液免疫应答一、单项选择1. 再次体液免疫应答时,下述哪项是正确的?A. 抗体产生的潜伏期短B. 抗体亲和力低C. 抗体维持时间短D. 以IgM抗体为主E. 刺激的抗原与初次不同2. 体液免疫应答主要由下列哪种免疫细胞所介导:A. T细胞B. B细胞C. NK细胞D. 吞噬细胞E. 上皮细胞3. 下述哪项不是记忆性B细胞的特点:A. 大小与静止B细胞相仿B. 可持续分泌高水平IgC. CD27+D. 再次与相同抗原相遇时可迅速活化并分泌大量IgE. 存活时间较长4. B细胞对以下哪种抗原的应答需要T细胞辅助:A. 白喉毒素B. 脂多糖C. 荚膜多糖D. 金黄色葡萄球肠毒素(SEA)E. 以上都不是5. 下列哪种描述不是再次免疫应答的特点:DA. 潜伏期短B. 抗体浓度增加快,快速到达平台期C. 抗体维持时间长D. 抗体水平持续很低E. 诱发再次应答所需抗原剂量小6. 与体液免疫应答无关的是:A. 中和作用B. 调理作用C. Fas/FasL途径杀伤肿瘤细胞D. ADCCE. 参与补体的溶细胞作用7. 关于B细胞介导的免疫应答,哪项是错误的?A.对TD-Ag的应答可以产生记忆细胞B.亲和力成熟需要有抗原的诱导C.体细胞高频突变发生于中枢免疫器官D.浆细胞是B细胞分化的终末细胞之一E.发生Ig类别转换的遗传学基础是同一V区基因与不同重链C区基因的重排8. 可产生免疫记忆的细胞是A.B1 细胞B.γδT细胞C.αβT细胞D.NK细胞E.巨噬细胞9. 能特异性识别结合APC表面抗原肽-MHC分子复合物的淋巴细胞是A.γδT细胞B.B1细胞C.B2细胞D.αβT细胞E.NK细胞10. 适应性免疫的特点是A.经遗传获得B.包括物理屏障和化学屏障作用C.无针对病原体的特异性D.感染早期迅速发挥作用E.可产生免疫记忆11.适应性免疫应答所不具备的特点是A.淋巴细胞与相应抗原的结合具有高度特异性B.具有再次应答的能力C.无需抗原激发D.T/B细胞库具有高度异质性E.精确区分“自身”和“非己”12. B细胞表面的抗原识别受体是:A. CD40B. BCRC.CD19D. CD21E.Igα/Igβ13.关于B细胞表面同时表达的IgM和IgD,下列说法正确的是:A. 具有相同的重链和不同的轻链B.除CH区外其余相同C.除VH区外其余相同D. 具有不同VH和VL区E. 具有不同的CH和CL区14. 对B细胞内化抗原描述错误的是:A. B细胞活化的第一信号是抗原与BCR结合后产生B. 只有产生了第一信号,B细胞才能内化抗原C. 内化的抗原可经MHC I类分子途径提呈D. 内化的抗原可经MHC II类分子途径提呈E. 抗原信息提呈给抗原特异性Th细胞15. B细胞识别特异性抗原:A. 具有MHC I类分子限制性B. 具有MHC II类分子限制性C.没有MHC限制性D. 需要细胞因子E. 通过CD3传递抗原信号16. 细胞间相互作用不受MHC限制的是:A. NK细胞与靶细胞B. CTL细胞与靶细胞C. Th细胞与B细胞D. 巨噬细胞与Th细胞E. 树突状细胞与Th细胞17. B细胞表面传递特异性抗原信号的分子是A. mIgB. MHC 分子C. CD4D. CD3E.Igα/Igβ18. 对Igα/Igβ描述错误的是:A. 与mIg组成BCR复合物B. 胞浆区有ITAMC.可传导B细胞第一活化信号入胞D. 通常以二聚体的形式存在E. 能与抗原非特异性结合19. B细胞表面能结合C3d的分子是A. CD19B. CD81C. CD21D. BCRE. MHC 分子20. 以下CD分子中与B细胞活化有关的是:A. CD2B. CD3C. CD4D. CD21E. CD821. B细胞不表达的分子是A. C3d受体B. CD2C. I类MHC分子D. II类MHC分子E.CD1922. 对B细胞活化共受体描述正确的是:A. 是由CD19与CD21、CD81以非共价键形式组成的复合物B. CD21可与C3d结合,但不能转导信号C.