真核生物转录特点与加工
原核生物和真核生物基因表达调控复制、转录、翻译特点的比较

原核生物和真核生物基因表达调控、复制、转录、翻译特点的比较1.相同点:转录起始是基因表达调控的关键环节①结构基因均有调控序列;②表达过程都具有复杂性,表现为多环节;③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性;2.不同点:①原核基因的表达调控主要包括转录和翻译水平。
真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。
②原核基因表达调控主要为负调控,真核主要为正调控。
③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。
④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。
⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。
原核生物基因以操纵子的形式存在。
转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。
翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。
真核生物基因表达的调控环节较多:在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。
在转录水平主要通过反式作用因子调控转录因子与TATA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。
在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。
在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。
真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。
RNA的转录及加工
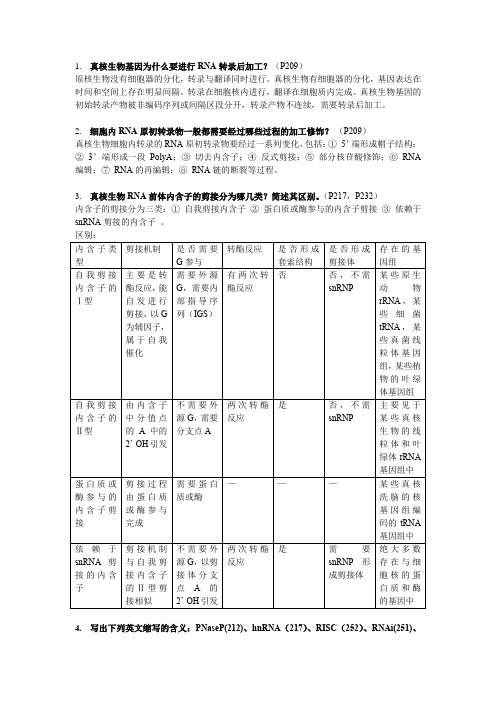
1.真核生物基因为什么要进行RNA转录后加工?(P209)原核生物没有细胞器的分化,转录与翻译同时进行。
真核生物有细胞器的分化,基因表达在时间和空间上存在明显间隔。
转录在细胞核内进行,翻译在细胞质内完成。
真核生物基因的初始转录产物被非编码序列或间隔区段分开,转录产物不连续,需要转录后加工。
2.细胞内RNA原初转录物一般都需要经过哪些过程的加工修饰?(P209)真核生物细胞内转录的RNA原初转录物要经过一系列变化,包括:①5’端形成帽子结构;②3’端形成一段PolyA;③切去内含子;④反式剪接;⑤部分核苷酸修饰;⑥RNA 编辑;⑦RNA的再编辑;⑧RNA链的断裂等过程。
3.真核生物RNA前体内含子的剪接分为哪几类?简述其区别。
(P217,P232)内含子的剪接分为三类:①自我剪接内含子②蛋白质或酶参与的内含子剪接③依赖于snRNA剪接的内含子。
区别:4.写出下列英文缩写的含义:PNaseP(212)、hnRNA(217)、RISC(252)、RNAi(251)、剪接体(220)、自我剪接(228)、反义RNA(251或上课PPT)、RNA干涉(251)、siRNA(252)、选择性剪接(235)、核酶(229)PNaseP:催化切除5’端额外核苷酸的酶hnRNA:核内不均一RNARISC:沉默复合物RNAi:RNA干涉剪接体:是mRNA前提在剪接过程中组装形成的多组分复合物,由多种snRNA和蛋白质因子组成,即剪接体是具有催化剪接过程的核塘核蛋白复合体。
自我剪接:rRNA的内含子能够自我剪接,无需剪接体反义RNA:与mRNA互补的RNA分子,也包括与其它RNA互补的RNA分子RNA干涉:在双链RNA引导的抑制过程中存在某种扩增效应,且有某种没活性参与其中。
