cyclin review
甲状腺癌患者Cyclin D1及Ki-67的临床意义研究

88例;年龄15 ~78岁,平均(45.3 ±8.2)岁。对照 结转移的患者相比,有淋巴结转移的患者具有更高的
组中男性38例,女性84例;年龄20 ~74岁,平均 Cyclin D1表达阳性率和较低的Ki -67表达阳性率
(47.1 ±8.7)岁。2组患者间的性别和年龄差异均 (P <0.05),见表 1°
80
( ) 64 80.00
( ) 16 20.00
69
( ) 56 81.16
( ) 13 18.84
62
( ) 39 62.90
( ) 23 37.10
2. 531 0. 137 8. 195 0. 3644 2. 202 5. 771 5. 461
>0.05 >0.05 <0.05 >0.05 >0.05 <0.05 <0.05
] [关键词 细胞周期蛋白;增殖细胞核抗原;甲状腺癌;病理特征;预后 [ ] ] 中图分类号 R581 [文献标识码 A
Clinical significance of Cyclin DI and Ki 一 67 in patients with thyroid carcinoma WANG Ying' , HAN Enshan2 ,XIA Xiao . 1. Wuzhong Peoples Hospital,Wuzhong 751100, China*;. Department of Pathology,General Hospital of Ningxia Medical University,Yinchuan 750004, China [Abstract] Objective To investigate the expression of cylin D1 and Ki — 67 in patients with thyroid cancer and their correlation with the pathological characteristics and prognosis. Methods 131 paraffin specimens of TC tissues and adjacent normal tissues were col lected ,and the clinical pathological data of the patients were collected. The positive expression of cylin D1 and Ki — 67 were detected by immunohistochemistry and the correlation with clinicopathological features and prognosis of the patients was analyzed. Results In thyroid carcinoma, the expressions of Cyclin D1 and Ki — 67 were not statistically different in gender, age, histopathological type, and capsular in vasion (P > 0. 05 ). However,the positive rates of Cyclin D1 and Ki — 67 were significantly correlated with tumor diameter, pathological stage and lymph node metastasis ( P < 0. 05),and their expression was significantly correlated with five — year survival rate of patients. Conclusion Cyclin D1 and Ki — 67 are related to the pathological characteristics and poor prognosis of patients with thyroid carcinoma, and have the potential to become biomarkers for clinical diagnosis and prognosis evaluation of thyroid carcinoma. [Key words ] Cyclin D1 ; Ki — 67; Thyroid cancer; Pathological features ; Prognosis
the review of scientific instruments

The Review of Scientific InstrumentsIntroductionThe field of scientific research heavily relies on the use of various instruments and tools to conduct experiments, collect data, and analyze results. The review of scientific instruments plays a crucial role in ensuring the accuracy, reliability, and efficiency of scientific studies. This article aims to explore the importance of instrument reviews and provide an in-depth analysis of their significance in scientific research.The Role of Instrument ReviewsInstrument reviews serve as an essential step in the scientific research process, as they enable scientists to select the most suitable tools for their experiments. By reviewing scientific instruments, researchers can assess their performance, quality, and suitability for a specific study. This evaluation process allows scientists to make informed decisions regarding instrument acquisition and usage, ensuring reliable and accurate scientific results.Factors to Consider in Instrument ReviewsWhen conducting instrument reviews, several factors should be taken into account. These factors include:1. Accuracy and PrecisionOne of the primary considerations in instrument reviews is the accuracy and precision of the measurement or analysis it provides. A reliable instrument should yield consistent and precise results, minimizingerrors and uncertainties in scientific experiments.2. Sensitivity and Detection LimitsThe sensitivity of an instrument refers to its ability to detect small changes or variations in the measured parameters. Instrument reviews should focus on the sensitivity of the tools, as this determines their capability to detect subtle changes in experimental conditions. Additionally, the detection limits of instruments should also be considered to ensure they can measure even the lowest concentrations or values accurately.3. Durability and LongevityScientific instruments can be a significant investment, makingdurability and longevity crucial factors in instrument reviews. Researchers need tools that can withstand rigorous usage and maintain their performance over an extended period. Instrument reviews should assess the robustness and reliability of instruments to ensure they can withstand the demands of scientific experiments.4. User-Friendliness and Ease of OperationInstrument reviews should also evaluate the user-friendliness and ease of operation of scientific tools. Researchers need instruments that are intuitive to use, with clear instructions and user interfaces. When instruments are user-friendly, scientists can save time and effort in conducting experiments and data analysis.Importance of Instrument Reviews in Scientific ResearchInstrument reviews play a vital role in scientific research for several reasons:1. Quality AssuranceThrough instrument reviews, researchers can ensure the quality and reliability of the data generated from scientific experiments. These reviews help identify any limitations or shortcomings of instruments and prevent inaccurate or misleading results.2. Cost-EffectivenessSelecting the most appropriate instruments for scientific studies is essential to optimize costs. Instrument reviews allow researchers to compare different options and choose tools that provide the necessary performance at the most cost-effective price.3. ReproducibilityReproducibility is an essential aspect of scientific research, ensuring that experiments and results can be independently verified and validated. By reviewing instruments, scientists can increase the reproducibility of their experiments, as other researchers can acquire the same tools and achieve comparable results.4. Innovation and AdvancementInstrument reviews contribute to the progress and innovation inscientific research. By evaluating existing instruments, scientists can identify areas for improvement and develop advanced tools with enhanced capabilities. This continuous cycle of instrument reviews and innovation drives scientific discoveries and technological advancements.ConclusionThe review of scientific instruments is a critical step in thescientific research process. It helps scientists select the mostsuitable tools, ensures the quality and reliability of data, and contributes to the advancement of scientific knowledge. By considering factors such as accuracy, sensitivity, durability, and user-friendliness, researchers can make informed decisions and conduct experiments with confidence. Additionally, instrument reviews promote cost-effective approaches, reproducibility, and innovation, ultimately leading to groundbreaking discoveries and advancements in various scientific fields.。
男性尿道原发性弥漫大B细胞淋巴瘤1例报告
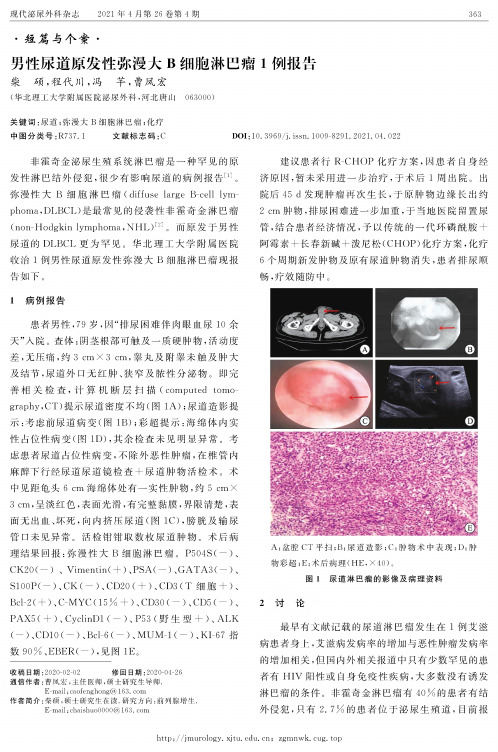
现代泌尿外科杂志2021年4月第26卷第4期363•短篇与个G •男性尿道原发性弥漫大B 细胞淋巴瘤1例报告柴硕,程代川,冯芋,曹凤宏(华北理工大学附属医院泌尿外科,河北唐山063000)关键词:尿道;弥漫大B 细胞淋巴瘤;化疗中图分类号:R737. 1 文献标志码:C DOI :10. 3969/j. issn. 10098291. 2021. 04. 022非霍奇金泌尿生殖系统淋巴瘤是一种罕见的原 发性淋巴结外侵犯,很少有影响尿道的病例报告「门。
弥漫性大B 细胞淋巴瘤(diffuse large B-cell lym phoma, DLBCL ) 是最常见的侵袭性非霍奇金淋巴瘤 (non-Hodgkin lymphoma ,NHL )「2*。
而原发于男性 尿道的DLBCL更为罕见。
华北理工大学附属医院 收治1例男性尿道原发性弥漫大B 细胞淋巴瘤现报 告如下。
1病例报告患者男性,79岁,因“排尿困难伴肉眼血尿10余 天”入院。
查体:阴茎根部可触及一质硬肿物,活动度 差,无压痛,约3 cmX 3 cm,睾丸及附睾未触及肿大 及结节,尿道外口无红肿、狭窄及脓性分泌物。
即完 善相关检查,计算机断层扫描(c omputed tomo graphy, CT) 提示尿道密度不均(图1A);尿道造影提 示:考虑前尿道病变(图1B);彩超提示:海绵体内实 性占位性病变(图1D),其余检查未见明显异常。
考 虑患者尿道占位性病变,不除外恶性肿瘤,在椎管内 麻醉下行经尿道尿道镜检查+尿道肿物活检术。
术 中见距龟头6 cm 海绵体处有一实性肿物,约5 cmX3 cm,呈淡红色,表面光滑,有完整黏膜,界限清楚,表 面无出血、坏死,向内挤压尿道(图1C),膀胱及输尿 管口未见异常。
活检钳钳取数枚尿道肿物。
术后病 理结果回报:弥漫性大B 细胞淋巴瘤。
P504S (—)、CK20( — )、Vimentin ( + )、PSA ( — )、GATA3(—)、S100P ( — )、CK ( — )、CD20 ( + )、CD3 ( T 细胞 + )、Bcl2( + )、C-MYC(15% + )、CD30( — )、CD5(—)、PAX5 ( +)、C yclinD1 ( — )、P 53 (野生型 + )、A LK(—)、CD10 ( — )、Bcl-6 ( — )、MUM-1 ( — )、KI-67指 数 90%、EBER (—),见图 1E 。
circRNA在心血管疾病中的研究进展