信号由CD19传向胞内可加强BCR复合物传导的信号D. 上列A和B两项E. 上列A、B和C三项23. Th细胞活化静止B细胞主要靠下列哪两种分子间的相互作用A. B7和CD28B. B7和CTLA-4C. CD40L和CD40D. CD4和MHC II 类分子E. ICAM 和LFA-124. 作为抗原提呈细胞,B细胞通过下列哪种分子提供T细胞活化的第二信号A. CD28B. CD40C. CD40LD. B7E. MHC II类分子25. 下列哪种细胞高表达CD40LA. B细胞B. 树突状细胞C. 静止的T细胞D. 活化的CD4+ T细胞E. 巨噬细胞26. T细胞辅助抗原特异性B细胞是在A. 淋巴组织的T细胞区和生发中心B. 骨髓C. 胸腺D. 皮肤E. 粘膜27. 关于体细胞高频突变,错误的说法是A. 发生在V区基因B. 增加了抗体的多样性C. 在抗原诱导下发生D. 发生在生发中心E. 与抗体亲和力成熟无关28. 可直接调节Ig类别转换的细胞是A. Th细胞B. Tc细胞C.NK细胞D. B细胞E. 中性粒细胞29.发生类别转换前,B细胞表达A. IgAB. IgEC. IgM和IgDD. IgGE. IgG和IgD30. 浆细胞A. 能分泌抗体B. 来源于T细胞C. 发育为B细胞D. 分泌大量IFN-γE. 具有细胞毒作用31. 关于记忆性B细胞,下列说法错误的是A. 在生发中心形成B. 与静息B细胞的大小相似C. 合成大量IgD. 接触同一抗原可迅速活化E. 介导B细胞的再次免疫32. 具有免疫记忆的细胞是A. 巨噬细胞B. 中性粒细胞C. 淋巴细胞D. 肥大细胞E. NK细胞33. 关于TI抗原描述错误的是:A. 即胸腺非依赖性抗原B. 如某些细菌多糖及脂多糖C. 能在无胸腺鼠诱导Ab应答D. 能刺激初始B细胞E. 可诱导T细胞应答34. 对TI-1抗原描述错误的是A. 又称B细胞丝裂原B. 高浓度时多克隆活化B细胞C. 低浓度时激活抗原特异性B细胞D. 诱导B细胞应答比TD抗原早E. 可诱导产生记忆性B细胞35. TI-1抗原单独不足以诱导A. 抗体亲和力成熟B. Ig类别转换C. 记忆B细胞形成D. 上列A和B两项E. 上列A、B和C三项36. 对TI-2原描述错误的是A. 多为细菌胞壁与夹膜多糖B. 有高度重复序列C. 可激活不成熟B细胞D. 抗原表位密度太低不足以激活B细胞E. 抗原表位密度太高可使B细胞无应答37. 对TI-2抗原发生应答的主要细胞是A. 记忆性B细胞B. 前B细胞C. 不成熟B细胞D.B-1细胞E. T细胞38. 关于TI-2抗原,下列说法错误的是A. 具有重复序列B. 仅能激活成熟的B细胞C. 主要应答细胞是B1细胞D. 能诱导婴幼儿抗体应答E. 无需Th细胞辅助39. 不能诱导婴幼儿产生有效抗体应答的是A. TI-1抗原B. TI-2抗原C. TD抗原D. 上列A和B两项E. 上列A、B和C三项40. Th细胞辅助抗体产生A. 依靠T细胞识别结合于BCR的完整抗原B. 依靠T细胞识别经APC细胞处理、提呈的抗原C. 涉及B细胞上的MHC I类分子D. 可发生于经X-线照射的小鼠E. 发生在对LPS的应答过程中41. 在B细胞对TD抗原的应答中,Th细胞在下列哪项不发挥作用:A. 对抗原的特异性识别B细胞的活化 C. 生发中心的形成D. Ig类别转换E. 抗体亲和力成熟42. 抗体产生过程中,下列说法错误的是:A. Th细胞与B细胞的相互作用受MHC限制B. 浆细胞是产生抗体的细胞C. TI抗原刺激B细胞产生抗体需T细胞的辅助D. 应答早期主要产生低亲和力的IgME. Ig的类别转换需要Th细胞的辅助43. 下列哪种物质可以被动转移特异性体液免疫A. 抗体B. IL-2C. TNFD. T细胞E. IL-144. 免疫应答过程不包括A. 细胞在骨髓内分化成熟B. B细胞对抗原的特异性识别C. 巨噬细胞对抗原的处理和提呈D. T、B细胞的活化、增殖、分化E. 