肿瘤血管生成与血管生成抑制因子的研究进展
血管新生抑制剂的研究进展

V・ 2 2 Au r
l2 l 2( 0P  ̄
血 管新 生 抑制剂 的研 究进展
王春林 综逮 韩忠朝 审棱 ( 中国医学科学院 中国协和医科大学 血疽学研咒所, 3 0o 天津 1 2) 7 o
摘要:肿瘤的发生和转移依 赖于血管新 生. 这一概念的提 出为肿瘤豉血 管新生相 关性疾病 的治疗提供 了新的 方 击。基于对肿瘤血管新生机 制、 血管新生抑制荆的作用机 制豉特点的研 究 , 目前 已开发 出 了多种血管新 生抑 荆 , 其
一
态, 即该 肿瘤 处于 休 眠 期 。肿 瘤从 无 血管 新 生 期 或 血 管新生 前期 到血管新生 期 这个 过程 中存 在 一个 所 谓 的血 管新生 开关 (ni ei s .h , ag gn ,t ) 它是肿 瘤进 一 o c ,c d 步生 长 和转 移 时血管新生 形 成必 不 可少 的 。肿瘤 一
黾产 生 , 瘤 细 胞 、 周 围 的基 质 细 胞 和 内 皮 细 胞 之 肿 其
个 降调节 过 程。在 成 人 体 内 , 有某 些 生 理 过 程 只 血管 新生 , 但这个 过程仅产 生在 局 部并 且是 短暂 的 ,
如女 性月经周 期 、 毛发 的生 长 和 伤 口的 愈合 会 发 生 是 由多种 血管新生 的刺激 因子 和抑 制 因子调 控 的一 个平衡 过 程“ 而某些 病理过 程 如糖 尿病 性 视 网膜 病变 、 冠心 病 、 风湿 关 节 炎 、 瘤 及恶 性血 液 病 均 类 肿
存在 血管新 生异 常。
间发 生 相互作 用 , 它们 首 先 分 泌 和 激 活 多种 M s MP , 降解基 质 , 为肿 瘤 的生 长 和 转 移 提 供 空 问 , 然后 肿 瘤 细胞通 过分 泌 V G E F和 b G F F来 刺 激 内皮 细 胞 的 增殖 , 而形成 新的血管 , 肿瘤 的进 一步 生 长和 转 从 为 移提 供 了物质条 件 。 , 通 常 MV D值 高的肿瘤 其 预后 较 差 , 而在 某些 肿 组 织 中血 管 新生 的情 况 , 不能 特 异 地 反 映 肿瘤 而 的血 管新 生情况 。如在健 康人 的肺 组织 中也 可测 到 较高 的 M VD值 这 表 明 人类 的不 同肿瘤 其 血管 新 生 的情况 也不尽 相 同, 外 , 此 还可 通过 测定 内皮细 胞
TSP-1抗血管生成机制研究进展
各种血管生成 因素 的刺 激反 应而抑制血管生长 。本文就 T P一1 S 抗血 管生成 机制 研究 进展做一综述 。
【 关键词 】 凝血酶敏感蛋 白 一 ;t 成机 制 ; 1S管生 肿瘤 【 中图分类号 】 702 1 R 3 .3 【 文献标识码】 A 【 文章编号 】62 49 一(07 0 1 2 0 17 — 92 20 )9— 3 — 3 4
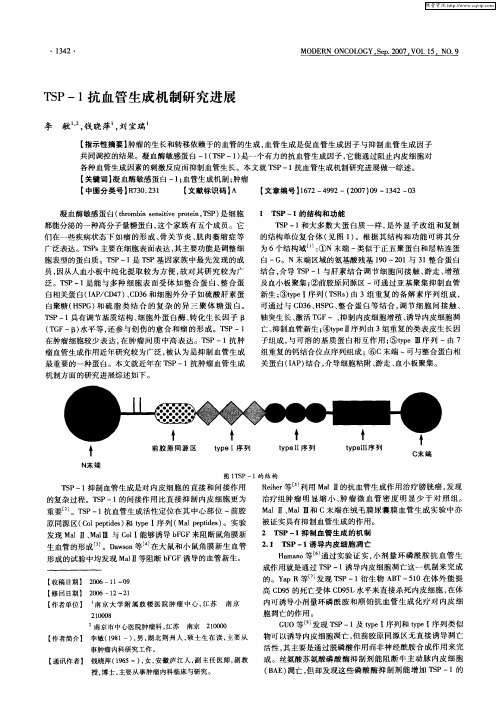
广泛表达 。T P 主要在细胞表面表达 , Ss 其主要功能是调整细
T P一1 S 和大多 数 大蛋 白质 一样 , 外显 子 改组 和复 制 是
的结构单位 复合体 ( 见图 1 。根据 其结 构和功 能可 将其 分 ) 为 6个结构域 … : ①N末端 一类似于 正五聚蛋 白和层粘 连蛋