[5]朱志悦.脑梗死后吞咽功能障碍的治疗及护理现状[J ].中国城乡企业卫生,2020,35(2):34-37.[6]曲淑芹,李英.延续性护理对老年脑梗死后遗症的效果分析[J ].中国继续医学教育,2020,12(15):194-196.[7]O K A B E Y O Y ,T A K E U C H I K T K ,I Z UM I M I M ,e t a l .P o s t e r i o rt e e t h o c c l u s i o n a n d d y s p h a g i a r i s k i n o l d e r n u r s i n g ho m e r e s i -d e n t s :a c r o s s -s e c t i o n a l o b s e r v a t i o n a l s t u d y [J ].J O r a l R e h a b i l i t ,2017,2(44):89-95.[8]S E R R A N O S A N T O S J M ,P O L A N D F ,WR I G H T D ,e t a l .M e d i -c i n e s a d m i n i s t r a t i o n f o r r e s i d e n t s w i t h d y s p h a gi a i n c a r e h o m e s :a s m a l l s c a l e o b s e r v a t i o n a l s t u d y t o i m pr o v e p r a c t i c e [J ].I n t J P h a r m ,2016,512(2):416-421.[9]P A R K Y ,B A N G H L ,H A N H ,e t a l .D y s p h a g i a s c r e e n i n g me a s -u r e sf o r u s e i n n u r s i ngh o m e s :a s ys t e m a t i c r e v i e w [J ].J K o r e a A c a d N u r s ,2015,45(1):1.[10]刘丽爽,张慧,郭亚娟,等.家庭延续性护理对脑卒中吞咽障碍患者居家康复的影响[J ].心血管外科杂志,2019,8(3):207.[11]黎巧玲,黄朝军,胡晓红,等.脑卒中吞咽障碍患者居家护理模式及效果评价[J ].中国康复,2019,34(6):291-294.[12]余红雨,钱荣,杨帆,等.微信平台在脑卒中患者延续性护理中的应用[J ].牡丹江医学院学报,2019,40(4):142-143.[13]汪薇.认知行为干预对脑卒中吞咽障碍患者进食体位依从性及误吸发生率的影响[J ].国际感染杂志,2018,7(4):175-177.[14]刘先娟,郭晓娟,黄天然,等.出院电话随访式延续护理对脑卒中患者日常生活能力的影响[J ].现代医药卫生,2019,35(15):2374-2376.[15]王元姣,吴金香.家庭延续性护理对脑卒中吞咽障碍患者居家康复的影响[J ].护士进修杂志,2017,32(18):1698-1700.[16]G O U V E I A B R ,J A R D I M H G ,MA R T I N S M M ,e t a l .A n e v a l u -a t i o n o f a n u r s e -l e d r e h a b i l i t a t i o n p r o gr a mm e (t h e P r o b a l a n c e p r o g r a mm e )t o i m pr o v e b a l a n c e a n d r e d u c e f a l l r i s k o f c o mm u n i -t y -d w e l l i n g o l d e r p e o pl e :a r a n d o m i s e d c o n t r o l l e d t r i a l [J ].I n t J N u r s S t u d y,2016,56:1-8.[17]刘玉昌,刘秀芝,金雷,等.社区康复护理对促进脑卒中患者功能康复作用分析[J ].上海医药,2016,37(22):58-59.[18]邓宪勇,余永恒.家庭病床服务模式在脑卒中康复期患者社区康复中的应用[J ].海南医学,2016,27(15):2556-2557.[19]颜志刚,何莉娜,李银娇,等.脑梗死合并吞咽障碍社区综合管理研究[J ].按摩与康复医学,2018,9(19):79-81.[20]高丽,刘均娥,崔允文,等.时间护理在护理工作中的应用现状[J ].护理管理杂志,2011,11(1):36-37.[21]胡娜,唐晓红,黄小丽.时间康复护理方案在促进脑梗死后吞咽障碍功能恢复中的应用研究[J /C D ].实用临床护理学电子杂志,2019,4(39):155.[22]张春妮,赵飞丽.时间康复护理方案对脑梗死后吞咽障碍患者功能康复的影响[J ].实用临床医药杂志,2016,20(6):10-12.[23]薛琴芬,王艳玲,薛玲玲.五行音乐疗法对结直肠癌手术患者负性情绪㊁睡眠质量及生存质量的影响研究[J ].检验医学与临床,2020,17(12):1763-1765.[24]冯淑芬,黄凤英,秦秀英.音乐疗法对急性脑梗死认知功能障碍患者的效果研究[J ].中国现代药物应用,2019,13(23):227-229.[25]郜彦鑫,王帅,王秋月.‘黄帝内经“中五行音乐疗法治疗抑郁症的研究进展[J ].山东化工,2019,48(5):69-70.[26]唐钦妹,施素华,黄向红,等.五行音乐疗法对MH D 初期患者焦虑状况的干预效果[J ].承德医学院学报,2016,33(6):482-484.[27]B A H I A M M ,MO U R ÀO L F ,C HU N R Y S .D ys a r t h r i a a s a p r e -d i c t o r o f d y s p h a g i a f o l l o w i n g s t r o k e [J ].N e u r o R e h a b i l i t ,2016,38(2):155-162.[28]安德连,陈妙霞,陈琼梅,等.吞咽障碍护理门诊的构建[J ].中华护理杂志,2017,52(2):219-221.[29]安德连,窦祖林,李慧娟,等.吞咽障碍护理门诊对卒中患者康复效果的影响[J /C D ].实用临床护理学电子杂志,2018,3(6):16-17.[30]庄红花,尹玉珊,米红艳.中风护理门诊与电话随访相结合在吞咽障碍患者延续性护理中的应用效果[J ].国际护理学杂志,2017,36(9):1186-1188.(收稿日期:2020-10-06 修回日期:2021-03-04)ә通信作者,E -m a i l :c h e n y u n q i n g.88@163.c o m ㊂㊃综 述㊃c i r c R N A 在心血管疾病中的研究进展李格格综述,陈运清ә审校(重庆医科大学附属第二医院心血管内科,重庆400010) [摘 要] 环状R N A (c i r c R N A )是一类内源性非编码R N A ,广泛存在于全血㊁血浆和细胞外小泡中,因具有共价闭合环状结构而不易被核酸外切酶降解,具有丰度高㊁稳定性好㊁保守性强及组织或发育特异的表达模式等特点㊂c i r c R N A 通过吸附m i R N A 等多种机制来调节基因表达㊂该文介绍了c i r c R N A 在多种心血管疾病中的研究进展,为c i r c R N A 在心血管疾病中发挥诊断和治疗作用提供了新的科学依据㊂[关键词] 环状R N A ; 微小R N A ; 心血管疾病D O I :10.3969/j.i s s n .1009-5519.2021.11.022中图法分类号:R 541文章编号:1009-5519(2021)11-1878-03文献标识码:A心血管疾病包括冠心病㊁心脏瓣膜病㊁心肌病㊁心律失常㊁心力衰竭等,在中国是导致死亡和过早死亡的主要病因,占中国人死亡原因的40%[1],所以对于心血管疾病的早期诊断和积极治疗是很有必要的㊂环状R N A (c i r c R N A )是前体m R N A 成熟过程中形成的一类丰富的内源R N A ,含有一个共价闭合的连续环㊂c i r c R N A 在真核细胞中广泛表达,在进化上是保守的,并且可以针对特定的细胞类型或发育阶段㊂此外,在细胞核和线粒体中也发现了c i r c R N A ㊂与线性R N A 不同,c i r c R N A 没有5'帽或3'尾结构,不易被细㊃8781㊃现代医药卫生2021年6月第37卷第11期 J M o d M e d H e a l t h ,J u n e 2021,V o l .37,N o .11胞中的核酸外切酶降解[2]㊂c i r c R N A可以调节基因表达,参与多种病理生理过程㊂目前,许多研究已经证实c i r c R N A与动脉粥样硬化㊁冠心病㊁心肌病㊁心房颤动㊁心力衰竭㊁瓣膜钙化等相关,且有望被用于疾病的治疗㊂1c i r c R N A的生物合成及作用机制c i r c R N A是由线性前体R N A通过剪接体机制介导的反向剪接环化而产生的,主要有以下2种环化方式:内含子配对介导的环化和套索介导的环化㊂由此产生了3种类型的c i r c R N A,分别是外显子c i r-c R N A㊁内含子c i r c R N A㊁外显子-内含子c i r c R N A㊂绝大多数c i r c R N A存在于细胞质中,少数存在于细胞核中㊂目前,有研究认为c i r c R N A主要通过以下4种途径参与调节病理生理过程:(1)c i r c R N A作为m i R-N A海绵来调节m i R N A的功能;(2)c i r c R N A与R B-P s(R N A结合蛋白)相互作用调节相关蛋白的功能;(3)含有内含子序列的c i r c R N A通过与U1s n R N P (剪切子)结合形成的复合物与R N A聚合酶Ⅱ相互作用来调节亲本基因的表达;(4)作为反向剪接的结果而存在的开放阅读框(O R F)与内部核糖体进入位点(I R E S)或m6A-修饰结合在一起诱导c i r c R N A进行翻译[3]㊂2c i r c R N A与心血管疾病2.1c i r c R N A与动脉粥样硬化动脉粥样硬化被认为是一种退行性病变,目前被认为是多因素(包括血脂异常㊁高血压㊁糖尿病㊁吸烟㊁遗传因素㊁体力活动减少㊁年龄和性别㊁酒精摄入㊁肥胖等)共同作用的结果,首先是血管平滑肌细胞㊁巨噬细胞及T淋巴细胞聚集;其次是胶原㊁弹力纤维及蛋白多糖等结缔组织基质的增生;再者是脂质积聚,其主要含胆固醇结晶及游离胆固醇㊂粥样硬化斑块中脂质及结缔组织的含量决定斑块的稳定性及是否易导致急性缺血事件的发生㊂我们发现c i r c R N A通过调节内皮细胞㊁血管平滑肌细胞和巨噬细胞的活化,在动脉粥样硬化的发生㊁发展中起着重要作用㊂研究发现,c i r c A N R I L通过与一种重要的核糖体成分P E S1的联合作用,抑制血管平滑肌细胞和巨噬细胞中核糖体的生成,导致核糖体应激和细胞死亡,抑制平滑肌细胞和人诱导多能干细胞来源的巨噬细胞的增殖,从而起到动脉粥样硬化保护作用,是治疗动脉粥样硬化的潜在治疗靶点[4]㊂C I R C_0003204主要定位于人主动脉内皮细胞的细胞质,过表达C I R C_ 0003204抑制氧化低密度脂蛋白(o x-L D L)诱导的内皮增生[5]㊂c i r c H I P K3过表达显著降低细胞凋亡和氧化应激标志物[包括活性氧(R O S)㊁超氧化物歧化酶(S O D)和丙二醛(M D A)的水平]㊂进一步的研究表明, c i r c H I P K3通过m i R-29a/I G F-1轴抑制氧化损伤[6]㊂有研究发现,h S A-C I R C-000595在缺氧的人主动脉平滑肌细胞(H A S M C)中表达增加,h S A-C I R C-000595可能通过m i R-19a/r h o B/c y c l i n D1/C D C25A 和MM P/α-S MA/S M22α轴诱导细胞凋亡[7-8]㊂高度保守的c i r c L r p6具有m i R-145的多个结合位点,m i R-145与多个靶点相互作用,包括I T G b8㊁f a s i n㊁K L F4㊁Y e s1和l o x㊂沉默c i r c L r p6可防止小鼠颈动脉内膜增生[9]㊂有研究鉴定了缺氧条件下人脐静脉内皮细胞中差异表达的c i r c R N A,发现c Z N F292是缺氧调控下表达最高的c i r c R N A㊂有研究表明,c Z N F292的沉默显著抑制了球体萌发和管状细胞的形成,并降低了内皮细胞的增殖,表明c Z N F292在缺氧条件下促进了内皮的增殖和管状细胞的形成[10]㊂Y A N G等[11]发现,在o x-L D L诱导的血管平滑肌细胞(V S M C)中, c i r c C H F R异常过表达㊂进一步研究发现,沉默C H F R通过m i R-370/F O X O1轴抑制V S M C s的增殖和迁移能力㊂2.2c i r c R N A与冠心病冠心病是指冠状动脉粥样硬化使管腔狭窄或阻塞,导致心肌缺血㊁缺氧而引起的心脏病,是严重威胁人类健康的疾病,在西方发达国家,其年死亡数可占到总死亡数的1/3左右㊂据WHO统计,冠心病目前是世界上最常见的死亡原因,超过所有肿瘤的总和㊂该病多发生于40岁以上,男性多于女性㊂c i r c N f i x是由一个与超级增强子结合的转录因子介导的,敲除c i r c N f i x可促进心肌细胞增殖和血管生成增加,从而阻止心肌梗死后的细胞凋亡,减少心功能不全,改善心肌梗死后的预后㊂反之,c i r c F nd c3b 在心肌梗死后的小鼠心脏和缺血性心肌病患者的人类心肌组织中表达下调㊂其过表达减少了心肌细胞的凋亡,改善了血管形成和左心室功能[12]㊂c i r c N c x1在氧化应激时升高,促使心肌细胞凋亡㊂c i r c N c x1作为m i R-133a的海绵,敲除c i r c N c x1后,可通过c i r c-N c x1-m i R-133a-C D I P1(诱导蛋白)轴减少心肌细胞死亡,进一步减轻小鼠心肌细胞的缺血再灌注损伤[13]㊂c i r c T t c3通过c i r c T t c3-m i R-15b-5p-A r l2(A D P核糖化因子)调节心肌细胞的活性㊂在心肌梗死后,在小鼠体内敲除c i r c T t c3可使心脏功能显著恶化,因此c i r c T t c3在心肌梗死中的上调具有保护心脏的作用[14]㊂心肌梗死损伤和缺氧处理的小鼠心肌细胞c i r c R N A C d r1a s表达上调㊂其过表达可加重小鼠心肌梗死面积,并导致心肌细胞凋亡㊂C d r1a s充当m i R-7a的海绵,并影响其下游目标㊂此前,m i R-7a的上调在心肌梗死损伤期间被描述为保护性的㊂因此,降低C d r1a s的表达水平可能会增加m i R-7a的水平,这可能成为治疗冠心病的一种新的治疗策略[4]㊂c i r-c R N A A C R可通过调节P I N K1/F AM65B通路来抑制缺血再灌注损伤,抑制自噬性细胞死亡,从而缩小心肌梗死面积[15]㊂心肌梗死组c i r c MA C F1和E M P1 (上皮膜蛋白1)的表达水平随m i R-500b-5p表达水平的升高而降低㊂c i r c MA C F1作为m i R-500b-5p的海㊃9781㊃现代医药卫生2021年6月第37卷第11期J M o d M e d H e a l t h,J u n e2021,V o l.37,N o.11绵上调E M P1的表达,c i r c MA C F1通过调节m i R-500b-5p/E M P1轴抑制AM I的进展㊂c i r c MA C F1可能是治疗急性心肌梗死的潜在治疗靶点[16]㊂在小鼠心肌缺血再灌注损伤模型中,c i r c P A N3的表达减少㊂过度表达c i r c P A N3通过c i r c P A N3-m i R-421-P I N K1轴,显著抑制了心肌细胞的自噬并减轻了细胞凋亡,这在体内通过减少自噬空泡和缩小心肌梗死范围进一步得到证实[17]㊂2.3c i r c R N A与心肌病心肌病是一组异质性的心肌疾病,病因多与遗传有关㊂临床主要表现为心肌肥厚㊁心脏扩大㊁心力衰竭㊁心律失常与猝死㊂最早鉴定出来的心脏表达的c i r c R N A之一是抗肥厚型H R C R㊂在异丙肾上腺素诱导的小鼠心肌肥厚模型中,H R C R 水平降低,过表达的H R C R作为m i R-223-5p的海绵来减弱心肌肥厚[18]㊂分析64例肥厚型心肌病患者和53例健康对照者血清中多种c i r c R N A(包括c i r c D-N A J C6㊁c i r c T M E M56和c i r c M B O A T2)的表达模式㊂结果表明,在调整了年龄和性别后,肥厚型心肌病患者的c i r c D N A J C6㊁c i r c T M E M56和C i r-c M B O A T2基因表达明显下调,O R值分别为0.048(0.012~ 0.198)㊁0.074(0.017~0.317)和0.135(0.041~ 0.447)㊂此外,R O C曲线分析表明,这些环状R N A 可以作为H C M的生物标志物,其A U C在0.738~ 0.819之间[19]㊂c i r c A m o t l1与P D K1和A K T1结合,导致A K T1磷酸化,并可能在阿霉素诱导的心肌病中发挥心脏保护作用[20]㊂对c i r c R N A在心脏分化过程中的表达和人类心脏在胎儿组织中特异性富集的研究发现,c i r c S L C8A1㊁c i r c C A C N A1D㊁c i r c S P H K A P 和c i r c A L P K2存在差异表达[21]㊂2.4c i r c R N A与心房颤动心房颤动易形成左房附壁血栓㊂血栓栓塞,尤其是脑栓塞是重要的致残和致死的原因㊂c i r c R N A高通量测序显示房颤组H A S_ C I R C_0005643和N E V E_C I R C_0077334表达增加㊂H A S_C I R C_0005643和N O V I C E_C I R C_0077334均被预测与m i R-221-5p结合,这可能解释了m i R-221-5p在心房颤动病理生理过程中减少的原因,m i R-221-5p作为心房颤动的一个新的生物标志物值得进一步研究[22]㊂2.5c i r c R N A与心力衰竭心力衰竭是由心脏结构或功能异常所导致的一种临床综合征,是心血管疾病的最严重的阶段,死亡率高,预后不良㊂心力衰竭患者的C D R1a s在血浆中表达上调,m i R-135a和m i R-135b水平下调,Hm o x1水平明显高于对照组,且与心功能高度相关㊂进一步研究发现,C D R1a s作为m i R-135a和m i R-135b的海绵,通过m i R-135a/Hm o x1和m i R-135b/Hm o x1信号轴调控人心肌细胞的增殖和凋亡,参与C H F的发生㊁发展[23]㊂2.6瓣膜钙化过表达的c i r c S a m d4a减少了瓣膜钙化的发生,而抑制了c i r c S a m d4a则促进了瓣膜钙化,表明c i r c S a m d4a具有抗钙化的特性㊂进一步研究发现, c i r c S a m d4a是m i R-125a-3p和m i R-483-5p的m i R N A 海绵,借此来参与调节瓣膜钙化的过程[24]㊂3结语与展望c i r c R N A吸附m i c r o R N A s(M i R N A s)并抑制其内源活性㊂A N N A D O R A Y等[25]设计了人工c i r-c R N A海绵(c i r c m i R s)来靶向已知的心肌促肥厚型m i R-132和m i R-212,实验证明表达的c i r c m i R s竞争性地抑制m i R-132和m i R-212的活性,并且表现出比线性海绵更大的稳定性㊂由此我们可以设想利用人工设计的c i r c R N A靶向m i R N A来治疗心血管疾病[25]㊂心血管疾病仍然是威胁人类健康的主要疾病,早期诊断和干预能够有效改善患者的远期预后和生活质量㊂随着现代分子生物学技术的发展,有关c i r-c R N A在心血管疾病中作用机制的研究将会更加深入,c i r c R N A有望成为新的诊断标志物及治疗靶点㊂参考文献[1]Z H A O D,L I U J,WA N G M,e t a l.E p i d e m i o l o g y o f c a r d i o v a s c u l a rd i se a s e i n C h i n a:c u r r e n tf e a t u r e s a n d i m p l i c a t i o n s[J].N a t R e vC a r d i o l,2019,16(4):203-212.