siRNA:短干涉RNA,发生转录后基因沉默的小的双链RNA选择性剪接:一个基因的初始转录产物在不同的分化细胞、不同的发育阶段乃至不同的生理状态下,可以有不同的剪接方式,得到不同的成熟mRNA和蛋白质产物核酶:RNA本身具有酶的活性称为核酶5.名词解释:套索结构(219)、转酯反应(227)、Dicer酶(253)、顺式剪接(239)、反式剪接(239)套索结构:RNA剪接过程中的中间结构,其中有形成的带尾巴的环形结构转酯反应:在剪接体上完成剪接反应的生化本质是磷酸二酯键的转移,又称转酯反应Dicer酶:能将双链RNA特异性切成大小均一的片段的酶称为Dicer酶顺式剪接:存在与同一基因中的两个或多个外显子和内含子的剪接,称为顺式剪接反式剪接:几个外显子不在同一基因甚至不在同意染色体上的剪接叫反式剪接6.什么是RNA的自我剪接?自我剪接有哪些类型?(217或232)RNA的自我剪接:能自发进行剪接,无需酶或蛋白质参与。
真核生物tRNA的转录后后加工

剪接过程需两次转酯反应
二次转酯反应
辽宁医学院
三、真核生物前体tRNA的加工包括核
苷酸的碱基修饰
DNA
TGGCNNAGTGC
GGTTCGARNA核苷酸转移酶、 连接酶
ATP ADP
(2) (1)
碱基修饰
(1)甲基化
(1)
因此,mRNA的降解必须首先解除这些稳定因素,脱 腺苷酸化及帽结构的水解是其中的重要步骤,故称为依赖 于脱腺苷酸化的mRNA降解。
依赖于脱腺苷酸化 的mRNA降解
(二)无义介导的mRNA降解是重要的真核 细胞mRNA质量监控机制
真 核 细 胞 mRNA 的 异 常 剪 接 可 能 会 产 生 无 义 (nonsense)的终止密码子,由此产生的mRNA降解称为 无义介导的mRNA降解(nonsense-mediated mRNA decay, NMD),是广泛存在的mRNA质量监控的重要机制。
一些噬菌体的mRNA前体及细菌tRNA前体也发现有这 类 自 身 剪 接 的 内 含 子 , 并 被 称 之 为 I 型 内 含 子 ( group I intron)。I型内含子以游离的鸟嘌呤核苷或鸟嘌呤核苷酸作 为辅因子完成剪接。鸟嘌呤核苷或鸟嘌呤核苷酸的3′-OH与 内含子的5′-磷酸共同参与转酯反应。这种转酯反应与前述的 mRNA内含子剪接的转酯反应类似,不过参与反应的不是分 支点A的2-OH,切除的内含子是线状,而不是“套索”状 。
那些含有提前终止密码子(premature translationaltermination codon, PTC)的mRNA会被选择性清除。
脱腺苷酸化的mRNA 降解; 后者包括无义介导的mRNA 降解、无终止降解、无停
真核生物基因的转录

(B’’, TBP, BRF)
TF III B
TF III A TF III C
Pol III
四、RNA 聚合酶 II 基因的转录
(一)RNA聚合酶 II 的启动子 1、组成:
核心启动子(core promoter): TATA盒(Hogness box): - 25 ~ -35bp
上游启动子(upstream promoter element,UPE) CAAT盒 :-70 ~ -80区 GC盒:-80 ~ -110区
TF II B —— 覆盖靠近起始点的启动位置,C端与TFIID和DNA 的复合物结合,N-端与TFⅡF协同作用募集RNA聚 合酶II。
TF II F ——结合Pol II并带向启动子;RAP74(ATP依赖性解 旋酶),RAP30(与细菌因子有同源性)
TF II E —— 扩大DNA覆盖区至+30
Module Consensus DNA bound Factor Distribution
TATA box TATAAAA
~10bp
CAAT box # GGCCAATC ~22bp
GC box
GGGCGG
~20bp
Octamer # ATTTGCAT
~20bp
``
``
23bp
B
GGGACTTTCC ~10bp
(2) TFIIA
▪ 含有至少3个亚基 ▪ 与TFIID结合,稳定TFIID-DNA复合体;可能通过解除
TAFs的抑制而激活TBP
TF II A
(3) TFIIB ▪ 覆盖靠近起始点的启动位置,C端与TFIID和DNA的复
合物结合,N-端与TFⅡF协同作用募集RNA聚合酶II
细菌和真核生物转录机制的差异

细菌和真核生物转录机制的差异
1不同的转录机制
细菌和真核生物的转录机制存在一定的差异。
这些差异主要体现在以下几个方面:
1.1启动子
首先,启动子是转录反应开始的地方,其作用是连接RNA聚合酶与DNA模板上。
