第六章 信号转导--第二信使-案例
第六章 同化物的运输复习思考题及答案

第六章同化物的运输、分配及信号的传导(一)名词解释源(source) 即代谢源,是产生或提供同化物的器官或组织,如功能叶、萌发种子的子叶或胚乳。
库(sink) 即代谢库,是指消耗或积累同化物的器官或组织,如根、茎、果实、种子等。
共质体运输(symplastic transport) 物质在共质体中的运输称为共质体运输。
质外体运输(apoplastic transport) 物质在质外体中的运输称为质外体运输。
P蛋白(P-protein)即韧皮蛋白,位于筛管的内壁,当韧皮部组织受到损伤时,P-蛋白在筛孔周围累积并形成凝胶,堵塞筛孔以维持其他部位筛管的正压力,同时减少韧皮部内运输的同化物的外流。
转移细胞(transfer cells)在共质体-质外体交替运输过程中起转运过渡作用的特化细胞。
它的细胞壁及质膜内突生长,形成许多折叠片层,扩大了质膜的表面积,从而增加溶质内外转运的面积,能有效地促进囊泡的吞并,加速物质的分泌或吸收。
比集转运速率(specific mass transfer rate, SMTR) 单位时间单位韧皮部或筛管横切面积上所运转的干物质的数量。
韧皮部装载(phloem loading) 同化物从合成部位通过共质体或质外体胞间运输,进入筛管的过程。
韧皮部卸出(phloem unloading) 同化物从筛管分子-伴胞复合体进入库细胞的过程。
空种皮技术(empty seed coat technique,empty-ovule technique) 切除部分豆荚壳和远种脐端的半粒种子,并去除另半粒种子的胚性组织,制成空种皮杯。
短时间内,空种皮杯内韧皮部汁液的收集量与种子实际生长量相仿,此法适用于研究豆科植物的同化物运输。
源库单位(source-sink unit) 在同化物供求上有对应关系的源与库合称为源-库单位。
源强和库强源强(source strength)是指源器官同化物形成和输出的能力;库强 (sink strength) 是指库器官接纳和转化同化物的能力。
植物细胞胞内信号转导与第二信使系统

第三节 胞内信号系统 三、环核苷酸信号系统
第三节 胞内信号转导与第二信使系统
植物细胞第 二信使系统
钙信号系统 磷酸肌醇信号系统 环腺苷酸信号系统
此外,还有一些化学物质,如乙烯、多胺类化合物、H+、 H2O2等,也可能是细胞中的胞内信使。
一、 钙信号系统
作为第二信使的一个重要条件: 该因子能够响应胞外刺激信号的变化,进而调节细胞的生理
活动。
植物细胞内的游离钙离子已被许多研究工作证实是细胞信号 传导过程中的重要的第二信使。
钙在植物细胞中的分布极不平衡。细胞质中Ca2+浓度很低, 细胞器的Ca2+浓度是细胞质的几百甚至上千倍,而细胞壁是 最大的Ca2+库。
仍留在质膜上的DAG使蛋白激酶C(PKC)激活,催化蛋白质 磷酸化。Ca2+-CaM和蛋白质(酶)磷酸化导致细胞反应。
IP3产生后很快被磷酸二脂酶水解,首先产生肌醇-1,4-二磷酸 (IP2),继而是肌醇单磷酸(IP)及磷脂酰肌醇(PI)返回膜 中。⊕表示活化。
肌醇磷脂信号系统与Ca2+信使系统的关系:
几乎所有的胞外刺激信号都可能引起胞内游离Ca2+离 子浓度的变化,而这种变化的时间、幅度、频率、区 域化分布等却不尽相同,不同刺激信号的特异性正是 靠钙浓度变化的不同形式体现的。