[2]L I N F,Y A N G Y,G U O Q,e t a l.A n a l y s i s o f t h e m o l e c u l a r m e c h a-n i s m o f a c u t e c o r o n a r y s y n d r o m e b a s e d o n c i r c R N A-m i R N A n e t-w o r k r e g u l a t i o n[J].E v i d B a s e d C o m p l e m e n t A l t e r n a t M e d,2020, 2020:1584052.[3]L I M T B,L A V E N N I A H A,F O O R S.C i r c l e s i n t h e h e a r t a n dc a rd i o v a s c u l a r s y s te m[J].C a r d i o v a s c R e s,2020,116(2):269-278.[4]G E N G H H,L I R,S U Y M,e t a l.T h e C i r c u l a r R N A C d r1a s P r o-m o t e s M y o c a r d i a l I n f a r c t i o n b y m e d i a t i n g t h e r e g u l a t i o n o f m i R-7a o ni t s t a r g e t g e n e s e x p r e s s i o n[J].P L o S O n e,2016,11(3):e0151753.[5]Z H A N G S,S O N G G,Y U A N J,e t a l.C i r c u l a r R N A c i r c_0003204i n h i b i t s p r o l i f e r a t i o n,m i g r a t i o n a n d t u b e f o r m a t i o n o f e n d o t h e l i a lc e l l i n a t h e r o s c l e r o s i s v i a m i R-370-3p/T G FβR2/p h o s p h-S MA D3a x i s[J].J B i o m e d S c i,2020,27(1):11.[6]WA N G Y,Z H A O R,L I U W,e t a l.E x o s o m a l c i r c H I P K3r e l e a s e df r o m h y p o x i a-p r e t r e a t e d c a r d i o m y o c y t e s r eg u l a t e s o x i d a t i v e d a m-a g e i n c a r d i a c m i c r o v a s c u l a r e n d o t h e l i a l c e l l s v i a t h e m i R-29a/I G F-1p a t h w a y[J].O x i d M e d C e l l L o n g e v,2019,2019:7954657.[7]S U N G,S O N G H,WU S.M i R-19a p r o m o t e s v a s c u l a r s m o o t hm u s c l e c e l l p r o l i f e r a t i o n,m i g r a t i o n a n d i n v a s i o n t h r o u g h r e g u l a-t i o n o f R a s h o m o l o g f a m i l y m e m b e r B[J].I n t J M o l M e d,2019, 44(6):1991-2002.[8]Z H E N G C,N I U H,L I M,e t a l.C y c l i c R N A h s a-c i r c-000595r e g u-l a t e s a p o p t o s i s o f a o r t i c s m o o t h m u s c l e c e l l s[J].M o l M e d R e p, 2015,12(5):6656-6662.[9]H A L L I F,C L I M E N T M,Q U I N T A V A L L E M,e t a l.C i r c_L r p6,a c i r-c u l a r R N A e n r i c h ed i n v a s c u l a r s m o o t h m u s c le c e l l s,a c t s a s a s p o n g er e g u l a t i n g m i R N A-145f u n c t i o n[J].C i r c R e s,2019,124(4):498-510.[10]B O E C K E L J N,J AÉN,H E UMÜL L E R A W,e t a l.I d e n t i f i c a t i o na n d c h a r a c t e r i z a t i o n o f h y p o x i a-r e g u l a t e d e n d o t h e l i a l c i r c u l a rR N A[J].C i r c R e s,2015,117(10):884-890.[11]Y A N G L,Y A N G F,Z H A O H,e t a l.C i r c u l a r R N A c i r c C H F R f a-c i l i t a t e s t h e p r o l i f e r a t i o n a nd m i g r a t i o n o f v a s c u l a r s m o o t h m u s c l ev i a m i R-370/F O X O1/C y c l i n D1p a t h w a y[J].M o l T h e r N u c l e i cA c i d s,2019,16:434-441.(下转第1921页)㊃0881㊃现代医药卫生2021年6月第37卷第11期J M o d M e d H e a l t h,J u n e2021,V o l.37,N o.11用,而MA L D I-T O F-M S与传统的微生物表型检测和生化检测方法相比,具有准确度高㊁灵敏度高㊁成本低㊁快速高效等优点,有良好的临床使用前景[11],因此使用MA L D I-T O F-M S质谱仪进行菌种鉴定也是有必要的㊂而未来的深入研究可结合16S r D N A测序㊁S h o t g u n等技术,以全面地研究皮肤菌群多样性与皮肤健康的联系㊂总之,表皮葡萄球菌是痤疮患者皮损局部体表和健康人皮肤表面样本中绝对优势菌种;丙酸杆菌尤其是痤疮丙酸杆菌在各类样本中检出率较高,其可能是主要的痤疮致病菌;痤疮患者皮肤菌群检出种数相较于健康人明显下降;皮肤微生态的失衡及皮肤菌群多样性的下降可能与痤疮的发生㊁发展相关㊂目前,对于皮肤微生态的研究还处在起步阶段,本研究通过对于痤疮皮肤菌群组成的初步探究,引出了皮肤微生态与痤疮患病的相关性,但此相关性还需要进一步的研究来证实㊂研究方向从针对单独一个菌种的研究向对多个菌种相互作用及人体皮肤微生态组成的研究的转变,将会为治疗痤疮提供新的思路㊂参考文献[1]R O D R I G U E S H A.T h e c u t a n e o u s e c o s y s t e m:t h e r o l e s o f t h es k i n m i c r o b i o m e i n h e a l t h a n d i t s a s s o c i a t i o n w i t h i n f l a mm a t o r y s k i n c o n d i t i o n s i n h u m a n s a n d a n i m a l s[J].V e t D e r m a t o l,2017,28(1):60-75.[2]项蕾红.中国痤疮治疗指南(2014修订版)[J].临床皮肤科杂志,2015,44(1):52-57.[3]B H A T E K,W I L L I AM S H C.E p i d e m i o l o g y o f a c n e v u l g a r i s[J].B r J D e r m a t o l,2013,168(3):474-485.[4]F I T Z-G I B B O N S,T OM I D A S,C H I U B H.e t a1.P r o p i o n i b a c t e r i-u m a c n e s s t r a i n p o p u l a t i o n s i n t h e h u m a n s k i n m i c r o b i o m e a s s o c i-a t e d w i t h a c n e[J].J I n v e s t D e r m a t o l,2013,133(9):2152-2160.[5]W I L L I AM S H C,D E L L A V A L L E R P,G A R N E R S.A c n e v u l g a r-i s[J].L a n c e t,2012,379(9813):361-372.[6]Y A N H A N W,S H E RW I N K,MU Y A S,e t a l.S t a p h y l o c o c c u s e p i-d e r m i d i s i n t h e h u m a n s k i n m i c r o b i o m e m e d i a t e s f e r m e n t a t i o n t o i n h i b i t t h e g r o w t h o f P r o p i o n i b a c t e r i u m a c n e s:i m p l i c a t i o n s o f p r o b i o t i c s i n a c n e v u l g a r i s[J].A p p l M i c r o b i o l B i o t,2014,98(1): 411-424.[7]K O N G H H,O H J,D E M I N G C,e t a l.T e m p o r a l s h i f t s i n t h e s k i nm i c r o b i o m e a s s o c i a t e d w i t h d i s e a s e f l a r e s a n d t r e a t m e n t i n c h i l-d r e n w i t h a t o p i c d e r m a t i t i s[J].G e n o m e R e s,2012,22(5):850-859.[8]T E R U A K I N,T I F F A N Y H,C H E N S,e t a l.A n t i m i c r o b i a l s f r o mh u m a n s k i n c o mm e n s a l b a c t e r i a p r o t e c t a g a i n s t s t a p h y l o c o c c u s a u r e u s a n d a r e d e f i c i e n t i n a t o p i c d e r m a t i t i s[J].S c i T r a n s l M e d, 2017,9(378):e a a h4680.[9]X I A O L I X,Z H I H E N G L,K E W E I L,e t a l.S t a p h y l o c o c c a l L T A-i n d u c e d m i R-143i n h i b i t s p r o p i o n i b a c t e r i u m a c n e s-m e d i a t e d i n-f l a mm a t o r y r e s p o n s e i n s k i n[J].J I n v e s t D e r m a t o l,2016,136(3): 621-630.[10]郑玉梅,孟若琳,彭海悦,等.青少年痤疮面部皮肤微生物群落结构变化[J].微生物学通报,2019,46(12):3414-3423. [11]胡继红,马筱玲,王辉,等.MA L D I-T O F M S在临床微生物鉴定中的标准化操作专家共识[J].中华检验医学杂志,2019,42(4): 241-249.(收稿日期:2020-08-20修回日期:2021-01-23)(上接第1880页)[12]P R E S T E S P R,MA I E R M C,WO O D S B A,e t a l.A g u i d e t o t h es h o r t,l o n g a n d c i r c u l a r R N A s i n h y p e r t e n s i o n a n d c a r d i o v a s c u l a rd i se a s e[J].I n t J M o l S c i,2020,21(10):3666.[13]L I M,D I N G W,T A R I Q M A,e t a l.A c i r c u l a r t r a n s c r i p t o f n c x1g e n e m e d i a t e s i s c h e m i c m y o c a r d i a l i n j u r y b y t a r g e t i n g m i R-133a-3p[J].T h e r a n o s t i c s,2018,8(21):5855-5869.[14]C A I L,Q I B,WU X,e t a l.C i r c u l a r R N A T t c3r e g u l a t e s c a r d i a cf u n c t i o n a f t e r m y o c a r d i a l i n f a r c t i o n b y s p o ng i n g m i R-15b[J].JM o l C e l l C a r d i o l,2019,130:10-22.[15]Z HO U L Y,Z HA I M,HU A N G Y,e t a l.T h e c i r c u l a r R N A A C Ra t t e n u a t e s m y o c a r d i a l i s c h e m i a/r e p e r f u s i o n i n j u r yb y s u p p r e s s i n ga u t o p h a g y v i a m o d u l a t i o n o f t h e P i n k1/F AM65B p a t h w a y[J].C e l lD e a t h D i f f e r,2019,26(7):1299-1315.