细菌只有一种RNA聚合酶,它能识别相对简单的DNA 启动子。
而真核生物有三类不同的RNA聚合酶,它们能够识别不同的DNA启动子。
1.2转录调节
其次,细菌的转录调节主要依赖启动子及其上游的启动子元件,而真核生物的调节除了启动子以外,还受到基因间まゝ数距离,和内部及外部环境因素的影响。
1.3转录加工
最后,细菌转录后不会经过加工就会被进行翻译,而真核生物经过mRNA加工后再被翻译,包括5'修饰(5'cap)、流式处理(polyA tail)、剪切(splicing)、转录本结构调节(RNA stability)等。
总之,细菌和真核生物的转录机制的主要差异体现在启动子的多样性、转录调节的复杂性及转录加工的必要性上。
《现代分子生物学》第五章 3 真核生物的转录后加工

各种参与剪接的成分形成一个剪接体系, 称为剪接体(spliceosome)。该体系由 几种snRNP和大量的其他的蛋白质分子 组成,这些蛋白质分子称为剪接因子, 估计有40多种。 剪接点和分支点序列由剪接体识别, snRNA和蛋白质都参与了识别,特别是 snRNA之间以及与mRNA间的碱基配对 起重要作用。
RNA印迹法(RNA blotting)可用于分析核 RNA剪接过程中的中间体,从而确定内含 子的去除顺序。 实验中可以发现含有不同内含子的中间产 物,因此内含子的去除似乎没有特定的顺 序,但也发现有一定的规律,说明剪接有 特定的途径。
剪接反应并不是按内含子在RNA前体上 的顺序进行的。 RNA的构象影响剪接点的选择。在去除 特定的内含子后,RNA的构象发生一定 的变化,再选择新的剪接点。
剪接体Splicesome: snRNA:U1, U2, U4, U5, U6 snRNP:U+蛋白 质
剪接体按一定的顺序组装,已分离到一些 组装的中间体。只有组装完整的剪接体才 有功能。在剪接反应中,剪接体还会释放 和添加某些成分。 转酯反应只是磷酸酯键的直接转移,没有 水解反应的出现,因此不需要外部的能量, 也不需要供能物质如ATP或GTP的参与。 剪接体中起催化转酯反应的成分尚未弄清 楚,也不知道是蛋白质还是RNA在起作用。
一、核基因mRNA的剪接 一、核基因mRNA的剪接
mRNA的基因在低等真核生物中只有少数含 有内含子,为不连续基因。随着进化程度的 增高,不连续基因的数目不断增加。 细胞核中有一类RNA,十分不稳定,平均长 度比mRNA要长,序列的复杂程度非常高, 称为核内不均一RNA( heterogeneous nuclear RNA,hnRNA )。
RNA processing 1
真核生物转录特点

真核生物RNA的转录与原核生物RNA的转录过程在总体上基本相同,但是,其过程要复杂得多,主要有以下几点不同(图3-27)。
⒈真核生物RNA的转录是在细胞核内进行的,而蛋白质的合成则是在细胞质内进行的。
所以,RNA转录后首先必须从核内运输到细胞质内,才能指导蛋白质的合成。
⒉真核生物一个mRNA分子一般只含有一个基因,原核生物的一个mRNA分子通常含有多个基因,而除少数较低等真核生物外,一个mRNA分子一般只含有一个基因,编码一条多态链。
⒊真核生物RNA聚合酶较多在原核生物中只有一种RNA聚合酶,催化所有RNA的合成,而在真核生物中则有RNA聚合酶Ⅰ、RNA聚合酶Ⅱ和RNA聚合酶Ⅲ三种不同酶,分别催化不同种类型RNA的合成。
三种RNA聚合酶都是由10个以上亚基组成的复合酶。
RNA聚合酶Ⅰ存在于细胞核内,催化合成除5SrRNA 以外的所有rRNA的合成;RNA聚合酶Ⅱ催化合成mRNA前体,即不均一核RNA(hnRNA)的合成;RNA 聚合酶Ⅲ催化tRNA和小核RNA的合成。
⒋真核生物RNA聚合酶不能独立转录RNA 。
原核生物中RNA聚合酶可以直接起始转录合成RNA ,真核生物则不能。
在真核生物中,三种RNA聚合酶都必须在蛋白质转录因子的协助下才能进行RNA的转录。
另外,RNA聚合酶对转录启动子的识别,也比原核生物更加复杂,如对RNA聚合酶Ⅱ来说,至少有三个DNA的保守序列与其转录的起始有关,第一个称为TATA框(TATA box),具有共有序列TATAAAA,其位置在转录起始点的上游约为25个核苷酸处,它的作用可能与原核生物中的-10共有序列相似,与转录起始位置的确定有关。
第二个共有序列称为CCAAT框(CCAAT box),具有共有序列GGAACCTCT,位于转录起始位置上游约为50-500个核苷酸处。