白 一G。N末 端区域的氨基酸残基 10~2 1与 3 整合 蛋 白 9 0 1 结合 , 介导 T P一1 S 与肝 素结 合调 节细 胞 间接 触 、 游走 、 殖 增
南京 2 00 100
6 2 【 回日期】 20o —1 —21 修
江 【 作者单位 】 南 京大 学 附属 鼓 楼 医院 肿瘤 中心 , 苏 南 京
21 o 8 0 O
生血管 的形 成 ] a sn等 在大 鼠和小 鼠角膜 新生 血管 。D w o
形成的试验 中均发现 Ma Ⅱ等阻断 b G l F F诱 导的血管新生。
6 【 收稿 日期】 2oo — 11 , a ao 小剂 量环 磷酰胺 抗 血管 生
凝血酶敏感蛋 白(ho bnsniv rt n T P 是细 胞 trm i e si po i ,S ) te e
姜黄素抗肿瘤血管生成分子机制研究进展

姜黄素抗肿瘤血管生成分子机制研究进展高文;何彦津;梁凤鸣【摘要】姜黄素是从姜黄中提取的一种酚类色素,具有多种药理活性,具有抗炎、抗氧化、抗凝、降血脂、抗动脉粥样硬化、抗肿瘤、抗突变作用,其抗瘤作用机制不但与诱导肿瘤细胞凋亡相关,抗血管生成也是姜黄素抗肿瘤机制之一,主要机制为抑制血管内皮细胞增殖并促进其凋亡;抑制血管生成促进因子的表达,如通过抑制血管内皮生成因子(VEGF)及其对应受体,从而抑制肿瘤血管生成;降低基质金属蛋白酶(MMPs)的活性;通过磷脂酰肌醇-3-激酶(PI3K)信号途径抑制肿瘤血管形成:其机制为下调PI3K/AKT和促分裂原活化蛋白激酶(MAPK)途径,从而下调VEGF、Ang-1、Ang-2、VEGFR-2等.深入探讨姜黄素抗肿瘤血管生成的作用机制将为更有效的切断肿瘤血供奠定基础,有利于恶性肿瘤的防治.【期刊名称】《国际眼科杂志》【年(卷),期】2016(016)003【总页数】3页(P466-468)【关键词】姜黄素;肿瘤血管生成;血管内皮生成因子;基质金属蛋白酶;磷脂酰肌醇-3-激酶【作者】高文;何彦津;梁凤鸣【作者单位】300192,中国天津市,天津中医药大学第一附属医院眼科;300000,中国天津市,天津医科大学眼科医院;300192,中国天津市,天津中医药大学第一附属医院眼科【正文语种】中文姜黄素是从姜黄中提取的一种酚类色素,具有多种药理活性,具有抗炎、抗氧化、抗凝、降血脂、抗动脉粥样硬化、抗肿瘤、抗突变作用,其抗瘤作用机制不但与诱导肿瘤细胞凋亡相关,抗血管生成也是姜黄素抗肿瘤机制之一,主要机制为抑制血管内皮细胞增殖并促进其凋亡;抑制血管生成促进因子的表达,如通过抑制血管内皮生成因子(VEGF)及其对应受体,从而抑制肿瘤血管生成;降低基质金属蛋白酶(MMPs)的活性;通过磷脂酰肌醇-3-激酶(PI3K)信号途径抑制肿瘤血管形成:其机制为下调PI3K/AKT和促分裂原活化蛋白激酶(MAPK)途径,从而下调VEGF、Ang-1、Ang-2、VEGFR-2等。
肿瘤抗血管治疗的研究进展

选 择 性 VE R 酪 氨 酸激 酶 抑 制 剂 , GF 目前 处 于 Ⅲ期 临 床 , 有 具
良好 的 耐 受 性 和 突 出 的化 学 稳 定 性 l 。s nt i 另 一 具 有 _ 3 u i nb是 ] i 多 重 的 酪 氨 酸激 酶抑 制 活 性 的分 子 靶 向 治 疗药 物 , 以通 过 对 可医学杂志 20 09年 3月第 3 O卷第 3 期
I a m JL bMe , r 09 V 1 0N . d Mac 2 0 , o 3 , o 3 h .
