遗传学第五章
遗传学第五章 遗传物质的分子基础
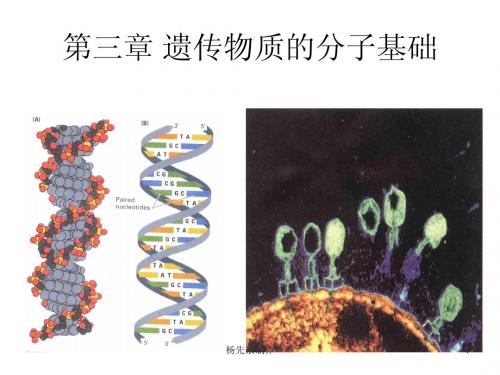
DNA双螺旋结构模型的意义 2. DNA双螺旋结构模型的意义
• DNA双螺旋模型结构同时表明: DNA双螺旋模型结构同时表明: 双螺旋模型结构同时表明 –DNA复制的明显方式 DNA复制的明显方式 半保留复制。 DNA复制的明显方式——半保留复制。Waston 半保留复制 Crick在1953年就指出 DNA可以按碱基互补 年就指出: 和Crick在1953年就指出:DNA可以按碱基互补 配对原则进行半保留复制。 配对原则进行半保留复制。而在此之前对复制 方式人们一无所知。 方式人们一无所知。 –基因和多肽成线性对应的一个可能的理由: 基因和多肽成线性对应的一个可能的理由: 基因和多肽成线性对应的一个可能的理由 DNA核苷酸顺序规定该基因编码蛋白质的氨基 DNA核苷酸顺序规定该基因编码蛋白质的氨基 酸顺序;DNA中的遗传信息就是碱基序列 中的遗传信息就是碱基序列; 酸顺序;DNA中的遗传信息就是碱基序列;并 存在某种遗传密码(genetic code), 存在某种遗传密码(genetic code),将核苷酸 序列译成蛋白质氨基酸顺序。 序列译成蛋白质氨基酸顺序。 • 在其后的几十年中,科学家们沿着这两条途径前 在其后的几十年中, 探明了DNA复制、 DNA复制 杨先泉制作 进,探明了DNA复制、遗传信息表达与中心法则等 17 内容。 内容。
杨先泉制作 6
1)DNA(脱氧核糖核酸) DNA(脱氧核糖核酸) a、脱氧核糖 腺嘌呤( )、胞嘧啶 胞嘧啶( )、鸟嘌呤 鸟嘌呤( )、胸腺 b、腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)、胸腺 嘧 啶(T) 双链、 c、双链、长 RNA(核糖核酸) 2)RNA(核糖核酸) a、核糖 腺嘌呤( )、胞嘧啶 胞嘧啶( )、鸟嘌呤 鸟嘌呤( )、尿嘧啶 b、腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)、尿嘧啶 (U) 单链、 c、单链、短 2、分布 DNA主要存在于细胞核的染色体上 主要存在于细胞核的染色体上, DNA主要存在于细胞核的染色体上,少量存在于细胞质的 叶绿体和线粒体中; RNA主要存在于细胞质和细胞核的核仁 叶绿体和线粒体中; RNA主要存在于细胞质和细胞核的核仁 少量存在于染色体上。 上,少量存在于染色体上。
遗传学 第五章 连锁遗传

3.距离较远的两个基因间的双交换 4.邻近的两个交换位置具有干涉。
生命科学学院
怎样判断两个基因之间是连锁遗传还是独立遗传?