[16]Z H A O B,L I G,P E N G J,e t a l.C i r c MA C F1a t t e n u a t e s a c u t e m y o-c a rd i a l i n f a r c t i o n t h r o u g h m i R-500b-5p-E M P1a x i s[J].J C a r d i o-v a s c T r a n s l R e s,2020[17]Z H A N G C L,L O N G T Y,B I S S,e t a l.C i r c P A N3a m e l i o r a t e sm y o c a r d i a l i s c h a e m i a/r e p e r f u s i o n i n j u r y b y t a r g e t i n g m i R-421/ P i n k1a x i s-m e d i a t e d a u t o p h a g y s u p p r e s s i o n[J].L a b I n v e s t, 2021,101(1):89-103.[18]WA N G K,L O N G B,L I U F,e t a l.A c i r c u l a r R N A p r o t e c t s t h eh e a r t f r o m p a t h o l o g i c a l h y p e r t r o p h y a n d h e a r t f a i l u r e b y t a r g e t i n gm i R-223[J].E u r H e a r t J,2016,37(33):2602-2611. [19]S O N N E N S C H E I N K,W I L C Z E K A L,D E G O N Z A L O-C A L V OD,e t a l.S e r u m c i r c u l a r R N A s a c t a s b l o o d-b a s e d b i o m a r k e r s f o rh y p e r t r o p h i c o b s t r u c t i v e c a r d i o m y o p a t h y[J].S c i R e p,2019,9(1):20350.[20]Z E N G Y,D U W W,WU Y,e t a l.A C i r c u l a r R N A b i n d s t o a n da c t i v a t e s A K T p h o s p h o r y l a t i o n a n d n u c l e a r l o c a l i z a t i o n r e d u c i n ga p o p t o s i s a n d e n h a n c i n g c a r d i a c r e p a i r[J].T h e r a n o s t i c s,2017,7(16):3842-3855.[21]L I M T B,A L I WA R G A E,L U U T,e t a l.T a r g e t i n g t h e h i g h l y a-b u n d a n tc i r c u l a r R N A c i r c S l c8a1i n c a rd i o m y o c y te s a t t e n u a t e sp r e s s u r e o v e r l o a d i n d u c e d h y p e r t r o p h y[J].C a r d i o v a s c R e s,2019, 115(14):1998-2007.[22]Z HA N G Y,S H E N H,WA N G P,e t a l.I d e n t i f i c a t i o n a n d c h a r a c-t e r i z a t i o n o f c i r c u l a r R N A s i n a t r i a l a p p e n d a g e o f p a t i e n t s w i t h a t-r i a l f i b r i l l a t i o n[J].E x p C e l l R e s,2020,389(2):111821. [23]C H E N C,S H E N H,H U A N G Q,e t a l.T h e C i r c u l a r R N A C D R1a s r e g u l a t e s t h e p r o l i f e r a t i o n a n d a p o p t o s i s o f h u m a n c a r d i o m y o c y t e s t h r o u g h t h e m i R-135a/HM O X1a n d m i R-135b/HM O X1a x e s[J].G e n e t T e s t M o l B i o m a r k e r s,2020,24(9):537-548.[24]R Y U J,A H N Y,K O O K H,e t a l.T h e r o l e s o f n o n-c o d i n g R N A si n v a s c u l a r c a l c i f i c a t i o n a n d o p p o r t u n i t i e s a s t h e r a p e u t i c t a r g e t s[J].P h a r m a c o l T h e r,2020,2020:107675.[25]L A V E N N I A H A,L U U T,L I Y P,e t a l.E n g i n e e r e d C i r c u l a rR N A s p o n g e s a c t a s m i R N A i n h i b i t o r s t o a t t e n u a t e p r e s s u r e o-v e r l o a d-i n d u c e d c a r d i a c h y p e r t r o p h y[J].M o l T h e r,2020,28(6): 1506-1517.(收稿日期:2020-11-13修回日期:2021-02-22)㊃1291㊃现代医药卫生2021年6月第37卷第11期J M o d M e d H e a l t h,J u n e2021,V o l.37,N o.11。
阿尔茨海默病的脑神经元凋亡机制_杨小慧

CHEMISTRY OF LIFE 2007,27(4)miR-289与CaMKⅡ mRNA的CaMKⅡ 3'非编码区的序列碱基配对,抑制了CaMKⅡ mRNA在运输过程中的表达。
该抑制作用一直持续到在突触的神经刺激下RISC组分在蛋白酶体作用下降解。
miRNA还参与了树突棘发育过程的调控[17]。
一个特定的miRNA,miR-134,抑制了Limk1蛋白mRNA的表达;Limk蛋白可调控树突棘的发育,而FMRP的缺失导致树突棘形态异常,两者的关系目前未明。
3. 问题与展望一直以来,X脆性综合征的研究重点是了解FMRP的功能。
在不同的发育时期、不同的细胞内位置,FMRP执行的功能是不同的。
这主要由于与FMRP结合的蛋白质分子及RNA分子的不同[6,10],其中包括非编码RNA。
这些非编码RNA引起了越来越多研究者的重视,可以预计将有更多的与X脆性综合征及FMRP相关的非编码RNA会被发现。
非编码RNA与FMRP相互作用机制的问题也有待解决,如RMRP与miRNA之间的关系,是FMRP利用miRNA作为指向靶mRNA的工具,还是FMRP作为RISC的组分参与了miRNA的翻译抑制,抑或二者都有?另有研究表明,非编码RNA可调控蛋白质功能[2],我们猜想是否存在能调控FMRP的非编码RNA呢?这些问题的解决将加深我们对X脆性综合征发病机制的理解,进而找出基因治疗的方法。
参 考 文 献[1]O'onnell WT et al. Annu Rev Neurosci,2002,25: 315-38[2]Cao X et al. Annu Rev Neurosci,2006,29: 77-103[3]Handa V et al. Nucleic Acids Res,2003,31(21): 6243-6248[4]Verdel A et al. Science,2004,303(30): 672-676[5]Jin P et al. Nat Cell Biol,2004,6(11): 1048-1053[6]Bagni C et al. Nat Rev Neurosci,2005,6(5): 376-387[7]Zalfa F et al. Cell,2003,112(3): 317-327[8]Zalfa F et al. J Biol Chem,2005,280(39): 33403-33410[9]Gabus C et al. Nucleic Acids Res,2004,32(8): 2129-2137[10]Zalfa F et al. Curr Opin Neurobiol,2006,16: 265-269[11]Wang H et al. J Neurosci,2002,22: 10232-10241[12]Bartel DP et al. Cell,2004,116(2): 281-297[13]Caudy AA et al. Genes Dev,2002,16(19): 2491-2496[14]Ishizuka A et al. Genes Dev,2002,16(19): 2497-2508[15]Jin P et al. Nat Neurosci,2004,7(2): 113-117[16]Ashraf SI et al. Cell,2006,124(1): 191-205[17]Schratt GM et al. Nature,2006,439(7074): 283-289文章编号: 1000-1336(2007)04-0307-03阿尔茨海默病的脑神经元凋亡机制杨小慧 戴雪伶 姜招峰1( 首都师范大学生命科学学院,北京 100037;1 北京联合大学应用文理学院,北京 100083 )摘要 :阿尔茨海默病(AD)是一种中枢神经系统退行性疾病,临床主要表现为认知功能障碍、行为异常及日常生活能力下降,主要神经病理改变有神经元变性、丢失引起的脑萎缩,细胞外的老年斑(senile plaque,SP)和细胞内的神经元纤维缠结(neurofibrillarytangle,NFT)。
植物MYB类转录因子研究进展

综 述Review2002 01 15收到,2002 01 28接受。
国家重点基础研究发展规划项目(973项目G1999011604)资助。
*联系人,E mai l:zywang @,Tel:021 ******** 4423。
植物MYB 类转录因子研究进展陈 俊 王宗阳*(中国科学院上海植物生理研究所,上海200032)摘要:植物M Y B 转录因子以含有保守的M YB 结构域为共同特征,广泛参与植物发育和代谢的调节。
含单一M YB 结构域的M Y B 转录因子在维持染色体结构和转录调节上发挥着重要作用,是M YB 转录因子家族中较为特殊的一类。
含两个M YB 结构域的MY B 转录因子成员众多,在植物体内主要参与次生代谢的调节和控制细胞的形态发生。
含3个M Y B 结构域的M YB 蛋白与c M YB 蛋白高度同源,可能在调节细胞周期中起作用。
关键词:M YB 结构域,M Y B 转录因子,组合调控学科分类号:Q 74随着多种模式生物基因组计划的完成,如何从这些浩如烟海的DNA 序列中揭示基因的功能以及它们有序的时空表达,已成为后基因组时代的重要课题。
人类基因组计划的完成显示人类只有30000~50000个基因,生命体是如何以如此少的基因完成如此复杂的生命活动的呢?很重要的一点在于基因的表达调控,使得每一个基因能适时、适地、适量地表达,并且使得某些基因可以产生多种功能各异的蛋白质。
真核基因的表达随细胞内外环境的改变而在不同层次上受到精确调控,如染色体DNA 水平、转录水平及转录后水平的调控等。
而转录水平的调控发生在基因表达的初期阶段,是很多基因表达调控的主要方式。
转录水平的调控指一类称为转录因子(有时又称反式作用因子)的蛋白质特异结合到靶基因调控区的顺式作用元件上,或调节基因表达的强度,或应答激素刺激和外界环境胁迫,或控制靶基因的时空特异性表达。
转录因子通常是一种模块化的蛋白,一般由几个独立的功能域组成,包括DNA 结合功能域,转录激活功能域,蛋白 蛋白相互作用功能域,信号分子结合功能域,核定位信号区等。
细胞周期蛋白D_在宫颈癌的研究进展

Advances in Clinical Medicine 临床医学进展, 2023, 13(10), 15608-15613Published Online October 2023 in Hans. https:///journal/acmhttps:///10.12677/acm.2023.13102183细胞周期蛋白D在宫颈癌的研究进展梁铭阁1,刘丽2*1黑龙江中医药大学研究生院,黑龙江哈尔滨2黑龙江中医药大学附属第一医院宫腔镜室,黑龙江哈尔滨收稿日期:2023年9月6日;录用日期:2023年10月1日;发布日期:2023年10月9日摘要宫颈癌是常见的女性生殖系统恶性肿瘤,近年来其发病率呈上升趋势,严重影响女性生活质量,给患病个人、家庭甚至社会带来了严重负担。
细胞周期蛋白D (Cyclin D)的异常表达对细胞周期的调控作用已经受到广泛认可,其在恶性肿瘤的发生、增殖、迁移、侵袭、转移中所参与的机制逐渐被研究发现。
笔者就Cyclin D的分类、Cyclin D与细胞周期调控及Cyclin D在宫颈癌发生发展中的作用进行综述,旨在为宫颈癌相关研究提供参考。
关键词细胞周期蛋白D,宫颈癌,研究进展Research Progress of Cyclin D in CervicalCancerMingge Liang1, Li Liu2*1Graduate School of Heilongjiang University of Chinese Medicine, Harbin Heilongjiang2Hysteroscopy Room,The First Affiliated Hospital of Heilongjiang University of Chinese Medicine, HarbinHeilongjiangReceived: Sep. 6th, 2023; accepted: Oct. 1st, 2023; published: Oct. 9th, 2023AbstractCervical cancer is a malignant tumor of female reproductive system. In recent years, the incidence of cervical cancer has had an upward trend, which seriously affects the quality of women’s life and brings serious burden to the affected individuals, families and even society. The abnormal expres-*通讯作者。
scoping review

scoping review
scoping review中文叫“范围审查”,是指一种较新的证据综合的方法,Scoping review调查或探索一个研究领域或主题的研究现状、程度、方法学,对于那些尚未得到广泛研究的领域,这是绘制研究领域当前研究状况的有用方法。
Scoping review能够更详细地描述特定研究领域的研究结果和研究范围,可以提供一种向决策者、从业者和消费者传播研究成果的机制。
1.确定某一特定领域中可用证据的类型。
2.确定文献中的关键概念/定义。
3.审查如何对某一特定主题或领域进行研究。
4.确定与概念相关的关键特征或因素。
5.作为系统评价的前提工作。
在系统评价开展之前可以先做Scoping review,来确定系统评价是否可行以及潜在的范围和成本。
6.确定和分析知识差距。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