・
综 述
・
肿瘤 抗血 管 治疗 的研 究进 展
赵 亮 综述 罗 以勤 审校
【 要】 实体 肿 瘤 的生 成 依 赖 于 肿 瘤本 身新 生 血 管 的 形 成 , 瘤 血 管 的 再 生 受 到 众 多 血 管 刺 激 摘 肿 及抑 制 因子 的 调控 。 随着 肿 瘤 血 管 再 生 理论 和相 应 基 础 临 床 研 究 的快 速 进 展 , 肿 瘤 血 管 生 成 治疗 抗 已成 为肿 瘤 综 合 治疗 中 的一 项 重 要 内 容 。 目前 , 有 多 种血 管 生 长 抑 制 因 子 如 血 管 抑 素 、 瘤 抑 素 等 已 肿
管 依 赖性 的 。这 为 靶 向 血管 的抗 肿 瘤 研 究 奠 定 了 坚 实 的基 础 。 实 体 瘤 的 发展 分 为 无 血 管 期 和 血管 期 , 大 多数 人 体 肿 瘤 位 于 绝 原 发 部 位 , 月 到 数年 处 于 无 血 管 状 态 , 无 血 管 阶 段 , 瘤 组 数 在 肿
厚朴对肿瘤血管生成的抑制作用机制研究

厚朴对肿瘤血管生成的抑制作用机制研究随着人们对肿瘤治疗的不断探索,发现抑制肿瘤血管生成的疗法成为了一个重要的研究方向。
肿瘤血管生成是指肿瘤细胞通过血管形成新的供血网络,为肿瘤提供营养和氧气。
研究发现,抑制肿瘤血管生成可以有效阻断肿瘤的生长和转移。
在众多的中药中,厚朴(Magnolia officinalis)作为一种植物药材,被广泛应用于中医药领域。
近年来,研究人员发现厚朴具有抗肿瘤的作用,并对其抑制肿瘤血管生成的机制进行了深入研究。
一项研究发现,厚朴中的主要有效成分为厚朴酸,具有抑制肿瘤血管生成的作用。
厚朴酸通过调节多个信号通路,包括VEGF信号通路、FGF信号通路和IL-6信号通路等,来达到抑制肿瘤血管生成的效果。
这些信号通路在肿瘤血管生成过程中发挥重要作用,因此厚朴酸的作用机制十分复杂。
厚朴酸可以通过降低VEGF的表达来抑制肿瘤血管生成。
VEGF是一种促血管生成因子,参与了血管内皮细胞的迁移和增殖过程。
研究发现,厚朴酸可以通过抑制VEGF的表达来减少血管内皮细胞的迁移和增殖,从而降低了肿瘤血管生成。
另外,厚朴酸还可以抑制FGF信号通路。
FGF即成纤维细胞生长因子,也是一种与肿瘤血管生成密切相关的细胞因子。
研究发现,厚朴酸可以抑制FGF的表达,并通过减少FGF的活性来阻断FGF信号通路。
这样一来,血管内皮细胞的迁移和血管生成过程就会受到抑制。
除了上述两条主要机制外,厚朴酸还可以抑制IL-6信号通路。
IL-6是一种细胞因子,参与了肿瘤血管生成过程中的炎症反应。
研究发现,厚朴酸可以减少IL-6的表达,并降低IL-6信号通路的活性。
这样一来,炎症反应得到减轻,进而抑制了肿瘤血管生成。
此外,厚朴还具有抗氧化和抗炎作用,可以抑制氧化应激和炎症反应对肿瘤血管生成的促进作用。
氧化应激和炎症反应是肿瘤血管生成过程中的重要因素,因此厚朴的抗氧化和抗炎作用对于抑制肿瘤血管生成非常重要。
总结起来,厚朴通过多个信号通路的调节,包括VEGF信号通路、FGF信号通路和IL-6信号通路等,来抑制肿瘤血管生成。