Ca2+信号受体:
胞内Ca2+信号通过其受体-钙调蛋白传递信息。
植物中的钙调蛋白主要有三种: (1)钙调素(calmodulin,CaM); (2)钙依赖型蛋白激酶
当完成信号传递后,Ca2+被Ca2+泵或者 Ca2+/H+逆向转 运器 ,泵出胞外或者泵进胞内Ca2+库。胞质中的 Ca2+又回落到静止状态水平。同时Ca2+与受体蛋白 分离。
脱落酸介导气孔开闭的信号转导

胞质Ca2+浓度的增加 来源一是胞外Ca2+通 过质膜内流通道进入 胞质;二是Ca2+从细 胞内Ca2+库(如液泡) 释放到胞质。
ABA调节气孔保卫细胞胞质Ca2+浓度增加有两条途
径:一条是ROS(活性氧)途径;另一条为IP3cADPR(环化ADP核糖)途径。
第六章 植物信号转导
第五节 脱落酸介导气孔关闭的信号转导
1.简述ABA调节气孔保卫细胞Ca2+浓度增加 的ROS途径和IP3-CAPR途径的异同点。
第六章 植物细胞信号转导 第五节 脱落酸介导气孔开闭的信号转导
第六章 植物信号转导
主要内容:
第五节 脱落酸介导气孔关闭的信号转导
一、细胞对干旱胁迫信号的感知 二、脱落酸对气孔运动调节的信号转导
第五节
脱落酸介导气孔开闭的 信号转导
概述:
植物对干旱胁迫应答的起始点是细胞对干旱胁迫 信号的感知。
ABA是一种胁迫激素,参与植物干旱应答反应的许多生 理过程。
图6-11 气孔保 卫细胞中ABA 信号转导途径 模式图
⑥cADPR和IP3的升高激活了液泡膜上的其他钙通道,更多的 Ca2+从液泡释放出来;⑦胞内钙的升高阻断了质膜上的Kin+通道; ⑧胞内钙的升高促进了质膜上Clout-通道的打开,引起质膜去极 化;⑨质膜上的质子泵受到ABA诱导的胞内钙增加和胞内pH升 高的抑制,质膜进一步去极化;⑩膜的去极化活化了质膜上的 Kout+通道;(11)K+和阴离子首先跨膜从液泡释放至胞质中。离 子从细胞中的外流,诱导气孔关闭或抑制了气孔开放。
2.介导ABA信号的胞内信使及其转导途径
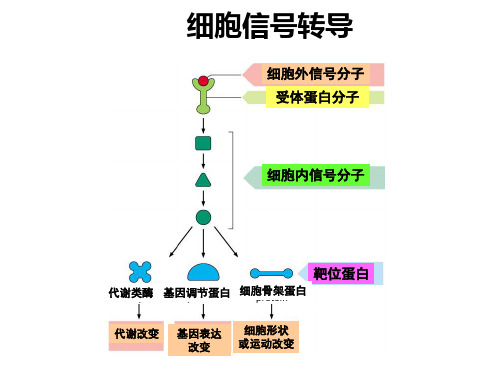
细胞信号转导

1基本概念信号转导signal transduction——细胞内外的信号,通过细胞的转导系统转换,引起细胞生理反应的过程。
化学信号chemical signals——细胞感受刺激后合成并传递到作用部位引起生理反应的化学物质。
物理信号physical signal——细胞感受到刺激后产生的能够起传递信息作用的电信号和水力学信号等物理性因子。
G蛋白G protein——全称为GTP结合调节蛋白(GTP binding regulatory protein),此类蛋白由于其生理活性有赖于三磷酸鸟苷(GTP)的结合以及具有GTP水解酶的活性而得名。
在受体接受胞间信号分子到产生胞内信号分子之间往往要进行信号转换,通常认为是通过G蛋白偶联起来,故G蛋白又称为偶联蛋白或信号转换蛋白。
第二信使second messenger——能被胞外刺激信号激活或抑制的、具有生理调节活性的细胞内因子。
第二信使亦称细胞信号传导过程中的次级信号。
在植物细胞中的第二信使系统主要是钙信号系统、肌醇磷脂信号系统和环核苷酸信号系统等。
动作电波action potential,AP——也叫动作电位,指细胞和组织中发生的相对于空间和时间的快速变化的一类生物电位,它是植物的一种物理信号,可通过输导组织传递。
