基因工程:第四章-酵母基因工程
基因工程第四章载体
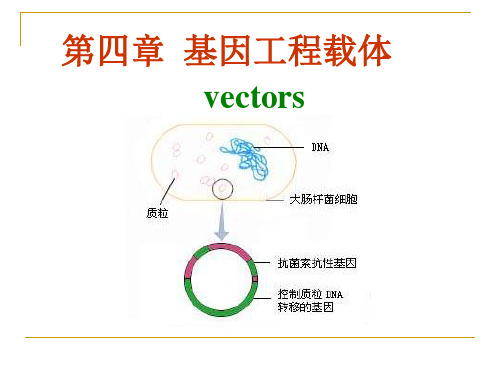
(4) 插入失活型质粒载体
载体的克隆位点位于其某一个选择性 标记基因内部。
如pDF41、pDF42、pBR329。
外源DNA
抗菌素抗性
无抗菌素抗性
(5)正选择的质粒载体 Direct selection vectors
直接选择转化后的细胞。
只有带有选择标记基因的转化菌细胞才 能在选择培养基上生长。
如pUR2、pTR262等。
目前通用的绝大部分质粒载体都是正 选择载体。
(6) 表达型质粒载体
主要用来使外源基因表达出蛋白质产物。
注意启动子的性质,终止子、起始 密码、终止密码的阅读正确。
如果在大肠杆菌里表达,必须把所克隆的 真核生物的基因置于大肠杆菌的转录—翻 译信号控制之下。
表达载体的结构
1)普通载体元件
b)细菌抗性原理 Ampr基因编码-内酰胺酶,特异地 切割氨苄青霉素的-内酰胺环。
ii)氯霉素(chloramphenicol,Cml)
a)抑菌原理 通过与50S核糖体亚基结合,干扰细胞 蛋白质的合成并阻止肽键的形成。杀死 生长的细菌。
b)细菌抗性原理
Cmlr 编码乙酰转移酶,特异地使氯霉 素乙酰化而失活。
(2)长度 6.3 kb。
(3)选择标记
大肠杆菌素(colicin)E1和对E1免疫 的基因(immE1)
① colicin E1基因的结构
cea 结构基因
imm
kil
免疫基因 溶菌基因
② 杀死不含有ColE1细菌的原因 cea + kil基因产物
③ 不被其他细菌的colicin E1所杀死的原因 imm基因
① 双抗菌素抗性选择标记 插入失活,分两次先后选择: 没有获得载体的寄主细胞 在Amp或Tet中都死亡。
酵母菌在基因工程中的应用

酵母菌在基因工程中的应用酵母菌是一类单细胞真核生物,是生物科学研究中的一种常见模式生物。
它们普遍存在于自然界中,可以在发酵食品的制备以及生命科学研究领域发挥着重要的作用。
在基因工程领域中,酵母菌更是被广泛应用,成为了基因工程领域的重要工具之一。
下面我们就来看看,酵母菌在基因工程领域中都有哪些应用吧。
一. 酵母菌作为表达宿主酵母菌是一类常见的蛋白表达宿主,能够快速高效地表达蛋白质,是一种常见的蛋白质产生工具。
一般来说,通过基因工程手段将需要表达的蛋白质的基因导入酵母菌中,利用其自身繁殖特性,迅速高效地表达出需要的蛋白质。
此外,在表达蛋白质的过程中,酵母菌的生长条件相对简单,可以通过温度、氧气、营养等因素的控制来实现高效的表达。
二. 酵母菌在药物研究中的应用当前,越来越多的药物研发都依赖于基因工程技术,而酵母菌则成为了药物研发中的重要工具之一。
通过将需要研发的靶点基因导入酵母菌中,可以模拟药物对生物体内靶点的作用过程。
此外,还可以通过酵母菌对药物副作用的研究,为药物的准确作用机制提供参考。
三. 酵母菌在癌症研究中的应用对于癌症的研究一直以来都是生物学家们所关注的重要问题之一。
而酵母菌则成为了癌症研究中的重要研究工具之一。
通过将癌症相关基因导入到酵母菌中,并通过对其复制、修复和细胞凋亡等过程的研究,可以更好地理解癌症的发生机制和治疗过程,为癌症的诊断和治疗提供更好的参考。
四. 酵母菌在基因组研究中的应用对于生命科学研究而言,基因组研究是一项重要的研究领域。
而目前,酵母菌的基因组研究也在不断地发展。
利用酵母菌基因组研究这一工具,可以揭示基因与生物型之间的关系,探寻基因突变造成遗传性疾病的可能机制,还可以帮助人们更好地理解基因间相互作用,促进基因工程技术的发展。
总之,随着基因工程技术的不断发展,酵母菌作为一种常见的模式生物,也在越来越多的领域中发挥着重要的作用。
通过其快速高效的蛋白表达能力以及对生物学过程的模拟研究,酵母菌为人们揭示了生物世界中的许多秘密。
第四章 基因工程的质粒载体