效应细胞产生效应分子45. TD抗原引起免疫应答的特点是A. 产生免疫应答的细胞为B1细胞B. 只诱导体液免疫应答C. 可形成记忆性细胞D. 可直接诱导T、B细胞产生免疫应答E. 只诱导细胞免疫应答46. 关于再次应答,下列说法错误的是A. 再次应答较初次应答的潜伏期短B. 抗体浓度增加快C. 抗体维持时间长D. 诱发再次应答所需抗原剂量大E. 主要产生高亲和力的IgG抗体47. 初次应答中产生的抗体主要是A. IgGB. IgMC. IgED. IgDE. IgA48. B细胞介导的再次应答的强弱主要取决于A. 抗原的化学性质B. 再次接触的抗原量C. 两次抗原刺激的间隔长短D. 抗原进入体内的方式E. 抗原分子量大小49. 对体液免疫再次应答Ab产生描述有误的是A. 再次接受较少量相同Ag刺激即可诱发B. 潜伏期大约为初次应答时的一半C. 达平台期快,平台高且时间长D. 主要为IgG,亲和力高且较均一E. 下降期短或无50. 再次免疫应答是基于A. 记忆性细胞活化B. 补体激活C. 肥大细胞脱颗粒D. 巨噬细胞的吞噬作用E.NK细胞的杀伤作用51. 造成抗体多样性的机制不包括:A. 组合造成的多样性B. 连接造成的多样性C. 体细胞高频突变造成的多样性D. 重链和轻链间的组装E. 抗体亲和力成熟52. 编码Ig重链可变区的基因片段是A. V、D、J片段B. V、J片段C. D、J片段D. C片段E. S区53. 关于体细胞高频体突变,错误的说法是A. 发生于已完成V基因重排的B细胞B. 发生在抗原刺激后外周淋巴器官生发中心的B细胞C. 突变的主要方式是点突变D. 能增加抗体的多样性E. 与抗体的亲和力成熟无关54. B细胞区别于T细胞的特征是A. 可被美洲商陆(PWM)激活B. 表达膜型Ig直接识别抗原C. 表达CD3分子D. 属于淋巴细胞E. 可分化为记忆性细胞55. 关于T 细胞和B细胞介导的免疫应答,下列说法错误的是:A. 对TD-Ag的应答都产生记忆性细胞B. 均产生效应产物C. 效应产物的作用都是特异性的D. 都能形成免疫耐受E. 对TD-Ag的应答均需T细胞的参与二、不定项选择1. 在抗体形成过程中,下列哪项叙述是正确的:A. 浆细胞是产生抗体的细胞B. 所有B-1细胞活化都必须有双信号C. 对TD抗原的应答不需Th细胞参加D. Th细胞和B细胞的相互作用受MHC限制E. 再次应答时抗体产生快,效价高2. Ig同种型转换:A. 在抗原诱导下发生B. 受T细胞分泌的细胞因子的调节C. IgV区不变D. IgC区不变E. IgV区和IgC区均改变3. Igα/Igβ的主要功能是:A.转导抗原与BCR结合产生的活化信号B.转导抗原与TCR结合产生的活化信号C.参与B细胞活化的第一信号转导D.作为协同刺激分子促进淋巴细胞活化E.T细胞活化的辅助受体4. T细胞辅助B细胞是在:A. 淋巴组织的B细胞区B. 淋巴组织的生发中心C. 脂筏D. 外周淋巴组织的T细胞区E. 冠状带5. 对TD-Ag的免疫应答过程包括:A.APC对抗原的摄取、处理和提呈B.T、B细胞对抗原的特异性识别C.T细胞在胸腺内的发育D.T、B细胞的活化、分化、增殖E.效应细胞与效应分子的产生和作用6. 具有免疫记忆的细胞有:A. APCB. NK细胞C. B细胞D. T细胞E. 中性粒细胞三、名词解释1. Ig亲和力成熟2. Ig的类别转换3. 再次应答4. 亲和力5. 亲合力四、简答与论述1. Th细胞在B细胞的免疫应答有何作用?2. 体液免疫应答中的再次应答和初次应答有何不同?3. 抗体的多样性是怎样形成的?。
第十三章 B淋巴细胞介导的特异性免疫应答

第十三章B淋巴细胞介导的特异性免疫应答抗原进入机体后诱导相应的抗原特异性B细胞活化、增殖并最终分化为浆细胞,产生特异性抗体进入体液,发挥免疫效应。
由于抗体存在于体液,故此过程也称为体液免疫应答(humoral immune response)。