钙调素calmodulin,CaM——是最重要的多功能Ca2+信号受体,为单链的小分子酸性蛋白。
当外界信号刺激引起胞内Ca2+浓度上升到一定阈值后,Ca2+与CaM结合,引起CaM构象改变。
而活化的CaM又与靶酶结合,使其活化而引起生理反应。
磷脂酰肌醇phosphatidylinositol,PI——亦称肌醇磷脂(lipositol),即其肌醇分子六碳环上的羟基被不同数目的磷酸酯化,PI为磷脂酰肌醇;PIP为磷脂酰肌醇-4-磷酸;PIP2为磷脂酰肌醇-4,5-二磷酸。
肌醇磷脂参与细胞胞内的信号转导。
肌醇-1,4,5-三磷酸inositol-1,4,5-triphosphate,IP3——植物细胞内信号分子,通过调节Ca2+浓度来传递信息。
6-2信号转导

受触及的含羞草小叶在 1至2 秒钟向下弯,这 是由于电波引发叶枕运 动细胞中大量的K+和 Ca+2转运,引起膨压改 变的结果
图17.14 Albizia pulvini 背侧和腹侧的运动细胞之间的离 子流调节了小叶的开放与闭合。 •
产生PIs
电信号通过后去子叶
伤害
怀尔登(Wildon)等用番茄做实验,指 出由子叶伤害而引起第一真叶产生蛋白酶 抑制物PIs的过程中,动作电位是传播的主 要方式。他们采取让电信号通过后马上就 除去子叶以及使子叶叶柄致冷以阻碍筛管 运输、排除化学物质传递的试验,其结果 都证明单有电信号就可以引起PIs反应,而 且他们也首次证明了电信号可引起包括基 因转录在内的生理生化变化。
第五节 植物体内的信号传导
生长发育是基因在一定时间、 重力 空间上顺序表达的过程,而基因表 达除受遗传信息支配外,还受环境 光合作用的光 的调控。 光周期 光形态建成的光 植物在整个生长发育过程中, 湿度 受到各种内外因素的影响,这就需 温度 草食动物 要植物体正确地辨别各种信息并作 风 出相应的反应,以确保正常的生长 和发育。 乙烯 例如植物的向光性能促使植物 病原体 向光线充足的方向生长,在这个过 程中,首先植物体要能感受到光线, 寄生虫 然后把相关的信息传递到有关的靶 土壤质地 细胞,并诱发胞内信号转导,调节 土壤微生物 水分状况 有毒物质 基因的表达或改变酶的活性 矿质营养 光质→光受体→ 光质→光受体→信号转导组分 →光调节基因→向光性反应 光调节基因→ 各种外部信号影响植物的生长发育
在多数情况下信号分 子与细胞表面的受体分子 结合。许多亲水的分子如 肽、碳水化合物和渗透的 信号不易通过质膜,而与 细胞表面的受体相结合 一般认为受体存在于 质膜上。然而植物细胞具 有细胞壁,它可能使某些 胞间信号分子不能直达膜 外侧,而首先作用于细胞 壁。一些外界刺激有可能 通过细胞壁—质膜—细胞 骨架蛋白变构而引起生理 反应。
植物细胞的信号转导-PPT课件

细胞内受体(intra cellular receptor):存在于亚细胞 组分(如细胞核等)的受体。
细胞表面受体(cell surface receptor):位于细胞质膜上 的受体。
细胞表面受体
➢酶联受体 (enzyme-linked receptor)
细胞的信号转导过程是一个级联放大的过程。
细 胞 信 号 传 导 的 主 要 分 子 途 径
?思考题
1、名词解释: 受体,G蛋白,CaM
2、问答题 植物细胞信号转导的大致途径是怎样的?