SC
2 质粒DNA的转移
(1)质粒的类型:在大肠杆菌中的质粒,可 以分为:
接合型质粒:能自我转移
具有自主复制的基因,控制细菌配对和质粒接合转 移的基因。
非接合型质粒 不能自我转移
按接合转移功 能分类
非接合型质粒
主要基因
自主复制基因,产生大肠杆菌素基因
按抗性记号 分类
Col质粒
接合型质粒
自主复制基因,抗菌素抗性基 因
第二代 酵母表达 穿梭质粒 体系
第三代 哺乳类细 病毒、脂质体 胞表达体系
第四代 基因直接 DNA本身 导入
细菌 酵母 培养动物细胞 生殖细胞、 体细胞、个体
(三)基因工程载体必须具备的条件:
※(1)有复制起点 ※(2)具有若干个限制性内切酶的单一识别位点 ※(3)具备合适的筛选标记 ※(4)具备合适的拷贝数目
(c)所示,F质粒无力帮助mob-突变体进行转移,其中F性须和转移装置虽已 形成,但ColE1 DNA并没有发生缺口。
(d)表示另一种具mob+表型并带有一个顺式显性突变的ColE1突变体,它缺 失了bom位点。在这样的寄主细胞中,虽然能够合成mob蛋白质,但由于不 能发生缺口,因此仍然不能够转移。
3.若质粒DNA经过适当的核酸内切 限制酶切割之后,发生双链断裂形成 线性分子(IDNA),通称L构型
第4章 载体的选择与构建

大多数自主转移质粒都有tra基因和oriT位点,它们在质粒 自主转移过程中起着重要作用。
tra 基因:大多数 tra 基因的产物与性菌毛的形成 (F 、 RP4 和 pKMl01 都 编码一根性菌毛 )、杂交对的形成有关;有些 tra基因编码作用于 oriT位 点的内切酶,使质粒中的一条 DNA链产生切口,从而开始转移;有些 tra基因则与质粒转移调控等有关。
pMB1, colE1 replicon 修饰的pMB1 replicon pSC101 pUB110 pE194 replicon
拷贝数15~20
宿主范围小:肠细菌
拷贝数500~700 宿主范围小:肠细菌(pUC系列) 拷贝数5 50 5 宿主范围广 多种革兰氏阴性菌 广 多种革兰氏阳性菌 广 多种革兰氏阳性菌
pSC101 replicon
1.4 质粒的不稳定性 ★
分离不稳定性:在细胞分裂过程中,有一个子细胞没有获得质粒 DNΑ
拷贝,并最终增殖成为无质粒的优势群体;
结构不稳定性:由转位作用和重组作用所引起的质粒 DNA的重排与缺失
质粒的分配方式: 主动分配 平均分配:每个子细胞刚好获得一半数目的质粒拷贝 配对位点分配:只有一对质粒呈主动分配,其余的是随机分配。 主动分配存在着有效的质粒拷贝数控制系统,从而保证了质粒的高度稳 定性 随机分配 分配不稳定,部分子细胞没有质粒,并在生长过程中具有优势而逐渐使 含质粒细胞比例越来越少
RBS,融合tag) 基因敲除质粒(筛选系统,目的基因的 同源片段) 辅助性质粒(例如提供位点特异性重组酶)
1.3 质粒的复制 ★
1.3.1 复制子(replicon) 包括复制起点(ori)、复制控制元件、复制蛋白编码基因的遗 传单元
酵母基因工程技术的综述与进展展望