在F1的测交后代中,亲本性状组合大于50%,重新组 合型小于50%的为连锁遗传。
若亲本性状组合与重新组合型等于50%,测交后代 呈1:1:1:1的比例,则为独立遗传。
生命科学学院
连锁类型
一.完全连锁
同一连锁群上的基因在配子形成过程中作为一个整体, 随 染色体传递下去,而不发生染色体片段的交换的遗传现象。
果蝇:灰身B,黑身b,长翅V,残翅v
P ♀灰身残翅×♂黑身长翅
BBvv ↓ bbVV
F1
♂灰身长翅×♀黑身残翅
BbVv ↓ bbvv
灰身残翅 黑身长翅
Bbvv bbVv
卡平方X2检测
①明确理论假说 ②求卡方值
χ2=
(O
E E
)
2
(O是实际数,E是理论数)
③求自由度 总项数-1=(n-1) ④据卡方和自由度查卡方表得P值 ⑤确定理论与实验的相符程度(P值标准)
P≥0.05 成立-----实际结果符合理论 P<0.05 理论不能解释实际实验的结果 ⑥结论
生命科学学院
生命科学学院
2.断裂融合学说:
1937年,英国细胞学家C.Darlinton提出。
联会时同源染色体相互缠绕,染色体内的扭力和 染色体间的扭力平衡。
当同源染色体分开时,染色单体受应力的作用而 断裂。
断头松开螺旋后,断头不与原来的染色单体相接,
而与其同源非姊妹染色单体的断面相接,造成了
染色体片段的交换。
遗传学

第五章连锁遗传和性连锁(一) 名词解释:1.交换:指同源染色体的非姊妹染色单体之间的对应片段的交换,从而引起相应基因间的交换与重组。
2.交换值(重组率):指同源染色体的非姊妹染色单体间有关基因的染色体片段发生交换的频率。
3.基因定位:确定基因在染色体上的位置。
主要是确定基因之间的距离和顺序。
4.符合系数:指理论交换值与实际交换值的比值,符合系数经常变动于0—1之间。
5.干扰(interference):一个单交换发生后,在它邻近再发生第二个单交换的机会就会减少的现象。
6.连锁遗传图(遗传图谱):将一对同源染色体上的各个基因的位置确定下来,并绘制成图的叫做连锁遗传图。
7.连锁群(linkage group):存在于同一染色体上的基因群。
8.性连锁(sex linkage):指性染色体上的基因所控制的某些性状总是伴随性别而遗传的现象,又称伴性遗传(sex-linked inheritance)。
9.性染色体(sex-chromosome):与性别决定有直接关系的染色体叫做性染色体。
10.常染色体(autosome):性染色体以外其他的染色体称为常染色体。
同配性别11.限性遗传(sex-limited inheritance):是指位于Y染色体(XY型)或W染色体(ZW型)上的基因所控制的遗传性状只限于雄性或雌性上表现的现象。
12.从性遗传(sex-influenced inheritance):常染色体上基因所控制的性状,在表现型上受个体性别的影响,只出现于雌方或雄方;或在一方为显性,另一方为隐性的现象。
13.交叉遗传:父亲的性状随着X染色体传给女儿的现象。
14.连锁遗传:指在同一同源染色体上的非等位基因连在一起而遗传的现象。
(二) 是非题:1.雄果蝇完全连锁是生物界少见的遗传现象。
这仅指X染色体上的连锁群而言。
因为它的X染色体只有一条,所以,不会发生交换。
(-)2.基因连锁强度与重组率成反比。
(+)3.基因型+ C/Sh +的个体在减数分裂中有6%的花粉母细胞在Sh和C之间形成一个交叉,那么,所产生的重组型配子++和 Sh C 将各占3%。
【遗传学】第五章 染色体和连锁群