双信号系统
ABA引起气孔关闭机理的模 型
在这个模型中, ABA与受 体(R)结合,导致了Ca2+ 的输入或Ca2+从胞内钙库 中的释放,
(1.ABA使胞外Ca2+通过 Ca2+通道进入保卫细胞 ;2.IP3激活液泡和内质网膜 上的Ca2+通道开放,向胞质 释放Ca2+)
从而使细胞质中的Ca2+浓 度升高,促进了质膜上阴离 子与K+Out通道的开放,并 抑制了K+in通道的开放。当 离开细胞的离子比进入细胞 的多时,细胞就会失水,从 而使得气孔关闭。
➢钙调素(CaM)
一种钙受体蛋白,是耐热、酸性的小分子球蛋白,具有148 个氨基酸的单链多肽。其上有四个Ca2+结合位点。
作用方式:
直接与靶酶结合,诱导靶酶的活性构象,而调节靶酶的活性。 与Ca2+结合,形成活化态的Ca2+ CaM复合体,然后再与靶 酶结合将靶酶激活。CaM与Ca2+有很高的亲和力,一个CaM 分子可与4个Ca2+结合。
• 离子通道连接受体(ion-channel-linked receptor)
医学分子生物学第六章_信号转导
调节蛋白质功能 水平,调节细胞分化和增
和表达水平
殖
受体的结构特点
• 结合结构域-----识别外源信号分子并与之结 合
• 效应结构域-----转换配体信号,使之成为细 胞内分子可识别的信号
3、信号转导分子和分子开关
• 信号转导分子(signaling molecule):细 胞内执行信号转导的成分的一些蛋白质分 子和小分子活性物质。
• 信号转导分子组织在支架蛋白上的意义:
① 保证相关信号转导分子容于一个隔离而稳定的信号转导 通路内,避免与其他不需要的信号转导通路发生交叉反 应,以维持信号转导通路的特异性;
② 增加调控复杂性和多样性。
信号转导通路中的一些环节是由多种分子聚集形成的 信号转导复合物(signaling complex)来完成信号 传递的。
激酶
磷酸基团的受体
蛋白丝氨酸/苏氨酸激酶 蛋白酪氨酸激酶 蛋白组/赖/精氨酸激酶 蛋白半胱氨酸激酶 蛋白天冬氨酸/谷氨酸激酶
丝氨酸/苏氨酸羟基 酪氨酸的酚羟基 咪唑环,胍基,ε-氨基 巯基 酰基
蛋白磷酸酶衰减或终止蛋白激酶诱导的效应
• 蛋白质磷酸酶(phosphatidase)使磷酸化的 蛋白分子发生去磷酸化,与蛋白激酶共同 构成了蛋白质活性的调控系统。
及信息传递,是指一个细胞发出的信息通过介 质传递到另一个细胞并与靶细胞相应的受体相 互作用,然后通过信号转导产生胞内一系列生 理生化反应,最终表现为细胞整体的生物学效 应的过程。
T淋巴细胞
(一)细胞通讯的方式
靶细胞
细胞间隙连接
细胞表面分子接触通讯 可溶型信号分子
化学信号介导通讯
❖分泌化学信号
根据体内化学信号分子作用距离,可以将 其分为三类:
化学物质传递与信号转导

化学物质传递与信号转导化学物质的传递与信号转导是生命活动中十分重要的过程。
这一过程是指,化学物质在细胞内、细胞间或细胞与外界之间传递,并通过一系列化学反应最终转化成细胞内信息,控制细胞的行为。
典型的例子包括神经递质传递、荷尔蒙作用、信号转导等。
这些过程的理解对于学习和应用生命科学起到了非常重要的作用。
神经递质传递是一种为神经元提供信息的过程,涉及到神经元之间或神经元与细胞之间的传递。
神经递质主要有两种类型:兴奋性神经递质和抑制性神经递质。
前者能够刺激神经元发放冲动,后者则相反。
神经递质传递是一种基于电化学信号的过程,由神经元释放到突触间隙,通过密切接触的神经元或肌肉细胞感受到其中的化学物质,从而传达信息。
在此过程中,神经递质与受体之间的特定相互作用非常重要。
神经递质的作用可以通过多重学科的研究手段进行观察和解释。
例如,生理学家可以通过测量神经递质对细胞响应的影响来研究其功能,而化学家则立足于发掘神经递质的化学结构和制备新型递质类分子。