B细胞介导的免疫应答依据抗原的不同可分为对T细胞依赖抗原(TD-Ag)的免疫应答和对T细胞非依赖抗原(TI-Ag)的免疫应答。
在应答过程中,前者需要Th细胞的辅助,后者不需要。
第一节B细胞对TD抗原的免疫应答一、B细胞对TD抗原的识别BCR是B细胞识别特异性抗原的受体。
BCR识别抗原对B细胞的激活有两个相关联的作用:BCR特异性结合抗原,产生B细胞活化的第一信号;B细胞内化BCR所结合的抗原,并对抗原进行加工,形成抗原肽-MHCⅡ类分子复合物(peptide-MHCⅡcomplex,pMHC),提呈给抗原特异性Th识别,Th活化后通过表达的CD40L与B细胞表面的CD40结合,又提供B细胞活化的第二信号。
BCR对抗原的识别与TCR识别抗原不同:①BCR不仅能识别蛋白质抗原,还能识别多肽、核酸、多糖类、脂类和小分子化合物类抗原;②BCR能特异性识别完整抗原的天然构象,或识别抗原降解所暴露表位的空间构象;③BCR对抗原的识别不需APC的加工和提呈,亦无MHC限制性。
二、B细胞活化需要的信号与T细胞相似,B细胞活化也需要双信号:特异性抗原提供第一信号启动B细胞活化,而共刺激分子提供的第二信号使B细胞完全活化。
B细胞活化后的信号转导途径也与T细胞相似。
(一)B细胞活化的第一信号B细胞活化的第一信号又称抗原刺激信号,由BCR-CD79a/b (BCR-Igα/β)和CD19/CD21/CD81共同传递。
1. BCR-CD79a/b信号BCR与抗原特异性结合后即启动B细胞活化的第一信号。
但由于BCR重链胞质区短,自身不能传递信号,故需经BCR复合物中的CD79a/b将信号转入B细胞内,与CD3类似,CD79a/b胞质区亦存在ITAM。
【免疫学总结】B淋巴细胞介导的体液免疫应答

【免疫学总结】B淋巴细胞介导的体液免疫应答B淋巴细胞介导的体液免疫应答外来抗原进入机体后诱导抗原特异性B细胞活化、增殖,并最终分化为浆细胞,产生特异性抗体,存在于体液中,发挥重要的免疫效应作用,此过程称为特异性体液免疫应答第一节 B细胞对TD抗原的免疫应答一、B细胞对TD抗原的识别特点①不仅能识别蛋白质抗原,还能识别多肽、核酸、多糖类、脂类、小分子化合物②可特异性识别完整抗原的天然构象或识别抗原降解所暴露的表位的空间构象③识别抗原无需经APC的加工处理,无MHC限制性二、B细胞活化需要的信号(1)第一信号1.第一活化信号经由Ig/αIgβ传导入胞内BCR被多价抗原交联后,ITAM模体中酪氨酸磷酸化,募集并活化Syk,活化细胞内信号转导的级联反应,经PKC、MAPK、钙调蛋白三条途径激活转录因子,参与并调控B细胞激活、增殖相互基因的表达2.B细胞活化中共受体的作用成熟B细胞表面,CD19、21、81非共价键结合成共受体复合物,提高敏感性(2)第二信号主要由黏附分子对的相互作用所提供,最重要的是CD40/CD40L(3)TB细胞相互作用与B细胞免疫应答一方面,B细胞可作为抗原提呈细胞活化T细胞,另一方面活化的T细胞可以提供B细胞活化的第二信号,并分泌多种IL-4等细胞因子协助B细胞的进一步分化三、B细胞增殖和终末分化需Th细胞的辅助,发生于外周淋巴器官的T细胞区和生发中心四、B细胞在生发中心的分化成熟在外周淋巴器官的T细胞区激活的部分B细胞进入初级淋巴小结,分裂增殖,形成生发中心,分裂增殖的B细胞称为生发中心母细胞,母细胞分裂增殖产生的子代细胞体积小,称为生发中心细胞生发中心分为两个区域:明区、暗区,前者FDC(滤泡树突状细胞)较多,后者生发中心母细胞紧密聚集DC的树突表面高表达CD21分子,抗原-抗体复合物通过C3d 