REVIEWCell cycle control of pituitary development and diseaseVictor Quereda and Marcos MalumbresCell Division and Cancer Group,Centro Nacional de Investigaciones Oncolo´gicas(CNIO),Melchor Ferna´ndez Almagro3,E-28029Madrid,Spain(Correspondence should be addressed to M Malumbres;Email:malumbres@cnio.es)AbstractThe pituitary gland regulates diverse physiological functions,including growth,metabolism,reproduction,stress response,and ageing.Early genetic models in the mouse taught us that the pituitary is highly sensitive to genetic alteration of specific cell cycle regulators such as the retinoblastoma protein(pRB)or the cell cycle inhibitor p27Kip1.The molecular analysis of human pituitary neoplasias has now corroborated that cell cycle deregulation is significantly implicated in pituitary tumorigenesis.In particular,proteins involved in cyclin-dependent kinase regulation or the pRB pathway are altered in nearly all human pituitary tumors.Additional cell cycle regulators such as PTTG1/securin may have critical roles in promoting genomic instability in pituitary neoplasias.Recent experimental data suggest that these cell cycle regulators may have significant implications in the biology of putative progenitor cells and pituitary homeostasis. Understanding how cell cycle regulation controls pituitary biology may provide us with new therapeutic approaches against pituitary diseases.Journal of Molecular Endocrinology(2009)42,75–86IntroductionThe pituitary gland is a central endocrine organ that regulates basic physiological functions includinggrowth,reproduction,and metabolic homeostasis.The mammalian pituitary is composed of three lobes:the posterior pituitary(PP),the intermediate lobe(IL, atrophic in humans),and the anterior pituitary(AP).The versatile endocrine functions of the gland arecarried out by six cell types residing in the AP and IL of the pituitary gland.These cell types are defined by thehormone they produce and secrete:corticotropesproducing ACTH,thyrotropes secreting TSH,somato-tropes secreting GH,lactotropes that produce prolac-tin,gonadotropes secreting LH,and FSH,and theIL-specific melanotropes secreting MSH(Fig.1).Theadult pituitary arises from progenitors of a neuroecto-dermic primordium known as Rathke’s Pouch in a temporal and spatial-specific fashion during pituitarydevelopment(Melmed2003,Zhu et al.2007).Byembryonic day(E)9.5,specific signaling gradients induce the formation of the Rathke’s Pouch from theoral ectoderm.The major proliferation phase and thepositional determination and lineage commitment ofthe pituitary take place by mid-gestation(E11.5–E13.5) and the gland is not terminally differentiated till birth.Major pathways implicated in the development of thepituitary include the Notch and Wnt regulatory networks,which are mainly active in the early phasesof pituitary organogenesis and are essential for the emergence of somatotropes,lactotropes,and thyrotropes (Zhu et al.2007).The regulation of the proliferative ability of pituitary cells in adulthood is not well established,although different classes of stem/progeni-tor cells have been postulated(Vankelecom2007).A side population that efficiently excludes the Hoechst 33342dye has been shown to segregate with sphere-forming cells in the pituitary(Chen et al.2005). Pituitary colony-forming cells that display notable clonogenic potential have also been isolated(Leporeet al.2005).More recently,stem-cell specific markers such as SOX2C,SOX9,or OCT4in addition to other epithelial markers have been found in a single-cell layer in the marginal zone suggesting the presence of stem/progenitor cells that may contribute to cell renewal in the adult pituitary(Fauquier et al.2008, Garcia-Lavandeira et al.2008,Gleiberman et al.2008). Control of the cell cycle by cyclin-depen-dent kinases and their regulatorsThe cell cycle is the process by which cells divide into daughter cells.Cell division is traditionally divided into four phases:S phase(synthesis of DNA)in which is produced the duplication of the genome,M phase75Journal of Molecular Endocrinology(2009)42,75–86DOI:10.1677/JME-08-0146 0952–5041/09/042–075q2009Society for Endocrinology Printed in Great Britain Online version via (mitosis)in which the genetic material is segregated into two identical daughter cells,and two phases of growing and transition,called G (gap)phases (Fig.2).G1phase occurs before S phase;and G2precedes mitosis.In mammalian cells,this process is driven by several protein kinases that regulate progression through the various phases of the cell cycle.Among these kinases,cyclin-dependent kinases (CDKs)are critical regulators of the transition through the different phases of the cell cycle (Malumbres &Barbacid 2005).CDK activity isby fluctuations in the cellular concentration activators (cyclins)or inhibitors (CDK inhibitors which are regulated by specific transcriptional by mitogenic and anti-mitogenic pathways proteolysis by the ubiquitin-proteosome system.of cyclin and CDK complexes participate in the of G1/S or G2/M transitions.D-type cyclins D2,and D3)act as sensors of multiple mitogenic to activate CDK4and CDK6and to facilitate the during G1.CDK2–cyclin E (E1and E2)become active at the end of G1and in the transition from G1to S phase.E-type are substituted by A-type cyclins (A1,A2)to activate CDK2and CDK1at the end of S phase and during G2.Finally,the mitotic complex formed of CDK1–cyclin B (mostly B1and B2)is involved in the progression through G2and entry into M phases.The specific inhibitors of CDKs (CKIs)also play a major role in the cell cycle as mediators of antimi-togenic signals or checkpoint responses.They counter-act CDK function,either by blocking their activation,or by impairing substrate/ATP access.There are two families of CKIs,the INK4family and the Cip/Kip family.The INK4family (p16INK4a ,p15INK4b ,p18INK4c ,and p19INK4d )inhibits progression through G1/S by binding CDK4and CDK6.By contrast,members of the Cip/Kip family (p21Cip1,p27Kip1,and p57Kip2)have different roles depending on the CDK–cyclin complex they bind to.Association to CDK2and CDK1complexes blocks their kinase activity,whereas the role of Cip orFigure 2Control of the cell division cycle by major regulators involved in pituitary biology.S,DNA synthesis;M,mitosis;G1and G2correspond to ‘gap’phases.Quiescence is frequently referred to as G0.Figure 1Development producing cells from progenitors.Some representative transcrip-tion factors and signaling pathways are indicated.The cell cycle regulator CDK4may be involved in the post-natal production of some AP cells such as somatotropes and lactotropes.The requirement for CDK4in other pituitary cells is not clear as the whole pituitary is smaller in Cdk4-null mice.PP,posterior pituitary;IL,intermediate lobe;and AP,anterior pituitary.V QUEREDAand M MALUMBRES .Cell cycle control of the pituitary76Journal of Molecular Endocrinology (2009)42,75–86Kip binding to CDK4–cyclin D or CDK6–cyclin D complexes is unclear(Malumbres&Barbacid2005). The primary substrates of the CDKs in G1pro-gression are the members of the retinoblastoma protein family(pRB).pRB negatively regulates entry into the cell cycle and G1/S progression(Malumbres& Barbacid2001).pRB binds to the transcription factor family E2F to target cell cycle-specific genes for repression.In non-cycling cells,pRB is hypo-phos-phorylated and this active form is able to repress cell cycle progression.CDK–cyclin mediated phosphoryl-ation of pRB provokes its release from E2F factors that are then active to induce the expression of cell cycle genes required for S and M phases.Pituitary function and mouse models of cell cycle deregulationLittle is known about the implication of cell cycle regulators in pituitary gland development.However, in the last years,several mouse models of cell cycle regulators,such as pRB,CDKs,or CKIs,have suggested that some endocrine tissues such as the pituitary gland are critical targets of cell cycle deregulation in cancer and other diseases.The initial link between cell cycle regulation and the pituitary comes from the seminal genetic analysis of pRB in the mouse(Clarke et al.1992,Jacks et al. 1992,Lee et al.1992).In contrast to humans,in whom individuals who inherit one defective copy of pRB gene have a roughly90%likelihood of developing retinoblastoma at an early age(Matsunaga 1980),mice heterozygous for pRB did not develop retinoblastoma but instead developed pituitary tumors by the age of12months(Jacks et al.1992;Table1). Tumor incidence and histological phenotype of the tumors was highly dependent on the mouse strain suggesting additional modifier genes in pituitary tumor development(Leung et al.2004).Tumor incidence provoked by the partial deletion of pRB is partially reverted by a mutation in pRB effectors such as E2f1(Yamasaki et al.1998)or E2f4(Lee et al.Table1Mouse models of cell cycle-related proteins involved in pituitary biologyPituitary phenotype Incidence(%)Latency(months)ReferencesModelPituitary hyperplasiapRb C/K IL tumors10016Jacks et al.