荷尔蒙是由内分泌细胞合成并分泌到全身循环系统中的化学物质。
荷尔蒙在人体内起到了非常重要的调节作用,例如控制新陈代谢、性发育、生殖和细胞生长等。
与神经递质传递主要依赖于电化学信号不同,荷尔蒙作用则主要依赖于化学信号的传递。
荷尔蒙与其靶细胞表面的受体相结合,从而启动一系列信号转导反应。
这一过程中,起着关键作用的是一个叫做“第二信使”的分子。
第二信使可以放大荷尔蒙的信号,增强细胞的响应。
另外,还有一些荷尔蒙通过阻止其他分子的合成或分解来影响细胞的活动。
荷尔蒙的合成、调节和应用领域都是医学和生物工程研究的重要课题。
信号转导是在化学和生物分子之间进行的一种信息传递过程。
这里的信号可以是物理刺激、荷尔蒙、神经递质等,而转导则是指将这些信号转化成细胞内的行为改变。
信号转导的过程包含了许多细节和理解难点,因而成为了生命科学中一个十分详细且研究深奥的方向。
目前,关于信号转导的研究涉及到许多不同的层次,从分子和生物物理学的层次,到分子生物学和细胞生物学的层次上。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
受 体
G 蛋 白
效 应 器
第 二 信 使
靶 酶 或 调 节 因 子
基因 表达 调控
长期 生理 效应
短期 生理 效应
跨膜信号传导
胞内信号传导
二、跨膜信号转换机制
1、受体(receptor)
2、G蛋白 偶联蛋白或信号转换蛋白接受 传导 反应 Nhomakorabea受体
激素 细胞壁
胞质 质膜
1、受体与信号的感受
• 受体(receptor) 是指在细胞质膜上或亚细胞组分中能与
……
生理反应
研究第二信使的方法
药理学实验
清除剂/抑制剂 激活剂
生理学实验
信号分子
水平测定
功能分析
分子遗传学
定性/定位 定量
三、胞内信号的转换
第二信使系统(second messengers)
将胞外刺激信号称作第一信使,由胞外信号激活或抑 制、具有生理调节活性的细胞内因子称第二信使。 (1). 钙信号系统 (2). 肌醇磷脂信号系统 (3). 环核苷酸 cAMP信号系统
保卫细胞中的细胞骨架
1 →3: from open to close 5 →7: from close to open
知识拓展---气孔运动信号转导
Munemasa S, Hauser F, Park J, Waadt R, Brandt B, Schroeder JI. Mechanisms of abscisic acid-mediated control of stomatal aperture. Curr Opin Plant Biol. 2015, 28:154-62
4
3.活化的α亚基继而 触发效应器(如磷酸脂 酶C) 把胞外信号转换 成胞内信号
4.而当α亚基所具有 的GTP酶活性将与α亚 基相结合的GTP水解为 GDP后,α亚基恢复到 去活化状态并与β和 γ亚基相结合为复合 体。
3
活化酶
异源三体G蛋白的活动循环
G蛋白参与的跨膜信号 的转换-cAMP
放大信号是指每个配体结合的受体可以激发多个G蛋白,每个G 蛋白激活一个腺苷酸环化酶,每个腺苷酸环化酶又可催化形成
受体
控制酶的合成 影响代谢过程
正化学信号与负化学信号
正化学信号(positive chemical signal) 随着刺激强度的增加,细胞合成量及 向作用位点输出量也随之增加的化学 信号物质。 负化学信号(negative chemical signal) 随着刺激强度的增加,细胞合成量及 向作用位点输出量随之减少的化学信 号物质。
IP3:肌醇-1,4,5-三磷酸
DAG:二酰甘油
3. 环核苷酸 cAMP信号系统?