如果该序列缺失会极大地降低生物的活体转录水平。
第三个区域一般称为增强子(enhancer),其位置可以在转录起始位置的上游,也可以在下游或者在基因之内。
原核生物和真核生物基因表达调控复制、转录、翻译特点的比较

原核生物和真核生物基因表达调控、复制、转录、翻译特点的比较1.相同点:转录起始是基因表达调控的关键环节①结构基因均有调控序列;②表达过程都具有复杂性,表现为多环节;③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性;2.不同点:①原核基因的表达调控主要包括转录和翻译水平。
真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。
②原核基因表达调控主要为负调控,真核主要为正调控。
③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。
④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。
⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。
原核生物基因以操纵子的形式存在。
转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。
翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。
真核生物基因表达的调控环节较多:在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。
在转录水平主要通过反式作用因子调控转录因子与TA TA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。
在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。
在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。
真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 转录因子与反式作用因子: 调节基因表达的蛋白为转录因子;
它们与特异的顺式作用元件相互作用, 并激活另一基因的转录
转录起始复合物
F
AD
TATA
结合顺序
B
RNApolⅡ
E
DNA
TF II D TF II A TF II B RNA-pol II/TF II F TF II E
一、mRNA前体的加工
(一)生成特点: 原核 :mRNA是多顺反子
(polycistronic),每分子RNA中含几种蛋白 质信息,RNA寿命短,转录没完时翻译即开 始
例:乳糖操纵子,含Z,Y,A三个基因,分别 编码半乳糖苷酶、透过酶及乙酰基转移酶
• 真核:单顺反子,一个mRNA仅编码一种蛋白质 外显子: 在真核生物基因中编码蛋白质的
(一)特点: 1、rRNA拷贝多,原核5-10个拷贝;真核:
果蝇260个拷贝;Hela细胞1100个拷贝
2、rRNA基因之间以纵向串联的方式重复 排列
(二)加工过程
1、剪切作用:需核酸酶参与 2、甲基化修饰:修饰在碱基上 3、自我剪接:一种核酶的作用
5S的加工变化不大 原核rRNA加工:rRNA含非转录的间隔区,其产 物中含tRNA 真核rRNA加工: 1. 5S自成体系加工少无修饰和剪接。 2. 45S加工中含剪切和甲基化修饰,需核酸酶
序列
内含子:非编码蛋白的序列,因其插于外显 子之间又称插入序列,居间序列
hnRNA: 为mRNA的前体,转录物中外显子与 内含子间隔排列,需经剪接加工后生成mRNA
(二) mRNA前体加工过程
• 真核:
5‘末端帽子
部位:核内
3‘端多聚A尾 部位:核内,胞质有酶也可进行
剪接作用
部位:胞核
甲基化(甲基化发生在剪接之前,在非编码区分子 中含1-2个m6A )
3’加尾
转录的终止
加尾信号
GC丰富序列
AATAAA ----- GTGTGTG
AAUAAA GUGUGUG
酶切
加尾
AAUAAA
GUGUGUG
AAUAAA
AAAAAAAAA……….