(1992)pRb C/K;E2f-1K/K IL tumors.Decreased versus pRBmutants6218Yamasaki et al.(1998)pRB C/K;E2f-4K/K IL tumors.Decreased versus pRBmutants7820Lee et al.(2002)p27K/K IL tumors10012Kiyokawa et al.(1996)andNakayama et al.(1996)p27CK K/CK K IL tumors7510.7Besson et al.(2007)p27K/K;Cdk2K/K IL tumors.No differences versusP27K/K;Cdk2C/C10012Martin et al.(2005)pRb C/K;p27K/K Cooperation in IL tumors907Park et al.(1999)p18K/K Tumors in IL and AP5015Franklin et al.(1998)p16K/K;p18K/K IL tumors.Shorter latency versus p18mutants5010Ramsey et al.(2007)p15K/K;p18K/K No differences versus p18mutants5015Latres et al.(2000)p19K/K;p18K/K No differences versus p18mutants5015Zindy et al.(2001)p21K/K;p18K/K Cooperation in IL tumors9013Franklin et al.(2000)p27K/K;p18K/K IL and AP undifferentiated tumors1003.5Franklin et al.(1998)pRB C/K;p21K/K IL tumors10012Brugarolas et al.(1998)Cdk4R24C/R24C AP tumors2515Rane et al.(2002)andSotillo et al.(2001)K5-Cdk4;p27K/K Cooperation in IL tumors1003Macias et al.(2008)Cdk4R24C/R24C;p27K/K Strong cooperation and undifferen-tiated tumors1002Sotillo et al.(2005)pRB C/K;Pttg1K/K IL tumors with decreased incidenceversus pRB mutants3013Chesnokova et al.(2005)pRB C/K;a GSU.PTTG1Overexpression of securin cooperatesin AP tumors10016Donangelo et al.(2006) Pituitary hypoplasiaCdk4K/K Defective proliferation and endocrinecell numbers100Postnatal Rane et al.(1999)Securin K/K Hypoplastic pituitary ND ND Melmed(2003)ND,Not determined.Cell cycle control of the pituitary.V QUEREDA and M MALUMBRES77 Journal of Molecular Endocrinology(2009)42,75–862002),indicating the relevance of the pRB/E2F pathway in pituitary tumorigenesis.The sole over-expression of another E2F family member,E2f3,is not sufficient to produce pituitary tumors,although these transgenic mice develop pituitary hyperplasia (Lazzerini Denchi et al.2005).The genetic analysis of pRB in the mouse clearly demonstrated a tumor suppressor function for this protein,and specifically in endocrine organs such as the pituitary.By that time,pRB function in the cell cycle was not fully explored and the relationship with the pituitary was not obvious.More than15years later,the reasons for the special sensitivity of endocrine tissues and particularly the pituitary,to pRB lost are not understood yet.However,this close relationship is not restricted to pRB protein.In1996, three groups reported multiple organ hyperplasia, including pituitary tumors in p27Kip1mutant mice (Fero et al.1996,Kiyokawa et al.1996,Nakayama et al. 1996).As in the pRB mutants,p27Kip1-deficient mice developed pituitary tumors by the age of12months (Kiyokawa et al.1996,Nakayama et al.1996). Although,in both cases the animals developed IL tumors,they present differential patterns in both the histological phenotype and the gene profile expression(Chien et al.2007).Soon after,a significant incidence of pituitary tumors was described in mice deficient in another cell cycle inhibitor,the member of the INK4family p18INK4c. Fifty percent of these animals developed aggressive pituitary tumors mostly from the IL by15months, although some tumors originated from the AP (Franklin et al.1998).Deficiency in either of the other INK4proteins,p16INK4a,p15INK4c,or p19INK4d does not result in pituitary tumors.However,genetic ablation of both p16INK4a and p18INK4c cooperates both in the incidence and the latency of the development of the pituitary tumors(median survival of10months;Ramsey et al.2007).No cooperation in pituitary tumor suppression is observed between p18INK4c and p15INK4b(Latres et al.2000)or p19INK4d(Zindy et al.2001).INK4proteins specifically inhibit CDK4and CDK6 kinases by competing with the obligate activator of these kinases,the cyclins.The relevance of INK4 proteins as key inhibitor of CDK4and CDK6is highlighted by a specific mutation in CDK4(Arg24to Cys)that prevents inhibition of this kinase by INK4 proteins.This mutation has been observed in both hereditary and spontaneous melanoma with low incidence(Malumbres&Barbacid2001).When a Cdk4R24C mutant protein is expressed in the mouse in substitution of the endogenous wild-type protein, these knock-in mice develop multiple tumors including frequent endocrine and mesenchymal tumors(Sotillo et al.2001,Rane et al.2002).Interestingly,pituitary tumors are also frequent(around25%in all the studies)in these knock-in mice suggesting the relevance of CDK4kinase activity in these neoplasias. Most of these pituitary tumors originated in the AP with an average latency of around15months.One or several cell cycle pathways in pituitary tumorigenesis?The former models suggest a clear relevance of the CDK(and their inhibitors INK4or KIP)/pRB pathway in pituitary tumorigenesis.However,the results obtained from the combination of some of these mutations in the mouse suggest a more complex molecular network.The combined deletion of pRB and p27Kip1results in shorter latency of pituitary tumors in p27(K/K);pRb(C/K)mice(Park et al.1999).In addition,the expression of p27Kip1mRNA is reduced in pituitary tumors from pRb(C/K)mice,suggesting that p27Kip1downregulation is necessary for the tumorigenicity of the pituitary even in a pRb-null background.Similarly,although p21Cip1-null mice do not develop pituitary tumors,this mutation cooperates with pRb mutation by decreasing the latency of pituitary tumors from12to9months(Brugarolas et al.1998). Similarly,both p27Kip1and p21Cip1deficiency accel-erates pituitary tumorigenesis in a p18INK4c-null back-ground(Franklin et al.1998,2000).This cooperation is dramatic in double p27Kip1;p18INK4c mutants,which develop pituitary adenomas within3months(Franklin et al.1998).Since both INK4and CIP/KIP proteins are CDK inhibitors,these results suggested that these molecules cooperate in tumor suppression by strongly inactivating CDK function in the pituitary(Fig.3).INK4proteins specifically inhibit CDK4/6kinases,whereas CIP/KIP proteins seem to preferentially inhibit CDK2and CDK1.In agreement with this model,no cooperation in pituitary tumor formation is observed in double Cdk4R24C;p18-null mice(Sotillo et al.2005).How-ever,the introduction of the mutated Cdk4R24C allele in a p27-null background dramatically accelerates the development of pituitary tumors that kill these mutant mice in8–10weeks(Sotillo et al.2005).No cooperation in pituitary tumor development is observed in mice mutant for Cdk4R24C and deficient in p21Cip1 (Quereda et al.2007).However,a dramatic cooperation in pituitary tumor development is observed in mutant mice carrying a combination of the Cdk4R24C, p21-null,and P27-null alleles(V Quereda and M Malumbres,unpublished observations).These results,together with the cooperation observed between pRb and p27Kip1(Park et al.1999),suggest the existence of two major pathways for G1/S phase deregulation in pituitary tumors.One branch is formedV QUEREDA and M MALUMBRES .Cell cycle control of the pituitary78Journal of Molecular Endocrinology(2009)42,75–of p18INK4c/CDK4/pRB,whereas the other one is represented by p27Kip1and perhaps p21Cip1(Fig.3). The preference of CIP/KIP proteins for CDK family members other than CDK4/6indicated that these inhibitors may target CDK2,the other interphase CDK involved in G1/S transition.However,p27Kip1defi-ciency provokes similar pituitary tumors in both Cdk2(C/C)and Cdk2(K/K)mice(Martin et al. 2005)indicating that CDK2is dispensible for these tumors and it is therefore not the critical target of p27Kip1.Whether the other major cell cycle protein, CDK1,is the critical target of p21Cip1or p27Kip1during pituitary tumor suppression has not been fully addressed yet.The complexity in the molecular pathways involved in pituitary tumorigenesis has recently increased after a new mouse model that suggests possible oncogenic functions of p27Kip1.In this model,the authors designed a p27Kip1mutant allele that does not bind cyclins and CDKs and is mostly localized to the cytoplasm(Besson et al.2007).These knock-in mice developed more aggressive tumors than the p27Kip1-null mice,and by6months all the animals showed aggressive pituitary tumors of the anterior lobe.This phenotype seems to be independent of the cell cycle inhibitory activity of p27Kip1and it may be related to the ability of p27Kip1to modulate stem cell function (Besson et al.2007).Finally,a completely new cell cycle pathway involved in pituitary oncogenesis is represented by PTTG1(pitu-itary tumor transforming gene)/securin,an oncogenic moleculefirst identified in GH4rat pituitary tumor cells (reviewed in Vlotides et al.