真核生物的转录过程共20页
真核生物基因的转录

(B’’, TBP, BRF)
TF III B
TF III A TF III C
Pol III
四、RNA 聚合酶 II 基因的转录
(一)RNA聚合酶 II 的启动子 1、组成:
核心启动子(core promoter): TATA盒(Hogness box): - 25 ~ -35bp
上游启动子(upstream promoter element,UPE) CAAT盒 :-70 ~ -80区 GC盒:-80 ~ -110区
TF II B —— 覆盖靠近起始点的启动位置,C端与TFIID和DNA 的复合物结合,N-端与TFⅡF协同作用募集RNA聚 合酶II。
TF II F ——结合Pol II并带向启动子;RAP74(ATP依赖性解 旋酶),RAP30(与细菌因子有同源性)
TF II E —— 扩大DNA覆盖区至+30
Module Consensus DNA bound Factor Distribution
TATA box TATAAAA
~10bp
CAAT box # GGCCAATC ~22bp
GC box
GGGCGG
~20bp
Octamer # ATTTGCAT
~20bp
``
``
23bp
B
GGGACTTTCC ~10bp
(2) TFIIA
▪ 含有至少3个亚基 ▪ 与TFIID结合,稳定TFIID-DNA复合体;可能通过解除
TAFs的抑制而激活TBP
TF II A
(3) TFIIB ▪ 覆盖靠近起始点的启动位置,C端与TFIID和DNA的复
合物结合,N-端与TFⅡF协同作用募集RNA聚合酶II
真核生物转录起始复合物的形成过程

真核生物转录起始复合物的形成过程转录是生物体内基因表达的关键过程之一,它是将DNA的信息转录成RNA的过程。
在真核生物中,转录的起始步骤是形成转录起始复合物,它由转录因子和RNA聚合酶组成。
本文将详细介绍真核生物转录起始复合物的形成过程。
转录因子是转录起始复合物中的重要成分,它们能够识别和结合到基因的启动子区域。
转录因子一般分为两类:一类是通用转录因子,另一类是特异性转录因子。
通用转录因子存在于所有真核生物中,它们的主要作用是在转录起始复合物形成过程中提供稳定的结构支持。
特异性转录因子则根据不同的基因和细胞类型的需要,选择性地结合到启动子区域,调控基因的转录。
在转录因子的辅助下,RNA聚合酶开始结合到启动子区域。
RNA聚合酶是一个复合酶,它负责合成RNA分子。
在真核生物中,有三种不同类型的RNA聚合酶,分别为RNA聚合酶Ⅰ、Ⅱ和Ⅲ。
其中,RNA聚合酶Ⅱ是最为重要的,它负责合成mRNA分子。
当RNA聚合酶Ⅱ与启动子结合后,转录因子会形成一个稳定的结构,这就是转录起始复合物的形成。
转录起始复合物形成后,RNA聚合酶Ⅱ开始进行转录作用。
它会解开DNA的双链结构,使得其中的一个链作为模板合成RNA分子。
在合成过程中,RNA聚合酶Ⅱ会逐渐沿着DNA链上游移动,同时合成RNA链。
这个过程称为转录延伸。
转录延伸的速度和效率受到多种因素的调控,包括转录因子的结合和启动子的组装状态等。
一旦RNA聚合酶Ⅱ合成到基因的终止信号区域,转录作用就会停止。
在这个时候,RNA聚合酶Ⅱ会与已合成的RNA分子和DNA模板解离。
随后,RNA分子会经过一系列的修饰和加工,最终成熟为mRNA分子。
mRNA分子可以进一步参与到蛋白质的合成过程中。
总结起来,真核生物转录起始复合物的形成过程是一个复杂而精确的调控过程。
转录因子和RNA聚合酶相互作用,通过结合到启动子区域,形成稳定的结构。
转录因子的选择性结合和RNA聚合酶的合成和解离,共同协调着基因的转录过程。
真核生物基因的转录过程和调控方式

真核生物基因的转录过程和调控方
式
真核生物基因的转录过程是指将DNA作为模板,通过RNA聚合酶识别和复制DNA序列并转换成mRNA,从而实现基因信息从DNA到RNA的传递。