抗肿瘤血管生成基因治疗的研究

生成 抑 素 , 它 在大肠 菌 中单 独表 达的 产物 的 但
血管 抑制 活性 比 K e3的产 物还高 。 内皮 抑 素的 N末端 1 氨基 酸 微 序列 分析 证 实 与 胶 . 8个 原 X 、 的非胶 原 区 ( C ) . Ⅷ N 1c末端 片 段 同 源 , 说 明 内皮 抑素 是胶 原 X 、 Ⅷ的 降解产 物。胶 原 X 、
表达 、 阻断 A -i 受体配体途径 、 调血管生 成抑 素、 p'e T 上 内皮抑 素的表 达发挥 治疗作用 。对 抗肿瘤血管生成基因治疗 的若干问题 尚有争 议=抗肿癌 血管生成的基 因治疗真正走 向临
床 还 有 一段 距 离 。
关键词 :肿瘤血管 ; 血管生成刺澈因子 ; 血 管生成抑制 因子 ; 基 因治疗
子, 从而 促 进 了 血 管 生 成 。 V G E F增 加 微 血 管
移也有重 要意 义。
2 肿瘤血管生成相美园子的生物学活性
收稿 日 : 0 g - 期 2 1 92 0 )A 作者简介 : 黄若凡 0 7- , . 94 )男 上海^, 硬士研究生 ;
林庚 盒 (9 8)男 . 建 ^ . 授 。 士 生导 师 J3. , 福 教 博
摘要 :肿瘤血 管为肿瘤 生长 提供 氧气 、 营养 , 排泄代 谢产 , 对肿 瘤转 移也 有 重要意 义=
与肿瘤血 管生成有关的刺 激因子有 V G 、阱 等 , EFb 抑制 因子 有血 管生成 抑素 、 内皮抑 索、 s t 等 。基 因治疗在抗肿瘤血管生成治疗中主要 利用 病毒载体通 过下调 V G  ̄- 1 E F及其 受体
Ⅷ是 新发 现 的类 胶 原 蛋 白 , 主要 位 于血 管 的外 周 基底 膜 。它 们 本 身 并 不 参 与血 管 生 成 的调 控 。作 为完 整 的蛋 白无 抑制血 管 生成活性 。血
肿瘤血管生成相关信号研究进展

[] 王 艳 艳 , 团结 . 花多 糖 提 取 工 艺 优 化 研 究 [ ] 现 代 中 药 研 6 王 红 J.
究 与实 践 , 0 0,4 6 : 6 8 2 1 2 ( ) 5 —5
还 具 有 止 血 、 护 肝 脏 、 护 心 脑 血 管 、 氧 化 、 菌 、 疫 调 保 保 抗 抗 免
J u n l0 qh rUni riy o e iie 2 1 Vo. 3, . 4 o r a fQii a vest fM dcn , 0 2, 13 No 1
肿 瘤 血 管 生 成 相 关 信 号 研 究 进 展
张 明
【 要】 血 管生 成 是 受 各 种 具 有 促 进 或 抑 制 血 管 生 成 的 功 能蛋 白调 节 的 高度 协 调 有 序 的过 程 。促 摘 血 管形 成 因子 包括 血 管 内皮 细 胞 生 长 因子 、 纤 维 细胞 生 长 因子 、 小板 衍 生 生 长 因子 、 岛 素 样 生 长 成 血 胰
[] 咸 丽娜, 士辉 . 癌 中药研究 进展[] 中国野生 植物 资源 , 2 钱 抗 J.