(2007)and Salehi et al. (2008)).PTTG1is involved in the mitotic checkpoint that prevents abnormal chromosome segregation(see below).In addition,this protein has multiple roles in cellcycle regulation at different stages(Fig.4).The absenceof this gene provokes a decrease in the incidence of pituitary tumors in pRB heterozygous mice,probably bytriggering ARF/p53/p21-dependent senescence(Ches-nokova et al.2005,2007).Overexpression of PTTG1inthe pituitary in transgenic mice provokes pituitary hyperplasia and focal microadenomas,and cooperateswith pRB heterozygosity in higher incidence of tumorsin the AP(Donangelo et al.2006).Deregulation of the cell cycle in human pituitary diseaseThe experimental analysis of cell cycle control in mousemodels predicts that several cell cycle mutations may bepresent in human pituitary diseases.Pituitary tumors are common intracranial neoplasms that cause significantmorbidity through mass effects and/or the inappropri-ate secretion of pituitary hormones.Pituitary adenomasare common intracranial neoplasms,comprising 10–15%of diagnosed brain tumors(Landis et al.1989).Data from autopsy studies suggest that pituitaryadenomas develop in17–25%of the population(Asa& Ezzat2002,Ezzat et al.2004).Approximately,3.5–8.5% of all pituitary tumors are diagnosed prior to the age of20years(Keil and Stratakis2008).About two-thirds of pituitary tumors express and secrete pituitary hormonesand produce various endocrine syndromes.Overall,prolactinomas account for about50%of pituitary adenomas.These adenomas cause hyperprolactinemia and subsequent problems associated to a high level of prolactin in blood(hypoestrogenism or amenorrhea in women or infertility in men).GH-producing adenomas are commonly associated with acromegaly and/or gigantism.ACTH-producing adenomas are associated with Cushing’s or Nelson’s syndromes(see below).TSH-producing tumors produce thyrotoxicosis,cardiac arrythmias,tremor,and weight loss.The rare gonado-troph adenomas and the major group of non-function-ally or non-secreting adenomas result in hypogonadism, visual deficits,and headaches(Asa&Ezzat2002, Melmed2003,Ezzat&Asa2006).Several genetic and epigenetic alterations have been observed in pituitary tumorigenesis.Some classic oncogenes such as RAS or MYC are implicated in these endocrine tumors.H-RAS mutations(codon12 (Gly/Val or Arg)or18(Ala/Tre))have been reported only in pituitary carcinomas(Karga et al. 1992,Cai et al.1994,Pei et al.1994).c-MYC,on the other hand,is frequently overexpressed in all kind of pituitary tumors in a range between20and50%depending on the type of the tumor(Woloschak et al.1994,Wang et al. 1996).Among classic tumor-suppressor genes,p53Figure3Major oncogenic and tumor suppressor pathwaysregulating the cell cycle in pituitary tumors.Cell cycle control of the pituitary.V QUEREDA and M MALUMBRES79 Journal of Molecular Endocrinology(2009)42,75–86accumulation (an indication of inactive p53function)seems to be more relevant in Cushing’s adenomas and invasive non-functional tumors than in non-functioning adenomas (Buckley et al .1994,Thapar et al .1996,Clayton et al .1997).In addition to these classic cancer genes,a significant number of genetic or epigenetic alterations in pituitary tumors target several cell cycle regulators as described in the following paragraphs (Table 2).From these data,it has been estimated that more than 80%of pituitary tumors display alterations at least in one of the regulators of the G1/S transition of the cell cycle (Malumbres &Barbacid 2001).Retinoblastoma proteinAlthough,early studies did not find loss of pRB alleles (Cryns et al .1993,Zhu et al .1994),later studies found loss of heterozygosity in the human pRB gene (RB1)in malignant or highly invasive pituitary tumors (Pei et al .1995;Table 2).Several studies based on immunodetec-tion in tumor sections found abnormal expression of pRB in different pituitary adenomas.In some cases,decreased expression correlates with hypermethylation of the pRB promoter (Simpson et al .2000,Ogino et al .2005)or deletion within the protein-pocket binding domain (Simpson et al .2000).Cyclins and cyclin-dependent kinase activityCyclin D1and D3are often overexpressed in pituitary tumors (Jordan et al .2000,Turner et al .2000,Saeger et al .2001,Simpson et al .2001a )with some evidence of cyclin D1allelic imbalance in one fourth of the tumor samples analyzed (Hibberts et al .1999).In general,although cyclin D1is overexpressed in most pituitary tumor types,this overexpresion is more relevant in non-functional tumors.Cyclin E is also deregulated in human pituitary tumors,with a sign-ificant increase in corticotroph neoplasias from patients with Cushing’s disease (Jordan et al .2000).Despite the dramatic effect of Cdk4hyperactivation in mouse models (Table 1),no CDK4mutations have been identified in human pituitary tumors (Simpson et al .2001a ,Honda et al .2003,Vax et al .2003).Figure 4PTTG1/securin functions in the cell cycle (for a comprehensive review see Vlotides et al .(2007)).V QUEREDAand M MALUMBRES .Cell cycle control of the pituitary80Journal of Molecular Endocrinology (2009)42,75–86INK4inhibitorsAlthough point mutations in INK4inhibitors are not frequent in human pituitary adenomas,the expression of p16INK4a and p15INK4b is often silenced.Silencing of the p16INK4a gene(CDKN2A)by hypermethylation was first reported in the late90s(Woloschak et al.1997).A detailed analysis suggested that CDKN2A methylation was confined to particular adenoma subtypes(Simpson et al.1999)and thesefindings were subsequently confirmed by several other groups concluding that hypermethylation of the CDKN2A is the most common epigenetic deregulation in these neoplasias(Morris et al.2005,Ogino et al.2005,Yoshino et al.2007). p16INK4a is able to inhibit cell proliferation in pituitary tumor cells in correlation with a shift in the phos-phorylation status of pRB,suggesting the relevance of this CDK inhibitor in the activation of pRB and pituitary tumor suppression(Frost et al.1999).CIP/KIP inhibitorsSoon after the publication of the phenotype of p27Kip1-deficient mice,several studies interrogated the alteration of this inhibitor in human tumors.Early studies detected no p27Kip1mutations in human pituitary tumors(Tanaka et al.1997,Dahia et al. 1998).The fact that p27Kip1is haploinsufficient for tumor suppression(Fero et al.1998),however,suggests that decreased expression may be relevant in tumor development.In fact,downregulation of p27Kip1 protein expression is commonly observed in pituitary carcinomas and corticotroph adenomas,and recurrent human pituitary adenomas show lower p27Kip1protein levels than non-recurrent adenomas(Bamberger et al. 1999,Lidhar et al.1999).p27Kip1mRNA levels are not generally decreased in tumors suggesting increased proteolysis of this cell cycle inhibitor in cancer(Bloom &Pagano2003).Ubiquitin-mediated degradation of p27Kip1is controlled by SKP2,an F-box protein with diverse oncogenic functions(Frescas&Pagano2008). Whether SKP2is the relevant F-box protein for degradation of p27Kip1in pituitary tumors is not yet clear(Musat et al.2002).Degradation of p27Kip1may also be induced by JAB1(JUN activation domain-binding protein),a transcriptional cofactor for AP-1 (Chamovitz&Segal2001).In addition to this function, JAB1is able to translocate phosphorylated p27Kip1toTable2Alteration in cell-cycle regulators in human pituitary tumorsCancer-associated alteration(incidence)Tumor type References Gene(symbol)pRB(RB1)LOH(100%)Highly-invasive or malignant tumors Pei et al.