真核生物基因的调控方式包括:
1、剪接调控:DNA的剪接可以改变基因的表达,这是由DNA片段的延伸或切断决定的。
2、启动子调控:启动子位于基因上游,它可以调节RNA聚合酶对该基因的转录程度。
一般来说,调控元件如转录因子结合到启动子位点,能够使基因的转录更加有效。
3、调控元件调控:调控元件可以结合到基因的启动子位点,从而改变其转录水平,也可以结合到基因的终止子,从而影响到基因的表达量。
4、miRNA调控:miRNA是一种小RNA分子,它可以与mRNA的3'UTR结合,影响mRNA的翻译或降解。
真核rna聚合酶2的转录过程

真核rna聚合酶2的转录过程
真核生物RNA聚合酶Ⅱ的转录过程包括以下几个步骤:
- 启动子的识别:全酶识别启动子序列。
- 转录起始复合体的形成:全酶与启动子形成闭合复合物,并进一步形成开放复合物。
- 起始:与最初的两个NTP结合,形成RNA聚合酶、DNA和新生RNA三元复合物。
- 延伸:尽快释放$\sigma$亚基,转录开始。
- 终止:转录完成。
真核生物RNA聚合酶Ⅱ的转录过程是一个复杂的过程,受到多种因素的调控。
如果你想要了解更多关于真核生物RNA聚合酶Ⅱ转录过程的信息,建议你查阅相关的生物学书籍或文献。
分子生物学--真核生物转录起始 ppt课件

PPT课件
6
核基质
核基质(nuclear matrix),细胞核内除核膜、染色质、核 仁和核孔外的骨架结构。
PPT课件
7
核基质与染色质核基质连 接区域
常染色质中附着于基质上 的DNA区域,富含AT,称为 基质相关区域(matrixassociated region, MAR) 或骨架附着区(scaffold attachment region, SAR) 连接在核基质上的DNA环, 称为结构域。
PPT课件
35
PPT课件
26
顺式调控元件
启动子(promoter) 增强子(enhancer) 负调控元件—沉寂子(silencer) 绝缘子 其它顺式调控元件: 1)应答元件(responsive elements) 真核细胞中对某些特定的环境作出应答的基因,常具有 相同的顺式元件—应答元件. 应答元件能被在一些特定情况下表达的调控因子识别(又 称为可诱导的顺式调控元件/反式作用因子)。 27 PPT课件 2)转座元件
染色体DNA中一种顺式 作用元件,具有稳定染 色质疏松结构的作用 实际上一种增强子 一个调控结构域,可能 由一个绝缘子、一个 LCR、一个核基质附着 点(MAR)和单个或多 个转录单位组成。
PPT课件 15
核小体重建—能量依赖
PPT课件
16
组蛋白修饰
乙酰化
乙酰化活化基 因组;去乙酰 沉默基因组
甲基化
沉默基因组
磷酸化 泛素化
PPT课件 17
PPT课件
18
DNA甲基化
DNA甲基化和组蛋白甲基化是非活性染色质的 特征 这两类甲基化常相互关联 大多数DNA甲基化的位点是CpG岛
62第六讲_Lecture_2_真核细胞的转录过程-讲义R
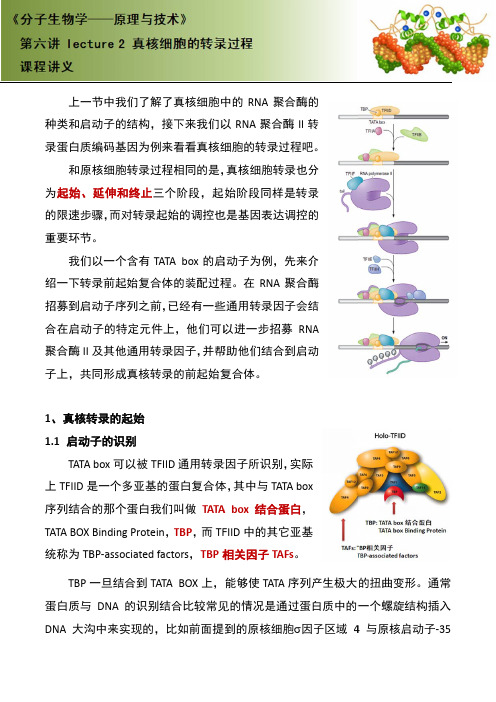
上一种类和启录蛋白质和原为起始、的限速步重要环节我们绍一下转招募到启合在启动聚合酶II 子上,共1、 真核转1.1 启动TATA 上TFIID 是序列结合TATA BOX 统称为TB TBP 蛋白质与DNA大沟一节中我们启动子的结质编码基因原核细胞转延伸和终步骤,而对转节。
们以一个含转录前起始启动子序列动子的特定及其他通共同形成真转录的起始子的识别A box 可以是一个多亚合的那个蛋Binding P BP ‐associa 一旦结合与DNA 的沟中来实现们了解了真结构,接下因为例来看转录过程相终止三个阶转录起始含有TATA 始复合体的列之前,已定元件上,通用转录因真核转录的始别以被TFIID 通亚基的蛋蛋白我们叫Protein ,T ated facto 合到TATA 的识别结合现的,比如真核细胞中下来我们以看看真核细相同的是,阶段,起的调控也box 的启的装配过程经有一些他们可以因子,并帮的前起始复通用转录蛋白复合体叫做TATA TBP ,而TF ors ,TBP 相BOX 上,合比较常见如前面提中的RNA 以RNA 聚细胞的转录真核细胞始阶段同也是基因表启动子为例程。