第五章染色体和连锁群本章重点一、连锁遗传:二对性状杂交有四种表现型,亲型多、重组型少;杂种产生配子数不等,亲型相等、重组型相等。
二、连锁和交换机理:粗线期交换、双线期交叉,非姐妹染色体交换。
三、交换值及其测定:重组配子数/总配子数;测交法测定,也可用F2 材料进行估计。
四、基因定位和连锁遗传图:确定位置、距离,基因位于染色体上;二点测验、三点测验;连锁群、连锁遗传图。
学时:91900年孟德尔遗传规律重新发现以后,生物界广泛重视,进行了大量试验。
其中有些属于两对性状的遗传结果不符合独立分配规律→摩尔根以果蝇为材料进行深入细致研究→提出连锁遗传规律→创立基因论→认为基因成直线排列在染色体上,进一步发展为细胞遗传学。
第一节连锁和交换一、连锁(一)连锁现象的发现1906年,贝特生(Bateson W.)和贝拉特(Punnett R. C.)在香豌豆的二对性状杂交试验中→首先发现性状连锁遗传现象。
第一个试验:P 紫花、长花粉粒(PPLL)×红花、圆花粉粒(ppll)↓F1紫、长PpLl↓⊗F2紫、长紫、圆红、长红、圆P_L_P_ll ppL_ppll总数实际个体数4831390393 13386952按9:3:3:1推算3910.51303.5 1303.5434.56952上述结果进行X2检验时,X2=3371.58,说明实计数与预计数差异极其显著,不可能由随机误差造成,应作重复实验。
以上结果表明F2 :①. 同样出现四种表现型;②. 不符合9:3:3:1;③. 亲本组合数偏多,重新组合数偏少(与理论数相比)。
第二个试验:P紫花、圆花粉粒(PPll)×红花、长花粉粒(ppLL)↓F1 紫、长PpLl↓⊗F2 紫、长紫、圆红、长红、圆P_L_P_ll ppL_ppll总数实际个体数22695971419按9:3:3:1推算235.878.578.526.2419结果与第一个试验情况相同。
《遗传学》第五章基因突变:第四节 基因突变的筛选与鉴定

◆碱基类似物: ◆碱基修饰物: ◆DNA插入剂:
47
(一)碱基类似物
◆5–溴尿嘧啶(5BU),5—溴去氧尿核苷、2—氨 基嘌呤等 在DNA复制时引起碱基配对上的差错 ☆转换(transition) : ☆颠换(transversion:
48
49
5–溴尿嘧啶
◆ 5–溴尿嘧啶类似胸腺嘧啶
玉米籽粒胚乳:非甜(Su)甜(su)
P:
甜粒亲本(susu)×非甜粒亲本(SuSu)
诱变处理
G:
su
Susu
F1:
Susu(非甜)
susu(甜粒)
正常花粉粒后代 突变花粉粒后代
亲本配置:具有花粉直感特征;性状显隐性差异明显; 亲本具有相对性
13
1个分蘖A-a突变 (5)其他未突变穗
AA Aa
AA
(1)突变穗 (2)2个穗行
9
(2)突变显隐性的鉴定 隐性突变可利用杂交试验加以鉴定 例如: 高杆——矮杆:隐性? 杂交 F1 显性?
10
(三)突变率的测定
体细胞突变频率一般是根据M2出现的突 变体比例估算。例如,在M2群体的10万 个个体数中出现5个突变体,突变频率为 5×10-5。
11
2. 斯特得勒花粉直感
12
花粉直感法测定突变(诱变)率
碱基烷基化 ◆烷化剂? 去烷基化(dealkylation)
酶
烷基转移酶 (alkyltransferase) 甲基转移酶 (methyltranferase)。
34
3. 单链断裂修复
DNA单链断裂
单链的的化学键?
DNA 连 接 酶 催 化 双 螺
旋结构中单链断裂处
形成磷酸二酯键
遗传学第五章

QuickTime?and a decompressor are needed to see this picture.