2 0 2 ( ): 73 1 4 0 9, 8 4 1 — 7
胃鳞 状 上 皮 增 生 和 癌 变 有 一 定 的 阻 止 作 用 , o Mo n的 实 验 研 究 证 明 柿 叶 黄 酮 类 物 质 对 离 体 肉瘤 一 10 sro 一 10 有 8 (acma 8 ) 抗肿瘤效果 , 同时 柿 叶 对 于 防 治 食 管 增 生 性 癌 变 也 有 一 定 程 度 的 抑 制 作 用 。S imoo等 人 实 验 研 究 证 明 柿 叶 提 取 物 有 hn t
2 0 8: 7~ 9 0 6, 9 8
参 考 文 献
血管抑素作用机制的研究进展

特定 的解 剖部位而言 , 是血 管生成还 是血管抑 制 , 取决于
血管生成信号的总和与血 管抑制信号的总和之间的抗衡。 在 肿瘤原 发灶中 , 管生成切 换有利 于血管生 成 , 血 而在远处转 移灶 中 ,血 管生成切换 则有利 于血管抑 制。这种 现象 曾被 F lm n等人[ o a k 3 1 描述 , 他们在某些 肿瘤的外科治 疗 中发 现 , 当
维普资讯
一
13 一 20
重庆 医科大学学报 20 0 7年第 3 卷 第 l 期 (o ra f o g igMe i l nv ri o 7 V 12N .1 2 l J u l n qn dc i st 2 0 . o. o1 ) n o Ch aU e y 3
一
A P S n ae 的关系 , i T yt s) h Nc k等人[ 究发现 I1以浓度依赖 8 1 研 F
性 、可饱 和性 和 P H值依 赖性 的方 式结 合在 A P合 酶的 F T 1
域 中 1 亚基 羧基 未端 的一个与 亚 基相联 系的区域 , 3 并通 过 中断 1 亚基 和 亚基 的联 系来抑制 A P合 酶的功 能。 3 T 由于 A P合酶 的水解作用 只需 F 亚基 即可 , T 1 但是 A P合酶 T 的合成作用则需要 F F 10全酶 ,所 以 I 1 A P合酶的抑制 F 对 T 是单 向的, I1 即 F 抑制 F 依赖 的 A P分解酶 , 1 T 但不抑制 A P T 合成酶 。 因此 , 尽管 I 1 F 不抑制内皮细胞表面的 A P的产 生 , T 但 它可以保 护细胞表面 的 A P 使得 内皮细胞在缺氧条件下 T, 能维持 A P水平 , T 尤其是在细胞外低 P H值时。因为 A P能 T
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
肿瘤血管生成与血管生成抑制因子的研究进展摘要:肿瘤血管生成在肿瘤的生长和转移中起着重要的作用,是肿瘤生长、侵袭、转移和复发的先决条件。
肿瘤的血管生成是一个复杂的生物学过程,包括血管内皮细胞的增殖、出芽和迁移等。
这一过程中有众多肿瘤血管生成因子和抑制因子参与调节和激活血管生成的开关。
因此, 研究肿瘤血管生成相关分子在肿瘤血管生成中的作用机制, 及其对肿瘤血管生成的调控,形成一种抗血管生成疗法,来达到控制肿瘤血管生长和转移的目的, 对肿瘤的治疗具有重要意义。
关键词:肿瘤血管生成;血管生成因子;血管生成抑制因子;血管生成开关恶性肿瘤的生长和转移依赖于血管生成。