与CD21分子结合,附着在FDC树突上,或结合于FDC树突上的Fc受体,聚集在一起,呈串珠状,称串珠样小体生发中心四个事件(1)体细胞高频突变和Ig亲和力成熟生发中心母C的轻链和重链V基因,可发生高频率的点突变,称体细胞高频突变(2)Ig的类别转换可变区相同而Ig类别变化的过程称Ig的类别转换,或称同种型转换,遗传学基础是同一V区基因与不同重链C基因的重排(3)浆细胞的形成浆细胞是B细胞分化的终末细胞,除了少量线粒体,内含大量粗面内质网,分泌抗体,BCR表达少(4)记忆B细胞的产生特异性表面标志:CD27,不产生抗体,再次遇同一抗原可迅速活化大量产生特异Ig第二节 B细胞对TI抗原的免疫应答某些抗原,如细菌多糖、多聚蛋白质及脂多糖等属TI抗原,能激活初始B细胞而无需Th细胞的辅助㈠B细胞对TI-1抗原发生的应答TI-1抗原又称B细胞丝裂原,成熟或不成熟的B细胞均可被TI-1抗原激活,诱导产生低亲和力的IgM,无记忆性㈡B细胞对TI-2抗原发生的应答TI-2仅能激活成熟B细胞,主要是B-1细胞TI-2抗原通过高度重复的抗原表位使B细胞的mIg广泛交联而被激活,(但过度交联使成熟B细胞产生耐受)可直接激活B-1细胞,T细胞分泌的细胞因子可明显增强此类B细胞的免疫应答,并发生抗体类型转换,可产生IgM IgG第三节体液免疫应答抗体产生的一般规律特定抗原初次刺激机体所引发的应答称为初次应答;初次应答中形成的记忆淋巴细胞再次接触相同抗原刺激后,可迅速、高效、持久的应答,即再次应答或回忆应答㈠初次应答特点为潜伏期长,抗体水平低,亲和力低,抗体升高所需时间长,抗体主要为IgM㈡再次应答①潜伏期短②抗体浓度增加快③抗体维持时间长④诱发再次应答所需抗原剂量小⑤再次应答产生高亲和力抗体IgG再次应答的强弱取决于两次抗原刺激的间隔长短,过长或过短反应均弱。
大连医科大学免疫学PPT人卫7版13B细胞介导的特异性免疫应答
(1)B1细胞 : CD5+ TI 抗原 IgM, SIgA
(2) B2细胞: CD5TD抗原 IgG
TI-1和TI-2抗原
(一) B细胞对TI-1抗原的应答
TI-1抗原
12
B1 细胞
(1)BCR (2)丝裂原受体
(二)B细胞对TI-2抗原的应答
高度重复的表位,B1 细胞的mIg广泛交联, 激活B1细胞。
(三)细胞因子的作用
Y
B
信号 1
信号 2
Th
细胞因子 IL-4 IL-5 IL-6 IFN-γ
TGF-β
B
B
B
B
BB B
PC
B
B
B BB B
B
增殖和分化
Th细胞分泌细胞因子的作用
(四)T、B细胞的相互作用
(四)T、B细胞的相互作用
(四)T、B细胞的相互作用
(六)浆细胞的形成
(七)记忆B细胞(Memory B Cells, Bm)的产生
一、初次应答
所需抗原量多、抗体潜伏期长、抗体效价低、 维持时间短、与抗原亲和力低、以IgM为主;
二、再次应答
所需抗原量少、抗体潜伏期短、抗体效价高、 维持时间长、与抗原亲和力高、以IgG为主。
第四节 B细胞介导的体液免疫应答效应
1 .调理作用 2 .补体激活 3. ADCC 4. 中和毒素
Figure 1-24 part 2 of 3
TI-2 抗原
1
B1 细胞
TI 抗原诱导的体液免疫应答
初次应答
再次应答
抗 100000
原
特 10000
异
性 1000
抗
13 B淋巴细胞介导的特异性免疫应答
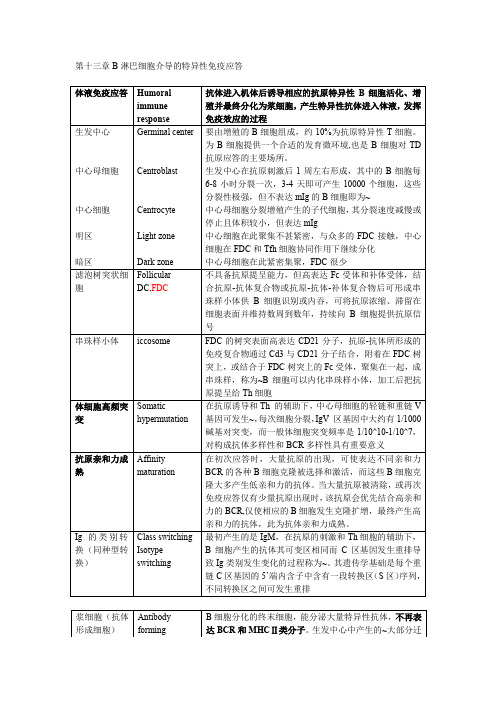
第十三章B淋巴细胞介导的特异性免疫应答体液免疫应答Humoralimmuneresponse 抗体进入机体后诱导相应的抗原特异性B细胞活化、增殖并最终分化为浆细胞,产生特异性抗体进入体液,发挥免疫效应的过程生发中心中心母细胞中心细胞明区暗区Germinal centerCentroblastCentrocyteLight zoneDark zone要由增殖的B细胞组成,约10%为抗原特异性T细胞。
为B细胞提供一个合适的发育微环境,也是B细胞对TD抗原应答的主要场所。