28, 0, 5
人和酵母的基因组比较
2.2 在核基因组中基因是如何组织的
C. 其他真核生物中的基因编排 • 可以建立一个假设:越是复杂的真核生物,它们的 基因组越不紧凑。
根据基因编 码的蛋白质 的结构域推 测的功能, 对基因进行 分类:
p216
酵母、拟南芥、线虫、果蝇和人基因种类的比较
2.3 基因组中有多少基因,它们的功能又是什么
C. 基因家族 • 具有相同或相似序列的基因家族是许多基因组的共 同特征。
• rRNA 基因属于最经典的多基因家族的例子,这些 家族是基因扩增产生的。
• 串联重复DNA也叫卫星DNA ,因为在用密度梯度离心法 分离基因组 DNA时,含有串 联重复序列的 DNA片段会形 成“卫星”带。
由重复序列组成
由单拷贝序列组成
• 如将人 DNA 截断成 50-100kb 的片段,离心后会形成一个 主带和三个卫星带。
人类基因组卫星DNA
2.4 真核生物基因组中的重复DNA序列
• 假基因是一种进化遗迹,表明基因组处于持续性变 化的过程中。
2.3 基因组中有多少基因,它们的功能又是什么
D. 假基因和其他进化遗迹 主要存在两种假基因: • (1)常规假基因:指基因的核苷酸序列因为突变而发 生改变,导致基因失活。如珠蛋白假基因。 • (2) 已加工的假基因:以某个基因的 mRNA 合成的 cDNA 拷贝再次插入到基因组后,由于缺少上游调 控序列不能转录而引起的基因失活。
A. 人类基因目录
• 在 人 的 3-4 万 个 基 因 中 , 约半数基因的功能是已知 的,其中绝大多数是蛋白 质编码基因,只有不到 2500个基因是编码不同类 其他多种活性 型的功能性 RNA。 复合物出入细胞的转运、蛋白质
遗传学(第3版)第5章核外遗传分析

sm-r mt + sm-s mt - 全部后代(99%)表现为sm-r sm-s mt + sm-r mt - 全部后代(99%)表现为sm-s
以上结果表明正反交结果不同,说明衣藻对链霉素的抗性或 敏感是由非孟德尔遗传因子所控制(图5-3)
研究表明:该性状由衣藻中叶绿体基
因组DNA上的基因所控制。交配型mt+ 和mt–细胞质中都含有叶绿体基因组 DNA,当发生交配时,两个亲本对合 子贡献的细胞质是相等的,而后代总 是保留mt+亲本与叶绿体基因组上控 制的性状有关的表型。一种分子解释
某些产物积累在卵母细胞的细胞质中,使子代表型不由自身 的基因型所决定而出现与母体表型相同的遗传现象,则称为 母体影响。 母体影响有两种: 一种是短暂的,只影响子代个体的幼龄期;
5.1.3衣藻抗生素抗性的遗传
衣藻通常行无性生殖,有时通过两种形态相同但交配型
不同的配子进行融合,行有性生殖。同型配子间不能融合,
仅相反交配型的配子能够融合产生二倍体的合子。衣藻的交 配型是由细胞核内一对等位基因mt +和mt -所决定的。配子
融合给合子提供了相等的细胞内含物,合子萌发时,通常立
即发生减数分裂,四个减数分裂的产物作为非顺序四分子在 一个子囊中,其中有两个mt +细胞和两个mt -细胞,即核基
②自体受精(Autogamy): 同一个体的两个小核经减数分裂,留下一个小核, 该小核分裂一次又相互合并,随后再分裂发育成小核和大核。 (Fig--45 ) 3).草履虫放毒型和敏感型的接合 若 某些草履虫 草履虫素 杀死其它的草履虫但对自己无害。 能产生草履虫素的个体叫做放毒型 而受害的个体就称为敏感型 根据遗传学实验,草履虫的放毒型必需有两种因子同时存在: ① 细胞质因子,叫做卡巴粒(Kappa particle , κ)
遗传学第五章 基因组
S.S. DNA
复性过程依赖于单链分子间的随机碰撞
( Depends on the collision of complementary S.S. DNA )
影响复性的因素:
• 温度
• 时间 • 离子强度 • DNA片段大小 • DNA序列复杂性