当肿瘤体积超过2-3mm3后局部缺血缺氧[ 1] , 需要产生新的毛细血管以维持肿瘤的生长, 丰富的血管可向肿瘤提供足够的营养物质, 清除各种降解产物, 肿瘤细胞亦经血管进入血液循环发生血行转移, 否则肿瘤将发生退行性坏死[ 2].新血管的形成发展取决于血管生成生长因子和血管生成抑制因子之间的动态平衡,当血管生长因子与血管抑制因子达到平衡时,血管生成不被启动; 当此平衡趋向于血管生成时,血管开关被开启,新生血管开始生成。
在正常组织中, 由于缺少血管生成生长因子或生长因子被高水平的血管生成抑制因子严格控制,血管生成的开关处于关闭状态;但在肿瘤组织中, 生长因子过度表达, 抑制因子表达过低, 改变了这个平衡, 开启了肿瘤血管生成的表型, 导致新血管的生长。
因此,有学者认为肿瘤血管形成的始动因素是血管生长因子和血管抑制因子失衡,并且这种失衡会持续出现在肿瘤生长过程中,并提出了通过抑制血管生长因子信号通路,来抑制肿瘤生长和转移。
[3]1.肿瘤血管生成开关机制实验和临床数据表明,大多数人类肿瘤生成早期不诱导血管生成和并存在于原位,在无血供数月至数年后,肿瘤中的一些细胞向血管生成的表型转变,这种现象称为血管生成开关。
这一机制的分子基础可能是血管生成抑制剂的减少以及血管生成因子产量增加所致[4]。
因此,血管生成表型的开关是由血管生成因子与血管生成抑制因子的动态变化来调节。
肿瘤新生血管形成主要有以下三个步骤:①肿瘤血管生成的启动:当肿瘤体积>2 mm3 后, 肿瘤细胞自发性增殖, 导致局部缺血缺氧, 刺激血管生成的同时产生多种促血管生成因子, 这些因子刺激产生大量蛋白酶, 从而降解基膜并形成新生血管的牙胚[5] ;②血管内皮细胞的增殖和迁移:牙胚周围的血管内皮细胞在各种促血管生长因子的作用下迅速增殖并穿过牙胚向肿瘤组织定向迁移;③肿瘤新生血管的成熟:新增殖的血管内皮细胞进一步与血管外基质和周围的间质细胞相互作用, 形成完整的血管结构[6-7]。
2.肿瘤血管生成因子肿瘤血管生成过程中, 由肿瘤细胞、内皮细胞和支持细胞等所分泌的各种生长因子是肿瘤血管生成所必需的。
它们不仅调节了内皮细胞的增殖, 还控制了血管生成的过程。
主要有血管内皮生长因子(VEGF)、血小板衍生生长因子(PDGF)、成纤维细胞生长因子(FGF)、表皮生长因子(EGF)、大肠杆菌转化生长因子(TGF)、基质金属蛋白酶(MMPS)、肿瘤坏死因子(TNF)、血管生成素、缺氧诱导因子。
2.1血管内皮细胞生长因子(VEGF)血管内皮生长因子又称血管通透因子(Vascular permeability factor,VPF),是肿瘤血管生成过程中重要的调控因子。
VEGF是特异性作用于内皮细胞的糖基化细胞有丝分裂素,它的作用有诱导血管生成[8]、刺激内皮细胞的生长和增殖[9]作为内皮细胞的存活因子[10]、增强血管的渗透性[11]、抑制内皮细胞凋亡[12]。
VEGF的结构及分类:VEGF的编码基因由8个外显子和7个内含子构成,由于mRNA不同的剪切方式,产生了VEGF121、VEGF145、VEGF165、VEGF183、VEGF189和VEGF206等6种蛋白形式。
其中VEGF165是人类组织中VEGF主要的基因产物,也是VEGF家族中最重要的血管生成调节因子[13]。
VEGF家族包括5种型:VEGF-A、B、C、D和胎盘生长因子( PIGF) ,主要由肿瘤细胞和巨噬细胞分泌[14]。