(1995)Promoter hypermethylation(60%of non-expressing pRB tumors)Somatotrophinoma and non-secretingadenomasSimpson et al.(2000)Promoter hypermethylation(35%)Pituitary adenomas Yoshino et al.(2007)Promoter hypermethylation(28.6%)Pituitary adenomas Ogino et al.(2005) Cyclin D1(CCND1)Allelic imbalance(25%)Invasive and non-invasive tumors Hibberts et al.(1999) Overexpression(30–50%)Somatotrophinomas&non-functioningtumorsSimpson et al.(2001a,b) Cyclin D3(CCND3)Overexpression(68%)Pituitary adenomas(all different types)Saeger et al.(2001) Cyclin E(CCNE)Overexpression(37%)Cushing’s disease adenomas Jordan et al.(2000) Cyclin A(CCNA1)Overexpression Pituitary adenomas Nakabayashi,et al.(2001) p16INK4a(CDKN2A)Promoter hypermethylation(90%ofnon-expressing p16tumors)Different pituitary tumors Woloschak et al.(1997) Promoter hypermethylation(59%)Pituitary adenomas Yoshino et al.(2007)Promoter hypermethylation(71.4%)Pituitary adenomas Ogino et al.(2005)Reduced expression levels(62%)Non-functioning adenomas or Machiavelli,et al.(2008)Reduced expression levels(40%)Macroadenomas(all different types)Machiavelli et al.(2008) p15INK4b(CDKN2B)Promoter hypermethylation(32%)Pituitary adenomas Yoshino et al.(2007) Promoter hypermethylation(35,7%)Pituitary adenomas Ogino et al.(2005)p18INK4c(CDKN2C)Reduced expression levels ACTH-secreting adenomas Morris et al.(2005)p27Kip1(CDKN1B)Reduced expression levels(75%lessthan10%cells-expressing in thetumor)Pituitary adenomas(all different types)Bamberger et al.(1999)Reduced expression levels(100%)Corticotropes&pituitary carcinomas Lidhar et al.(1999)JAB1(COPS5)Overexpression(100%)Pituitary carcinomas Korbonits et al.(2002)p21CIP1(CDKN1A)Reduced expression levels(71%)Non-functioning adenomas Neto,et al.(2005) Overexpression(77%)Hormone-producing tumors Neto et al.(2005)Overexpression(92%)GH-producing tumors Neto et al.(2005)Securin(PTTG1)Overexpression(90%pituitarytumors)Pituitary adenomas(all different types)Zhang et al.(1999)Cell cycle control of the pituitary.V QUEREDA and M MALUMBRES81 Journal of Molecular Endocrinology(2009)42,75–86the cytoplasm for protein degradation by the protea-some.Some pituitary carcinomas display a small but significant increase in JAB1levels possibly resulting in increased p27Kip1degradation(Korbonits et al.2002). Although,genetic alterations in p21Cip1are not commonly observed,this inhibitor may also be down-regulated through epigenetic modifications in pituitary neoplasias(Yoshino et al.2007,Zhu et al.2008). Although,the majority of pituitary tumors in humans are spontaneous,in some cases they are part of genetic syndromes predisposing to pituitary and other tumors. These inherited syndromes include multiple endocrine neoplasia(MEN)-1,carney complex,familial isolated pituitary adenomas,and the Cushing’s and Nelson’s syndromes(Melmed2003,Beckers&Daly2007,Keil& Stratakis2008).The MEN-1syndrome is characterized by predisposition to pituitary adenomas,parathyroid hyperplasia,and pancreatic endocrine tumors.Pituitary adenomas affect between25and30%of MEN-1 patients(Burgess et al.1998).These patients display germ line mutations in the MEN1gene,which increase the susceptibility to all major pituitary adenoma subtypes.MEN1has been described as a direct regulator of p27Kip1and p18INK4c(Karnik et al.2005, Milne et al.2005),and loss of function of MEN1results in down-regulation of these two inhibitors with the subsequent deregulation in cell proliferation.In recent mouse models,Men1mutations cooperate with p18INK4c but not p27Kip1inactivation(Bai et al.2007) suggesting that the MEN1protein is mostly acting upstream of p27Kip1(Fig.3).Recently,a mutation in CDKN1B,the rat gene encoding p27Kip1,has been reported to be associated with a MEN-1-like syndrome in a murine model(Pellegata et al.2006).A germ line nonsense mutation in the human CDKN1B gene was also identified in a MEN1mutation-negative patient presenting with pituitary and parathyroid tumors. Expanded pedigree analysis showed that the p27Kip1 mutation was associated with the development of an MEN-1-like phenotype in multiple generations (Pellegata et al.2006).PTTG1/securinPTTG1was initially identified through a differential display analysis of gene expression in rat pituitary tumor cells(Pei&Melmed1997).PTTG1,also known as securin,is an inactivating partner of separase,the major effector for chromosome segregation during mitosis(Zou et al.1999).PTTG1is overexpressed in more than90%of all type of pituitary tumors(Zhang et al.1999).In addition,this protein is frequently overexpressed in metastatic or genomically instable tumors,suggesting a relevant role for securin in tumor progression(Perez de Castro et al.2007).Securin is regulated by CDK1-mediated phosphorylation(Holt et al.2008)suggesting a link between the control of the cell cycle by CDKs and PTTG1function(Fig.3).Despite the frequent deregulation of PTTG1in pituitary and other tumors,it is not clear yet whether its oncogenic role is mediated by its mitotic functions or the ability of PTTG1or modulate DNA repair or Sp1-mediated transcription(Vlotides et al.2007;Fig.4).Future perspectives and therapeutic implications in pituitary diseaseThe implication of cell cycle deregulation in pituitary tumorigenesis is well established from experimental data in mouse models(Table1)and the molecular pathology of human tumors(Table2).Most cell cycle mutations affect regulators of the G1/S transition in the cell cycle,including the CDK4/pRB pathway and cell cycle inhibitors such as p27Kip1(Malumbres&Barbacid 2001).The role of the pioneer pituitary tumor oncogene PTTG1is not clear at present,although it may participate in tumor development at different levels.Overall,these mutations provoke a hyperactive cell cycle that ensures unscheduled proliferation and genomic instability in pituitary tumors.On the other hand,defective cell cycle function also affects pituitary homeostasis.Cdk4deficient mice are smaller than wild-type littermates and display partial sterility(Rane et al.1999).These phenotypes are linked to hypomorphic pituitaries with a significant decrease in hormone-producing cells.In particular,Cdk4is required for post-natal proliferation of somato/lacto-trophs of the pituitary(Moons et al.2002,Jirawatnotai et al.2004).Some recent results suggest that Cdk4may also modulate cell proliferation in specific pituitary progenitor cells(Macias et al.2008).Re-expression of Cdk4in the pituitary rescues the sterility indicating that this defect is secondary to the defects in hormone-expressing cells in the pituitary(Martin et al.2003). However,that re-expression of Cdk4in the pituitary does not rescue the smaller size of Cdk4-null mice suggesting that dwarfism in these animals is not due to pituitary dysfunction(Martin et al.2003).Pttg1-deficient mice also display pituitary hypoplasia and decreased proliferation of pancreatic b-cells (Melmed2003,Vlotides et al.2007).The similarity between Cdk4and Pttg1deficiency is striking,although the molecular reasons are unclear.To what extent these cell cycle control pathways contribute to pituitary development and homeostasis is not fully understood yet.However,these experimental results may suggest a relevant relationship between cell cycle regulators and the ability of the pituitary to develop and to respond to physiological stresses.Given the relevance of cell cycle regulators in the correct function of stem cells(Janzen et al.2006,Jablonska et al.2007,Pei et al.2007,MaciasV QUEREDA and M MALUMBRES .Cell cycle control of the pituitary82Journal of Molecular Endocrinology(2009)42,75–。