酵母基因工程技术的综述与进展展望引言:酵母是一类常见的真核生物,广泛存在于自然界中。
由于酵母具有独特的细胞结构和代谢特性,成为许多科学研究的理想模型生物。
基因工程技术的发展使得研究者们能够通过编辑和改造酵母的基因组,来实现多种生物学和应用学的目标。
本文将对酵母基因工程技术的现状进行综述,并展望未来的发展前景。
一、酵母基因工程技术的发展历程酵母基因工程技术的研究始于20世纪70年代。
最早的酵母基因工程是通过改变酵母细胞的遗传背景,来研究基因功能。
而后,随着重组DNA技术的引入,酵母基因工程迅速发展起来。
1981年,科学家们成功地将人类基因插入到酵母细胞中,这是一个重大突破。
随后的几十年间,酵母基因组测序的完成以及基因敲除和基因重组技术的发展进一步推动了酵母基因工程技术的成熟。
二、酵母基因工程技术的应用领域1. 功能基因组学研究:通过酵母基因组的全面敲除和突变,可以研究基因的功能和相互作用。
这有助于更好地理解酵母细胞的生物学过程,也有助于揭示生物学中的一些基本原理。
2. 药物筛选和开发:酵母作为模型生物,在药物筛选和开发领域具有重要地位。
通过构建酵母表达外源蛋白的系统,可以进行大规模的化合物筛选,以寻找新的药物靶点和治疗方法。
3. 工业应用:酵母在生物技术和食品工业中具有广泛的应用。
例如,酵母可以被用于生产酒精、酵母提取物和酵母蛋白等。
通过基因工程技术改造酵母菌株,可以增加产量和改良产品的品质。
三、酵母基因工程技术的挑战与限制尽管酵母基因工程技术在许多领域中取得了显著进展,但仍然面临一些挑战和限制。
1. 基因组稳定性:酵母细胞往往会发生基因组重排和位点突变等现象,这导致基因敲除和基因重组等操作的结果不一致。
因此,在酵母基因工程中,确保基因组的稳定性仍然是一个关键问题。
2. 效率和选择性:目前的酵母基因工程技术中,基因敲除和基因重组等操作的效率相对较低,并且选择性也较差,这限制了其在实际应用中的广泛推广。
基因工程-第四章

(4) 插入失活型质粒载体 载体的克隆位点位于其某一个选择性标记基因内部。
抗菌素抗性
外源DNA 无抗菌素抗性
(5)正选择的质粒载体(Direct selection vectors)
直接选择转化后的细胞。只有带有选择标记基因的转化 菌细胞才能在选择培养基上生长。
目前通用的绝大部分质粒载体都是正选择载体。
各类载体
pBR322 外源基因Pst I
Tet中存活 但在Amp中死亡
外源基因BamH I Amp中存活 但在Tet中死亡
pBR322 外源基因Pst I
Tet中存活 但在Amp中死亡
外源基因BamH I Amp中存活 但在Tet中死亡
3. pUC系列
•University of California 的 J. Messing 和 J. Vieria 于1978年,在pBR322 的基础上改造而成,如 pUC7、pUC8、pUC9、pUC10、pUC11、pUC18、 pUC19。 • 元件来源
• 质粒空间构型与电泳速率
分子量相同的,scDNA最快、l DNA次之、ocDNA最 慢。
OC L
SC
质粒的生物学基本特性
1.自主复制性
• 质粒复制子是质粒 DNA 中能自主复制并维持正常拷贝数的 一段最小的核酸序列单位。
• 两部分组成:复制起始区(ori)及其相关的调控元件。 • 质粒能利用寄主细胞的 DNA 复制系统进行自主复制。 • 质粒 DNA 上的复制子结构决定了质粒与寄主的对应关系
必要的条件能力。
理想载体至少必备的条件
① 能在宿主细胞中自主复制 ②容易进入宿主细胞 ③ 容易插入外来核酸片段 ④ 容易从宿主细胞中分离纯化,便于重组操作 ⑤ 具有合适的筛选标记 ⑥ 具有针对受体细胞的亲缘性或亲和性
基因工程概要