VEGF 受体( VEGFR) : VEGFR 家族成员主要是三个酪氨酸酶受体( VEGFR1-3) ,主要表达在血管内皮细胞、单核细胞和造血祖细胞。
血管内皮生长因子受体( VEGFR) 由7 个胞外Ig 样区、一个单一短链跨膜序列和一个含酪氨酸蛋白激酶( PTK) 的胞内区组成,其信号传导途径与PTK 的级联反应有关。
VEGF 与受体胞外区结合后,受体二聚体化,进而诱导受体胞内区的酪氨酸残基自动磷酸化和细胞内信号蛋白的酪氨酸磷酸化,传导胞内信号,发挥生物学效[15]。
VEGFR-1和VEGFR-2最初在内皮细胞中被发现,后来发现也部分表达在造血细胞,主要参与肿瘤血管生成。
VEGFR-3主要参与淋巴管的生成与转移。
VEGFR-2是VEGF的主要功能受体,它在VEGF 的信号转导及血管内皮生成中起主导作用。
因此,可以通过阻断VEGF/VEGFR-2的信号传导通路而抑制肿瘤新生血管的生成。
2.2血小板源性生长因子(PDGF)血小板衍生因子(PDGF)可分为分子量为31KD含有7%糖的PDGFI及28KD含4%糖的PDGFⅡ。
两者均由两条高度同源的A链及B链组成,这使PDGF具有三种形式的二聚体结构,即PDGF-AA,PDGF-BB及PDGF-AB。
体内单核/巨噬细胞是主要合成PDGF 的细胞。
在生理状态下,PDGF以α颗粒的形式储存于血小板中,受损的内皮细胞及可以分泌PDGF。
2.3 成纤维细胞生长因子(FGF)FGF 主要有酸性成纤维细胞生长因子(aFGF)和碱性成纤维细胞生长因(bFGF)两种。
它们在正常生理条件下, 参与了对细胞生长, 分化,胚胎发育和血管生长的调节。
FGF 的受体包括4种同源的酪氨酸激酶受体, 这些受体胞外均具有3个免疫球蛋白样与FGF 结合的功能区, 一个跨膜区和具有酪氨酸激酶的胞内区[16] 。
其中以bFGF 与肿瘤血管的形成关系较为密切。
有文章表明,在肿瘤组织中有高水平的bFGF 表达, 而且在一些大型肿瘤组织, 阻断VEGF 的信号传导并不能抑制肿瘤的生长, 而bFGF 则能替代VEGF 的作用;此外, VEGF 和bFGF 在体外血管形成过程中, 也有协同作用:VEGF 体外血管生成作用以及诱导纤溶酶原激活物(PA)的能力依赖于由内皮细胞产生的bFGF[17] 。
2.4表皮生长因子(EGF)它由11名成员和4个表皮生长因子受体(EGFR)组成。
EGFR通路上调促血管生成因子,如VEGF等,因此被视为血管生成的间接调节因子[13]。
2.5转化生长因子(TGF)TGF-β是由两个结构相同或相近的、分子量的12.5kDa亚单位借二硫键连接的双体。
几乎每种细胞类型均可产生TGF-β,其可参与血管形成、胚胎发育和伤口愈合同时具有较强的生长抑制性能。
不同水平的TGF-β可分别对血管生成产生促进或者抑制作用。
低水平的TGF-β通过上调血管生成因子促进血管的生成,而高水平的TGF-β则通过阻止pRb磷酸化抑制内皮细胞的生长和增殖 [18].2.6基质金属蛋白酶(MMPS)基质金属蛋白酶是一个大家族,因其需要Ca、Zn等金属离子作为辅助因子而得名。
其家族成员具有相似的结构,一般由5个功能不同的结构域组成 :(1)疏水信号肽序列;(2)前肽区,主要作用是保持酶原的稳定。