生发中心在抗原刺激后1周左右形成,其中的B细胞每6-8小时分裂一次,3-4天即可产生10000个细胞,这些分裂性极强,但不表达mIg的B细胞即为~中心母细胞分裂增殖产生的子代细胞,其分裂速度减慢或停止且体积较小,但表达mIg中心细胞在此聚集不甚紧密,与众多的FDC接触,中心细胞在FDC和Tfh细胞协同作用下继续分化中心母细胞在此紧密集聚,FDC很少滤泡树突状细胞FollicularDC,FDC不具备抗原提呈能力,但高表达Fc受体和补体受体,结合抗原-抗体复合物或抗原-抗体-补体复合物后可形成串珠样小体供B细胞识别或内吞,可将抗原浓缩、滞留在细胞表面并维持数周到数年,持续向B细胞提供抗原信号串珠样小体iccosome FDC的树突表面高表达CD21分子,抗原-抗体所形成的免疫复合物通过Cd3与CD21分子结合,附着在FDC树突上,或结合于FDC树突上的Fc受体,聚集在一起,成串珠样,称为~B细胞可以内化串珠样小体,加工后把抗原提呈给Th细胞体细胞高频突变Somatichypermutation在抗原诱导和Th 的辅助下,中心母细胞的轻链和重链V基因可发生~,每次细胞分裂,IgV 区基因中大约有1/1000碱基对突变,而一般体细胞突变频率是1/10^10-1/10^7,对构成抗体多样性和BCR多样性具有重要意义抗原亲和力成熟Affinitymaturation在初次应答时,大量抗原的出现,可使表达不同亲和力BCR的各种B细胞克隆被选择和激活,而这些B细胞克隆大多产生低亲和力的抗体。
医学免疫学13章体液免疫应答精品PPT课件

D V J Cm1 Cm2 Cm3 Cm4
AAA Cd1
Cd2 Cd3
RNA cleaved and polyadenylated at pA1
V D J Cm IgM mRNA
D V J Cm1 Cm2 Cm3 Cm4
Cd1
pA1
Cd2 Cd3
V D J Cd IgD mRNA
RNA cleaved and polyadenylated at pA2
TI-1 antigens are polyclonal B cell activators at high concentrations, whereas at low concenrations they
induced an antigen-specific antibody response
二、B细胞活化、增殖、分化阶段 (一)B细胞活化信号要求(双信号)
第一活化信号:BCR/共受体+Ag
由Igα/Igβ传入胞内
共受体CD19/CD21/CD81
第二活化信号:CD40+CD40L(活化 Th) CKs:Th2释放的CKs辅助B活化增殖分化
共受体
(CD19/CD21/CD81)
第二信号:CD40---CD40L
AAA
Molecular mechanisms for isotype switching of heavy chain
分化为浆细胞,产生Ab; 形成Bm(记忆性B细胞)
三、效应阶段(抗体的效应)
中和作用:virus,exotoxin 调理作用:Ab,C 激活补体:MAC ADCC效应:NK,Nue,Mψ
MHC-II类限制性
BCR信号的转导途径
13章体液免疫

第二节 B细胞对TI抗原的免疫应答 第三节 体液免疫应答抗体产生的一般规律
3
第一节 B细胞对TD抗原的免疫应答
基本过程:三个阶段
B细胞特异性识别Ag阶段 B细胞活化、增殖、分化阶段 效应分子抗体的产生及效应阶段
4
第一节 B细胞对TD抗原的免疫应答
两个去向 一部分B细胞到淋巴结髓质,增殖分化为短寿命
的浆细胞产生IgM抗体; 另一部分B细胞到初级淋巴小结继续增殖分化形
成生发中心。
19
B细胞在生发中心的分化成熟
20
3 2
生发中心细胞
生发中心母细胞 1
21
B细胞在生发中心要完成 Ig亲和力成熟和体细胞高频突变 Ig的类别转换 浆细胞的形成 记忆B细胞的产生
B细胞对TD抗原的识别
BCR直接识别抗原表位
识别特点:识别多种抗原
无需APC提呈 不受MHC限制
5
B细胞的活化、增殖和分化
B细胞活化 双信号