是在活细胞内最早被发现的 一种重要的第二信使。
胞外刺激信号---动物细胞--受体--G蛋白---腺苷酸环化酶
---胞质内的cAMP水平,
cAMP作为第二信使进而调 节细胞的生理生化活动。
问题?
是否植物细胞也存在类似于动物细胞中的 cAMP第二信使系统?
第三节 植物细胞信号转导事例
ABA诱导气孔关闭—应用
半根灌溉法
信号 受体 蛋白激酶
质膜
激酶串联(如 蛋白激酶)
蛋白激酶
Ca+-结合蛋白 细胞骨架
第二信号
传导蛋白
代谢
蛋白激酶 通道 生长 渗透运动 细胞骨架 发育
转录因素
植物细胞中信号传导网内的相互关系
MAPK,激化细胞分裂的蛋白激酶
思考题
1.细胞信号传导研究方法有哪些?
无催化作用的 结构功能域 有催化作用的 结构功能域
2、G蛋白
•在受体接受胞间信号分子到产生胞内信号分子之间,通常 认为是通过 G 蛋白将信号转换偶联起来,故 G 蛋白又称偶联
蛋白或信号转换蛋白。
•G蛋白全称为 GTP 结合调节蛋白,其生理活性有赖于三磷酸 鸟苷(GTP)的结合以及具有GTP水解酶的活性。 吉尔曼(Gilman)与罗德贝尔(Rodbell)因此获得1994年诺贝 尔医学生理奖
(a)高等植物细胞中是否普遍存在cAMP; (b)生理浓度的cAMP是否可引起植物细胞的生理生 化效应; (c)植物细胞内是否存在合成和降解cAMP的酶; (d)植物细胞内是否存在cAMP的靶蛋白。
植 物 细 胞 信 号 转 导 过 程
细胞信号转导过程学习导图
胞外
受体
胞内
刺激/第一信使
第二信使
生理反应
改变细胞生长和代谢
18.2 各种内部信号影响植物细胞的代谢、生长和发育
植物细胞的信号转导
第一节 植物细胞信号转导概述 第二节 植物细胞信号转导过程 第三节 植物细胞信号转导事例
学习 导图
外界刺激
信号产生? 信号 分子
信号转导?
……
生理反应
学习重点和难点
受体、第二信使、G蛋白等概念。 植物把环境刺激信号转导为胞内反应的途径。 钙信使介导的信号传导途径。 ABA诱导气孔关闭的信号转导
干旱
ABA
CTK
正化学信号
负化学信号
2.物理信号
能够起传递信息作用的电信号。 电信号传递是植物体内长距离传递信息的一种重要方式。 外界刺激诱导细胞膜发生去极化, 当去极化水平达到阈值时,可激
活离子通道,进而引起细胞内外
离子浓度改变。 叶枕运动细胞中大量的K+和Ca2+ 转运,引起膨压改变。
3. 信号的传递
钙调蛋白(calmodulin,CaM)
钙调素是最重要的多功能 Ca 2+ 信 号受体,由 148 个氨基酸组成的
单链的小分子(分子量为17
000~19 000)酸性蛋白。 CaM分 子有四个Ca2+结合位点。 目前已知有十多种酶受Ca2+-CaM的调控,如多种蛋白激 酶、NAD激酶、H+-ATPase、Ca2+-ATP酶、Ca2+通道等。 生长素、光、摩擦等都可引起CaM基因活化,使CaM含量 增加。
大量的cAMP. 这样可使信号放大1000倍左右。
细胞信号传导
胞 外 刺 激 信 号
受 体
G 蛋 白
效 应 器
第 二 信 使
靶 酶 或 调 节 因 子
基因 表达 调控
长期 生理 效应
短期 生理 效应
跨膜信号传导
胞内信号传导
细 胞 信 号 传 导 的 主 要 分 子 途 径
学习 导图
外界刺激
信号产生? 信号 分子 信号转导?