当该区域被外源性酶切断后,MMPs酶原被激活;(3)催化活性区,有锌离子结合位点,对酶催化作用的发挥至关重要;(4)富含脯氨酸的铰链区;(5)羧基末端区,与酶的底物特异性有关。
其中酶催化活性区和前肽区具有高度保守性。
MMPs通过降解细胞外基质和释放血管生成丝裂原存储在基质中,诱导肿瘤血管生成。
MMP-9、MMP-2通过蛋白水解的方式裂解并活化潜在的TGF-β,促进肿瘤血管生。
[18]2.7肿瘤坏死因子(TNF)TNF是由巨噬细胞、肥大细胞和T淋巴细胞释放的细胞因子。
作为一种巨噬细胞活化因子,其并激活这些细胞分泌血管生成因子,促进血管生成。
2.8血管生成素血管生成素1和2可通过其对Tie受体的激动和拮抗作用分别作为促血管生成和抗血管生成因子。
血管紧张素1刺激Tie 2,而血管紧张素2不激活受体并作为血管紧张素1的竞争性抑制剂。
2.9缺氧诱导因子(HIF)缺氧是肿瘤和非肿瘤性疾病血管生成的重要刺激因素,缺氧诱导因子包含HIF-1、HIF-2两种,其中HIF-1在血管生成的作用最为重要。
3.肿瘤血管生成抑制因子,肿瘤血管生长抑制因子通过直接抑制肿瘤血管生成,或通过抑制肿瘤生长刺激因子的生成间接抑制肿瘤血管的生成,而实现对肿瘤的控制。
现已识别的血管生长抑制因子有: 干扰素(INF)、白细胞介素(IL)、基质金属蛋白酶抑制剂(TIMP)、血管抑素、内皮抑素。
3.1干扰素(INF)干扰素分为IFN-α、IFN-β、IFN-γ三个亚型。
干扰素具有抗病毒、调节免疫及抗肿瘤作用。
干扰素与细胞表面的特异性受体结合,除了能诱导经典的JAK-STAT 途径,还能诱导IRF 途径、P38MAPK 途径、IRS-PI3K 途径、CrKL 途径而发挥其生物学效应。
肿瘤新生血管可为不断浸润生长的原发肿瘤提供营养,它们在肿瘤的发展转移过程中起到了重要的促进作用。
干扰素具有抗肿瘤血管生成、抑制肿瘤转移的作用[19]。
目前,IFN -α在治疗肿瘤的直接浸润和转移方面的影响还是未知的,但是,IFN -α可通过IRF信号转导途径而呈现出其抗细胞增殖效应。
[20]3.2白细胞介素(IL)白细胞介素即是由多种细胞产生并作用于多种细胞的一类细胞因子。
白细胞介素(IL)的结构决定其功能的发挥,其具有促进或抑制血管生成的作用。
IL 1a 是由活化的巨噬细胞分泌的一种细胞因子,通过促血管生成因子表达的增加,诱导血管生成。
在人类肿瘤中,IL 12可抑制VEGF mRNA的表达,促进细胞凋亡和抑制增殖率,减少肿瘤血管密度[ 21,22 ]。
IL-10下调肿瘤相关巨噬细胞分泌的VEGF、IL-1β、TNFα、IL-6和 MMP 9 [ 23 ]。
3.3基质金属蛋白酶抑制剂(TIMP)内皮细胞的迁移和增殖过程需要基底膜的降解和细胞外基质的重塑。
在细胞外基质重塑的过程中,基质金属蛋白酶(MMPs)具有中枢作用,而TIMP则通过抑制周围基质的降解抑制新生血管生成。
TIMP-1、TIMP-2可以抑制βFGF诱导的内皮细胞增殖,从而抑制内皮细胞通过明胶迁移 [24]。
3.4血管抑素( AS)血管抑素是血浆纤维蛋白溶解酶原和( 或) 纤维蛋白溶解酶的降解产物,它在生理状态下并不产生,而是在原发肿瘤存在的状态下,由肿瘤细胞或肿瘤组织浸润细胞产生或活化某些蛋白酶水解纤溶酶原和( 或) 纤溶酶形成。