BCR与特异性抗原(表位)结合,由 第一信号 Igα/Igβ传入胞内 第二信号 活化Th表达CD40L与B细胞表面CD40结合
6
TD-Ag
1
2
7
B细胞活化的双信号模型
35
(二)B细胞对TI-2抗原的免疫应答
TI-2抗原
BCR
BCR适度交联
B1细胞
信号转导
36
第三节 体液免疫应答抗体产生的一般规律 抗体的产生一般分为4个时期
Th、B细胞相互作用
Th 细胞活化、增殖
未成熟DC 淋巴液/血液
外周免疫器官
(T细胞依赖区)
TD-Ag
第十三章B淋巴细胞介导的体液免疫应答

B淋巴细胞介导的体液免疫应答2/21/2022青海大学医学院免疫学教研室1B淋巴细胞介导的体液免疫应答⏹⏹⏹概念特异性体液免疫应答◆B细胞对TD抗原的免疫应答一、B细胞对TD-Ag的识别特点➢➢➢一、B 细胞对TD-Ag 的识别二、B细胞活化需要的信号⏹⏹PLC-γ磷酸化胞内Ca 2+浓度增高钙调磷酸酶PKC Crb-2活化Ras-GDPRas-GTP MAP激酶活化甘油二酯(DAG)NFAT AP-1NF-κBBCR复合体介导的胞内信号转导P P PBCR CD79a CD79b C3d CD21CD19CD81抗原相关激酶激活下游激酶B细胞共受体在B细胞活化中的作用加强BCR 复合物转导的信号(二)B细胞活化的第二信号(三)T、B细胞相互作用与B细胞免疫应答✓✓步✓三、B细胞的增殖和终末分化四、B细胞在生发中心的分化成熟⏹⏹⏹⏹⏹B 记忆细胞暗区明区生发中心树突细胞初级淋巴小结早期生发中心生发中心B细胞在生发中心的分化成熟(一)体细胞高频突变和Ig亲和力成熟,称之抗体亲和力成熟★(二)Ig 的类别转换(class switch )概念重组细胞因子B 细胞合成抗体的类别转换和亲和力成熟IL-4IL-2IL-4IL-5IL-2IL-4IgM IgEIgA or IgG 2b IFN-gTGF-βIgG2aor IgG3活化B 细胞浆细胞Ig 类别转换(三)浆细胞(plasma cell)(四)记忆性B细胞(memory B cell)第二节B细胞对TI-Ag的免疫应答TI-1抗原诱导B细胞的激活(1)* * *★TD 、TI 抗原的比较TD 抗原TI 抗原抗原组成细胞性,蛋白质主要为脂多糖、多糖结构特点结构复杂,T表位+B表位,有载体决定簇结构简单,多重复B表位,无载体决定簇Mφ等APC参与需要不需要T细胞辅助需要不需要免疫应答类型体液+细胞免疫体液免疫产生Ig类型IgG等各类型Ig只有IgM抗体亲和力成熟有无Bm形成,记忆性有,再次应答无,只有初次应答抗体发生时期迟早诱导免疫耐受难易抗原实例白喉毒素、病毒血凝素、PPD等百日咳等G—菌胞壁脂多糖,荚膜多糖、SPA第三节体液免疫应答抗体产生的一般规律٭初次应答(Primary response):٭再次应答(Secondary response):特性初次应答二次应答抗原提呈非B细胞B细胞等抗原浓度高低抗体产生潜伏期长5—10天短2—5天抗体浓度上升慢快抗体高峰浓度低,维持短高,维持长抗体浓度下降快,等臂对称慢,不对称抗体类型主要为IgM主要为IgG(IgA)抗体亲和力低高无关抗体多少抗体产生的一般规律:★B细胞应答的效应中和作用免疫调理作用激活补体抗体依赖性细胞介导的细胞毒作用(ADCC)分泌型IgA的局部抗感染作用免疫损伤作用小结免疫应答的特点小结1.B细胞对TD、T I-1及T I-2抗原免疫应答的异同;2.Th细胞辅助B细胞应答的机制;3.初次应答与再次应答的概念与特点;4.抗体介导的免疫效应。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
TI-2抗原诱导B细胞的激活
第三节 体液免疫应答抗体产生的 一般规律
(一)初次应答
潜伏期:抗原刺激后至血清中能测到特异 抗体前的阶段; 对数期:抗体量呈指数增长; 平台期:血清中抗体浓度基本维持在一个 相当稳定的较高水平; 下降期:血清中抗体浓度; ②诱发再次应答所需抗原剂量小; ③抗体浓度增加快,快速到达平台期,平台高; ④抗体维持时间长; ⑤再次应答主要产生高亲和力的抗体IgG, 而初次应答中主要产生低亲和力的IgM。