信号物质特异性结合,并引发产生胞内次级信号的特殊成分。 与能受体结合的特殊信号物质称配体(Ligand)。 • 受体可以是蛋白质,也可以是一个酶系。
研究比较多受体
光受体 激素受体 激发子
(1)光受体(光形态建成)
植物体内至少存在三类光受体:
A.对红光和远红光敏感的光敏色素;
B.对蓝光和紫外光A敏感的隐花色素; C.对紫外光B敏感的紫外光受体。
2.研究细胞信号传导的模式植物主要有哪些?
颜色区表示Ca+2浓度,红的最高,蓝的最低
G蛋白-IP3介导的钙信号转导 存 在 问 题 是 ?
Ca信号传导途径:
a. 当细胞受到外界信号时,从质外体进入质膜内侧以及从 细胞内的钙库(内质网, 液泡)流向细胞液,使细胞液中 Ca2+量增大。 b. 中的钙受体蛋白与Ca2+结合,Ca2+受体复合体使一些功 能蛋白作用引起相应的生理生化反应。 c. 当完成信息传递后,Ca2+又迅速被泵出胞外或被胞内钙 库吸收,细胞液中Ca2+又回落到受刺激前的水平,Ca2+与 受体蛋白分离。
当环境信号的作用位点与效应位点在植物不同部位时, 胞间信号就要作长距离的传递。
植物之间的”对话”
(1)易挥发化学在体内气相的传递-茉莉酸甲酯
(2)化学信号的韧皮部传递-ABA,JA,SA和寡聚半乳糖
(3)化学信号的木质部传递-ABA
(4)电信号传递
(5)水力学信号传递
细胞信号转导过程
胞 外 刺 激 信 号
1.钙信号系统
Ca2+在植物细胞中的
分布极不平衡。 植物细胞质中Ca2+含 量在10-7~10-6mol· L-1, 而胞外Ca2+浓度约为104~10-3mol· L-1;胞壁是细
胞最大的Ca2+库(1~ 5mol· L-1);细胞器的
Ca2+浓度也是胞质的几百
到上千倍。
细胞质中开放的Ca+2通道附近Ca+2的分配
第一节 植物细胞信号转导概述 细胞信号转导
外界环境刺激因子和胞间信号分子等,作用于细胞表面或胞内 受体后,跨膜形成胞内第二信使,及经过其后的信号途径分级 联传递、诱导基因表达和引起细胞生理反应的过程。
胞 外 刺 激 信 号 靶 酶 或 调 节 因 子 基因 表达 调控 短期 生理 效应 长期 生理 效应
在质膜受体接受胞外信号 后,经G蛋白中介,由G-
蛋白激活磷酸酶C (PLC)
的水解作用形成两种信号 分子IP3和DAG
钙信使与双信使系统
IP3作用于液泡膜上的受体
后,在膜上形成Ca2+通道,
使Ca2+从液泡中释放出来, 引起胞内Ca2+水平增加, 启动胞内Ca2+信号系统,
调节和控制一系列的生理
反应。
(2)激素受体(植物生长物质)
乙烯受体 ABA受体
ABA受体研究故事
ABA受体
The flowering-time control protein FCA The Mg-chelatase H subunit ABAR the G proteincoupled receptor GCR2
FCA:F.A. Razem et al., Nature 439 (2006), pp. 290–294 ABAR:Y.Y. Shen et al.,, Nature 443 (2006), pp. 823–826 GCR2: X. Liu et al., Science 315 (2007), pp. 1712–1716