• DNA分子浓度
真核生物:
•第1组分(25%),快,高度重复序列; •第2组分(30%),中,中度重复序列;
序列能或不能被某一酶酶切,实际上相当于一对等位基因的差异。
• 如一对同源染色体二个 DNA分子,一个具有某种 酶的酶切位点,另一个无此 位点������ 酶切后形成的DNA片段长 度就会有差异,即多态性 (RFLP) ������ 根据该等位基因的遗传,将 RFLP作为标记定位在基因 组的某一位置上。
散在重复序列
散在重复序列:散在的方式分布于基因组内的重复序列。
短散在重复序列(SINEs),500bp
长散在重复序列(LINEs),1000bp
Alu序列家族:人类50-70万拷贝;
人和灵长类基因标志。 多聚(dT-dG)家族:10万拷贝
第二节 基因组研究
基因和基因组的结构 各种元件的序列特征 基因作图和基因定位 不同序列结构具有不同功能 基因表达的调控 基因与环境相互作用
•
•
(2) 简单序列长度多态性
(simple sequence length polymor-phisms,SSLP) • 简单序列长度多态性,又称为VNTR variable number tandem repeat 数 目可变的串联重复多态性。指重复单位相对较小,由重复单位的序列差异和 数目变化,可形成丰富的多态性。 包括:小卫星序列、微卫星序列 。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
紫花:红花(4831+390):(1338+393)=5221:1731≈3:1
长花粉:短花粉(4831+393):(1338+390)=5224:1728≈3:1
第二个试验:
紫花:红花(226+95):(97+1)=321:98≈3:1
长花粉:短花粉(226+97):(95+1)=323:96≈3:1
%48.21.81.848.2
上述结果证实F1产生的四种配子不等:
不是1:1:1:1 (25:25:25:25%)。其中(Ft)
亲本组合= (4032+4035)/8368×100% = 96.4%
重新组合= (149+152)/8368×100% = 3.6%
摩尔根(Morgan T. H.)等以果蝇为材料进行测交的结果:
*以上结果都受分离规律支配,但不符合独立分配规律。
*F2不符合9:3:3:1,则说明F1产生的四种配子不等。
可用测交法加以验证,∵测交后代的表现型种类以及比例可反映出F1配子的种类和比例。
利用测交法验证连锁遗传现象:
特点:连锁遗传的表现为:
♥两个亲本型配子数是相等,>50%;
♥两个重组型配子数相等,<50%。
二点测验、三点测验;
连锁群、连锁遗传图。
学时:9
1900年孟德尔遗传规律重新发现以后,生物界广泛重视,进行了大量试验。其中有些属于两对性状的遗传结果不符合独立分配规律→摩尔根以果蝇为材料进行深入细致研究→提出连锁遗传规律→创立基因论→认为基因成直线排列在染色体上,进一步发展为细胞遗传学。
第一节连锁和交换
第二个试验:
P紫花、圆花粉粒(PPll)×红花、长花粉粒(ppLL)
↓
F1紫、长PpLl
↓⊗
F2紫、长 紫、圆 红、长 红、圆
P_L_P_llppL_ppll总数
实际个体数22695971419
按9:3:3:1推算235.878.578.526.2419
结果与第一个试验情况相同。
F2
紫长
紫圆
红长
红圆8163ຫໍສະໝຸດ 2.25(O- E)2/E
216.68
640.19
635.99
1878.74
上述结果进行X2检验时,X2=3371.58,说明实计数与预计数差异极其显著,不可能由随机误差造成,应作重复实验。
以上结果表明F2:
①.同样出现四种表现型;
②.不符合9:3:3:1;
③.亲本组合数偏多,重新组合数偏少(与理论数相比)。
实际个体数483139039313386952