第十三章
B淋巴细胞介导的体液 免疫应答
第一节 B细胞对TD抗原的免疫应答
一、B细胞对TD抗原的识别
BCR特异性结合抗原,产生B细胞活化的第一 信号;B细胞内化与其BCR结合的抗原,并进行 加工处理,形成抗原肽-MHCⅡ类分子复合物, 提呈给抗原特异性Th细胞识别。
BCR识别的抗原无需经APC的加工和处理,也 无MHC限制性。
Igα/Igβ胞质区ITAM磷酸化
SyK被募集并活化 PLC-γ活化途径 MAP激酶活化途径
转录因子活化
相关基因转录
三、B细胞的增殖和终末分化
T、B细胞相互作用
Th细胞辅助B细胞免疫应答 ①Th细胞提供B细胞活化的第二信号 (CD40/CD40L); ②Th细胞分泌多种细胞因子协助B细胞分化 (Th1 IL-2、IFN-γ、Th2 IL-4、5、6) B细胞作为APC活化T细胞 ①B细胞提供T细胞活化的第一信号(抗原肽MHCⅡ类分子复合物) ②活化的B细胞提供T细胞活化的第二信号 (B7/CD28)
(三)浆细胞的形成
•又称抗体形成细胞,能合成和分泌特异性抗体; •大部分迁入骨髓,在较长时间内持续产生抗体; •表面不再表达BCR和MHCⅡ类分子,不能再与抗 原起反应,也失去了与Th相互作用的能力;
(四)记忆性B细胞的产生
•生发中心中存活下来的B细胞,或分化发育 成浆细胞,或成为记忆性细胞离开生发中心 进入血液参与再循环; •记忆性B细胞不产生Ig,但再次与同一抗原 相遇时可迅速活化,产生大量抗原特异的Ig。
(二)Ig的类别转换
•在免疫应答中首先分泌IgM,随后可表达IgG、 IgA或IgE,而其IgV区不发生改变,这种可变区 相同而Ig类别发生变化的过程称为Ig的类别转换 或同种型转换; •类别转换的遗传学基础是同一V区基因与不同重 链C基因的重排; •Ig的类别转换在抗原诱导下发生,接受Th细胞 分泌的多种细胞因子的调节。
•生发中心母细胞的轻链和重链V基因可发生高频率 的点突变,称为体细胞高频突变; •在每次细胞分裂中,IgV区基因中大约每1000个 bp中就有一对发生突变; •体细胞高频突变在抗原诱导下发生; •再次免疫应答时抗原会优先结合高亲和力的BCR, 产生高亲和力的抗体,称为抗体亲和力成熟; •导致BCR多样性及体液免疫应答中抗体的多样性。
二、B细胞活化需要的信号
B细胞活化的第一信号:BCR特异性结合抗 原启动第一信号,经由Igα/Igβ传导入胞内, B细胞共受体加强信号的传导; B细胞活化的第二信号(协同刺激信号) CD40/CD40L(活化的Th细胞)
B细胞活化的信号转导途径
BCR交联
活化酪氨酸激酶(Blk、Fyn或Lyn)
Th细胞对B细胞的辅助作用发生于外 周淋巴器官的T细胞区和生发中心。
四、B细胞在生发中心的分化成熟
生发中心中绝大多数B细胞发生凋亡,部分B
细胞在抗原刺激和T细胞辅助下继续分化发
育,在生发中心中完成体细胞高频突变和Ig
亲和力成熟、Ig类别转换,最终形成浆细胞
和记忆性B细胞。
(一)体细胞高频突变和Ig亲和力成熟
五、B细胞的效应
抗体的效应 记忆性B细胞的效应
第二节 B细胞对TI抗原的免疫应答
TI抗原分为TI-1和TI-2抗原两类。 TI-1抗原:又被称为B细胞丝裂原,如LPS,成熟或 不成熟的B细胞均可被激活。高浓度时经丝裂原受体 与B细胞结合诱导多克隆B细胞增殖和分化;低浓度时 激活抗原特异性B细胞。激活初始B细胞无需Th细胞辅 助,诱导产生低亲和力的IgM,单独不足以诱导Ig类 别转换、抗体亲和力成熟及记忆性B细胞形成。 TI-2抗原:多为细菌胞壁与荚膜多糖,具有高度重复 的结构,仅能激活成熟的B细胞,应答的主要是B-1细 胞。通过其高度重复的抗原表位使B细胞的mIg广泛交 联而被激活,但过度交联会使成熟B细胞出现耐受。