polyA尾(约20~200个A)
绝大部分真核生物 mRNA 3`端含 poly(A) 尾巴
高等真核生物(不包括酵母)的mRNA的共同 特征:poly(A)添加位点上游11-30个核苷酸区域 内存在高度保守的AAUAAA序列。
RNA编辑(某些mRNA的核苷酸序列,在生成转录产 物后还需插入、删除或取代一些核苷酸残基,方能生 成具有正确翻译功能的模板,遗传信息在mRNA水平上 的改变过程,称为RNA编辑)
原核:多顺反子mRNA在RNaseⅢ作用下转变为单独的顺反子
5’加帽
磷酸酶
pppG
5’pppG…
5’pG
ppi …
pi
断裂基因:真核生物的基因由若干个编
码区和非编码区互相隔开但又连续镶嵌而成
外显子(exon): 基因中出现在mRNA的序列
内含子(intron): 基因中不出现于mRNA而被剪接掉的序列
核内不均一RNA (hnRNA):真核生物中编码蛋白 质基因的初级转录本,包括5’帽子、3’poly(A)尾、 外显子、内含子,经剪接加工成为mRNA
poly(A)尾巴生物学功能:
I: 保持mRNA的稳定性,防止被降解; II: 与翻译起始有关。
应用一: ★Poly(A用来
从总RNA中分 离mRNA
应用二:
可设计寡聚Oligo (dT)片段合成cDNA mRNA: 5` NNNN…..NNNNNNAAA…AA 3`
TTT…TT 5`
cDNA: 3` NNNN…..NNNNNNTTT…TT 5`
剪接
• 剪接:在细胞核内, hnRNA剪切掉内含子,将多个外 显子连接为成熟mRNA的过程为剪接 例:同一转录本, 在不同的组织, 因剪接差异 产生各自不同的mRNA
• 剪接的本质:磷酸酯键的转移 • 剪接特点 :剪接部位的结构为内含子末端的特定
序列,分布在内含子的三个部位,5‘端剪切点为 GU;3’端剪切点为AG;靠近3‘端含A序列的分支点
甲基化酶
5’GpppG
mGpppG…
…
+CH3
O
N
H2N
N
帽0
C H3
7N+
9 N
帽1 帽2
N1
N2
N3
2'
O C H3
OCH
3
OH
OOO
O CH 2 O P O P O P O
P
P
P
HH
OH OH OH
H
H
OH OH
甲基鸟苷5’,5’-三磷酸
mRNA 5`末端帽子特征的生物学功能:
I: 使mRNA免遭核酸酶的破坏,保持其结构的稳定性; II: 利于蛋白质起始因子的识别,从而促进翻译的起始。
TATA
PIC
12 3
4
5
转录后的加工
原核RNA不需加工,边转录边翻译
rRNA 真核RNA需要
加工
tRNA
mRNA
5’加帽 在转录过 3’加尾 程中
剪接 转录后进行
真核生物转录后的加工
• 意义:转录生成的前体RNA,无生物学活性, 需加工变为成熟、有活性的RNA
• 加工种类:剪切与剪接、末端添加核苷酸、 修饰、RNA编辑
内含子
外显子 套索结构
二、tRNA前体的加工
1、切除多余的核苷酸:RNase P切除5'端多 余的核苷酸;RNase D切除3'端多余的核苷酸 2、剪切内含子:核酸内切酶切除内含子,连 接酶进行连接 3、修饰与3‘末端加-CCA: 修饰包括甲基化,脱 氨基,还原反应等,在核苷酸基转移酶催化下 完成3’末端添加CCA
tRNA的加工
内含子编码区
5
PPP
Байду номын сангаас
DNA
3
外切核酸酶
-OH 3’
内切核酸酶
3端加CCA-OH
CCA-OH 3
P
初级转录物
•5’和3’端的酶切 •3’加上CCA •去掉内含子 •特异部位碱基的加工
内含子 5 CCA-OH 3
内含子 除去内含子 (内切酶与连接酶)
成熟 tRNA
三、rRNA前体的加工
• mRNA的剪接:
hnRNA被剪接体作用,剪除内含子、 连接外显子,产生成熟的mRNA的过程
剪接体
mRNA的剪接
DNA mRNA
转录
外显子 内含子
形成套索RNA,外显子靠近
去除套索RNA,外显子连接 成熟mRNA
内含子5’端断开 外显子
前一个外显子的3’末端攻 击下后个外显子的5末端’
前后外显子的连接