按9:3:3:1推算3910.51303.51303.5434.56952
F2
紫长
紫圆
红长
红圆
总数
O
4831
390
393
1338
6952
E
3910.5
1303.5
1303.5
434.5
6952
(O-E)2
847320.25
834482.25
829010.25
第五章染色体和连锁群
本章重点
一、连锁遗传:
二对性状杂交有四种表现型,亲型多、重组型少;
杂种产生配子数不等,亲型相等、重组型相等。
二、连锁和交换机理:
粗线期交换、双线期交叉,非姐妹染色体交换。
三、交换值及其测定:
重组配子数/总配子数;
测交法测定,也可用F2材料进行估计。
四、基因定位和连锁遗传图:
确定位置、距离,基因位于染色体上;
总数
O
226
95
97
1
419
E
235.8
78.5
78.5
26.2
419
(O-E)2
96.04
272025
342.25
635.04
(O- E)2/E
0.41
3.47
4.36
24.24
X2=32.40,证明它仍然显著不符合孟德尔规律。
从这两个实验的数据可以看出一种共同的倾向:即与自由组合定律所预期结果相比较,F2中性状的亲本组合类型远远多于重组组合的类型,这等于说,在F1杂种形成配子时两对基因可能发生的4种类型中,有更多保持亲代原来组合的倾向,而且这种倾向与显隐性无关。
一、连锁
(一)连锁现象的发现
1906年,贝特生(Bateson W.)和贝拉特(Punnett R. C.)在香豌豆的二对性状杂交试验中→首先发现性状连锁遗传现象。
第一个试验:
P紫花、长花粉粒(PPLL)× 红花、圆花粉粒(ppll)
↓
F1紫、长PpLl
↓⊗
F2紫、长紫、圆 红、长 红、圆
P_L_P_llppL_ppll总数
1.相引组:
玉米(种子性状当代即可观察):
有色、饱满CCShSh× 无色、凹陷ccshsh
↓
F1有色饱满 × 无色凹陷
CcShsh ccshsh
配子CShCshcShcshcsh
Ft CcShsh Ccshsh ccShsh ccshsh
有色饱满有色凹陷无色饱满无色凹陷总数
粒数403214915240358368
∴F1配子中总是亲本型配子(CSh和csh或pr+vg+和prvg)数偏多,重新组合配子(Csh、cSh或pr+vg和prvg+)数偏少。
2.相斥组:
玉米:
有色、凹陷CCshsh× 无色、饱满ccShSh
↓
F1有色饱满 ×无色凹陷
CcShshccshsh
配子CSh Csh cSh cshcsh
Ft CcShsh Ccshsh ccShsh ccshsh
连锁遗传:原来亲本所具有的两个性状,在F2连系在一起遗传的现象。
相引组:甲乙两个显性性状,连系在一起遗传、而甲乙两个隐性性状连系在一起的杂交组合。如:PL/pl。
相斥组:甲显性性状和乙隐性性状连系在一起遗传,而乙显性性状和甲隐性性状连系在一起的杂交组合。如:Pl/pL。
(二)连锁遗传的解释
试验结果是否受分离规律支配?
红眼长翅pr+pr+vg+vg+× 紫眼正常翅prprvgvg
↓
F1红眼长翅 ×紫眼正常翅
pr+prvg+vgprprvgvg
配子pr+vg+ prvg+ pr+vg prvgprvg
Ft pr+prvg+vgprprvg+vgpr+prvgvgprprvgvg
红眼长翅紫眼长翅红眼正常翅 紫眼正常翅总数
个数133915415111952839
结果:亲本组合=((1339+1195)/2839)×100%=89.26%
重新组合=((154+151)/2839)×100%=10.74%
∴证实F1产生的四种配子数不等,两种亲型配子多,两种重组型少,分别接近1:1。
亲本具有的两对非等位基因(Cc和Shsh或pr+pr和vg+vg)不是独立分配,而是连系在一起遗传,如C-Sh、c-sh或pr+vg+和prvg常常连系在一起。