在RN 些通用转录以进一步帮助他们结复合体。
录因子所识体,其中与A box 结合FIID 中的其相关因子能够使TA 见的情况是到的原核聚合酶的聚合酶II 录过程吧胞转录也分同样是转录表达调控的例,先来介NA 聚合酶录因子会结招募RNA 结合到启动识别,实际TATA box合蛋白,其它亚基TAFs 。
ATA 序列产是通过蛋白核细胞因的转。
分录的介酶结A动产生极大的白质中的一子区域4的扭曲变一个螺旋与原核启变形。
通常旋结构插入启动子‐35常入5区的识别双螺旋的TBP 个平台。
在TBP 之的相互作也就是所方向发生再接被募集到得最后两动子上,包启动子就 别,而TBP的小沟中,结与TATbo接着TFII之后被募集作用是非对所有其他蛋生。
真核生物的转录过程
异的组织细胞或受到一些类固醇激素、生长
因子或其他因素刺激后,开始表达某些特异
蛋白质分子。
普遍转录因子
•普
有TFⅡA、TFⅡB、TFⅡD、
TFⅡE、TFⅡF、TFⅡH、TFⅡJ等多种。
TFⅡD由1个TATA盒结合蛋白(TBP)和多
个TBP结合因子(TAF)组成
有解链酶、ATP酶和蛋白激酶的活性, 它能使RNA聚合酶Ⅱ最大的亚基的羧基端 功能域(CDT)磷酸化。
RNA聚合酶Ⅰ的转录起始
转录的产物是
。
• RNA转录起始先由UBF1与核心启动子及 UCE中的G-C丰富序列结合,使这两部分靠 拢,然后SL1加入并与UBF1结合,组成转 录起始前复合物,随后RNA聚合酶Ⅰ与SL1 中的TBP结合形成起始复合物并起始转录。
• UBF1和SL1为上游结合因子和选择因子1
真核生物的转录过程
RNA聚合酶
• RNA聚合酶:真核生物中已发现有4中 RNA聚合酶,分别为 ,它 们专一性地转录不同的基因,因此由它们催 化的转录产物也各不相同。
• 原核生物靠RNA聚合酶就能完成从起始、
延长、终止的转录全过程,真核生物转录除
需RNA聚合酶外还需要另一些称为转录因
子的
参与转录的全过程。
延长阶段
• 转录起始复合物形成后,在位的RNA聚合 酶即开始依照碱基配对关系,按模板链的碱 基序列,从5'→3'方向逐个加入核糖核苷酸。
终止阶段
• 真核生物mRNA的3'端有poly(A)尾,这是
转录后才加进去的。在结构基因最后一个外
显子的3'端常有一组共同序列AATAAA,
其下游还有相当多的GT序列。这些序列称
:真核知之 甚少。
转录
2. 转录起始过程 •RNA聚合酶全酶 α2ββ′σ 与模板结合 σ发现识 聚合酶全酶(α ββ′σ)与模板结合 与模板结合, 聚合酶全酶 别位点,全酶与启动字-35序列结合 序列结合, 别位点,全酶与启动字 序列结合,随即酶移 序列, 向-10序列,跨入转录起始点; 序列 跨入转录起始点; •DNA双链解开 双链解开 •在RNA聚合酶作用下发生第一次聚合反应,形 在 聚合酶作用下发生第一次聚合反应, 聚合酶作用下发生第一次聚合反应 成转录起始复合物 5′-pppG -OH + NTP → 5′-pppGpN - OH 3′ + ppi ′ ′ ′ 3. 转录起始复合物 转录起始复合物: RNApol (α2ββ′σ - DNA - pppGpN- OH 3′ α ββ′σ) ′
-50
-40
-30
-20
-10
1
10
3. 真核生物的启动子结构
结构基因
-GCGC---CAAT---TATA
转录起始
-25 -75
增强子
TATA盒 盒 CAAT盒 盒 GC盒 盒
第二节 转录过程
一、原核生物的转录过程 (一)转录起始 1. 转录起始需解决的问题: 转录起始需解决的问题: • • RNA聚合酶必须准确地结合在转录模板的 聚合酶必须准确地结合在转录模板的 起始区域。 起始区域。 DNA双链解开,使其中的一条链作为转录 双链解开, 双链解开 的模板。 的模板。
编码链 模板链
mRNA
蛋白质
二、RNA聚合酶 聚合酶 (一)原核生物的RNA聚合酶 原核生物的 聚合酶
1. 聚合酶的组成
亚基 α β β′ σ
分子量 36512 150618 155613 70263
转录
• 32、在转录延长中,RNA聚合酶与DNA 模板的结合是 • A.全酶与模板结合 • B.核心酶与模板特定位点结合 • C.结合状态相对牢固稳定 • D.结合状态松弛而有利于RNA聚合酶向 前移动 • E.和转录起始时的结合状态没有区别
• 33、下列哪一序列能形成发夹结构? • A. A A T T A A A A C C A G A G A C A C G • B. T T A G C C T A A A T C A T A C C G • C. C T A G A G C T C T A G A G C T A G • D. G G G G A T A A A A T G G G G A T G • E. C C C C A C A A A T C C C C A G T C
第一节 模板和酶
大肠杆菌RNA聚合酶组分
亚基
α β β`
σ (singma)
分子量
36 512 150 618 155 613
70 263
功能
决定那些基因被转录
与转录全过程有关(催化) 结合DNA模板(开链)
辨认起始点
生物化学
第十二章 RNA的生物合成(转录)
第一节 模板和酶
(二) 真核生物的RNA聚合酶
• • • • • •
39、ρ因子的功能是 A.结合阻遏物于启动区域处 B.增加RNA的合成速率 C.释放结合在启动子上的RNA聚合酶 D.参与转录的终止过程 E.允许特定的转录启动过程
• 31、关于DNA指导RNA合成的叙述中哪 一项是错误的? • A.只有DNA存在时,RNA聚合酶才能催 化生成磷酸二酯键 • B.转录过程中RNA聚合酶需要引物 • C.RNA链的合成方向是5`→3`端 • D.大多数情况下只有一股DNA链作为 RNA合成的模板 • E.合成的RNA链没有环状的
转录ppt(共66张PPT)
RNA 聚合酶
Ⅰ
Ⅱ
Ⅲ
转录
产物
加工
45SrRNA
18SrRNA 28SrRNA
hnRNA tRNA 5SRNA
snRNA
mRNA
利福平
抑制原核生物的RNA聚合酶
鹅膏蕈碱
抑制真核生物的RNA聚合酶
利福平 主要用于治疗结核病、麻风病等
鹅膏蕈碱 鬼笔鹅膏
三、真核生物的转录产物为单顺
反子
与原核生物不同,真核生物一个转 录单位仅生成一个mRNA分子,经翻 译生成一条多肽链
(三)都形成3’, 5’-磷酸二酯键
二、复制与转录的不同点
模板
原料
酶
校读
复制 两条链 dNTP DNA聚合酶
有
转录 一条链
NTP RNA聚合酶
无
配对
产物
引物
复制
A=T GC
子代双
链DNA
需要
转录
A=U T=A
GC
mRNA tRNA rRNA
不需要
第一节 原核生物的转录
一、转录模板 二、RNA聚合酶 三、模板和酶的辨认结合
第 11 章
RNA的生物合成(转录)
DNA DNA DNA
RNA
蛋白质
中心法则
目录
前 言 复制与转录的异同 第1节 原核生物的转录
第2节 真核生物的转录特点
第3节 真核生物转录后加工
前言 复制与转录的异同
一、相同点 二、不同点
一、复制与转录的相同点
(一)服从碱基配对规则
(二)合成方向都是5’-3’
DNA-dependent RNA polymerase (二)合成方向都是5’-3’ (四)腺嘌呤脱氨成为次黄嘌呤 σ亚基从三元起始复合物上脱落后,核心酶的构象随之发生改变,并沿着模板链的3’→5’方向滑行,进入延长阶段 第1节 原核生物的转录 原核生物的RNA聚合酶仅1种,合成全部RNA 主要用于治疗结核病、麻风病等 真核生物rRNA转录后加工 α2 β β’ ω σ称为全酶