基因工程概要第一章:绪论让幸福的细胞不凋亡;让开心的基因多表达;让健康的质粒常转染;让快乐的双链不变异;让烦恼的片段永封闭。
一、基因工程的基本定义基因工程是指重组DNA技术的产业化设计与应用,包括上游技术和下游技术两大组成部分。
上游技术指的是基因重组、克隆和表达的设计与构建(即重组DNA技术);而下游技术则涉及到基因工程菌或细胞的大规模培养以及基因产物的分离纯化过程。
二、四大里程碑遗传物质的明确——DNA;DNA双螺旋结构理论(半保留复制及其中心法则);基因遗传密码子的破译;基因转移载体的发现。
三、三大技术发明工具酶的发明:内切酶、合成酶、连接酶;基因合成和测序(合成仪、测序仪);PCR技术(PCR扩增仪)。
四、基因的现代概念移动基因;断裂基因;假基因;重复基因;重叠基因;或嵌套基因五、工具酶种类核糖核酸酶;脱氧核糖核酸酶; DNA连接酶;DNA聚合酶;RNA聚合酶;反转录;限制性核酸内切酶。
第二章:重组DNA技术基础1、DNA组成与结构:核酸、一级结构、二级结构和高级结构;2、RNA的组成与功能:mRNA、tRNA、rRNA.3、核酸的理化性质:(1)一般性质:核酸的溶解度.;酸碱性;核酸的高分子性质粘度: DNA>RNA dsDNA > ssDNA;核酸的紫外吸收(OD260)单核苷酸 > ssDNA(或RNA) > dsDNA;核酸的化学性质:核酸中的嘌呤和嘧啶能进行脱氨、聚合、烷基化等反应等。
(2)DNA的变性:DNA变性的本质是双链间氢键的断裂;(3)DNA的复性与分子杂交 : DNA复性(renaturation)的定义:在适当条件下,变性DNA的两条互补链可恢复天然的双螺旋构象,这一现象称为复性。
热变性的DNA 经缓慢冷却后即可复性,这一过程称为退火(annealing) ;减色效应: DNA复性时,其溶液OD260降低的现象.(4)核酸酶:核酸酶是指所有可以水解核酸的酶.(5)核酶:催化性RNA 作为序列特异性的核酸内切酶降解mRNA; 催化性DNA人工合成的寡聚脱氧核苷酸片段,也能序列特异性降解RNA。
基因工程 酵母单杂交技术的原理及应用

酵母单杂交是在酵母双杂交的基础上,20世纪90年年代中期又发展起来的--用于核酸和文库蛋白之间的研究。
在酵母单杂交系统中,省略了在酵母双杂交系统中采用的BD-X蛋白质杂交体,而用特异的DNA序列取代DNAGal4结合位点。
将已知的特定顺式作用原件构建到最基本启动子(Pmin)上游,把报告基因连接到Pmin下游。
编码待测转录因子cDNA与已知酵母转录激活结构域(AD)融合表达载体导入酵母细胞,该基因产物如果能够和顺式作用原件结合,就能激活Pmin启动子,使报告基因得到表达。
转录因子与顺式元件结合,激活最基本启动子Pmin,使报告基因表达,若连接如3个以上顺式作用元件,可增强转录因子的识别和结合效率。
优点:简单易行,无需分离纯化蛋白,酵母菌属于真真核生物,杂交体系检测到的与DNA结合的蛋白质是处于自然构象克服了体外研究时蛋白通常处于非自然构象的缺点,因而灵敏性很高。
缺点:有时由于插入的靶元件与酵母内源转录激活因子可能发生相互作用,或插入的靶元件不需要转录激活因子就可以激活报道基因的转录,因而存在假阳性结果。
如果酵母表达的AD杂合蛋白对细胞有毒性或者融合蛋白在宿主细胞内不能稳定的表达,或者融合蛋白发生错误折叠,或者不能定位于细胞核内,以及融合的GAL4-AD封闭了蛋白上与DNA作用的位点则都可能干扰AD杂合蛋白结合于靶元件的能力,从而产生假阴性的结果。
酵母单杂交系统应用:1. 鉴别DNA结合位点,并发现潜在的结合蛋白基因,目前对于酵母单杂交技术的应用主要体现在这方面。
Chew et al(1999)应用酵母单杂交技术证实了在大鼠脑中存在的COUP-TFⅠ、EAR2和NURR1等蛋白质GRIK5基因的内含子结合蛋白。
2. 对DNA结合结构域进行分析如果能得到DNA结合结构域的结构信息,就可以用酵母单杂交技术对该结构进行分析.Mak et al(1996)运用此技术测试哺乳动物具有基本的螺旋- 环- 螺旋(bHLH)结构的转录因子,通过对肌调节因子4(MRF4)的研究,证实其具有转录活性。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
UBC4-UBC5双突变型:
UBC4-UBC5双突变型能大幅度削弱泛
素介导的蛋白降解。
7个泛素连接酶基因的突变对衰减蛋白 降解作用同样有效。
6、内源性蛋白酶缺陷型的突变宿主菌
酿酒酵母具有20多种蛋白酶 空泡蛋白酶基因PEP4野生型和
pep4-3突变株
B-半乳糖苷酶活性明显升高
(三) 酵母菌的载体系统
酵母基因工程
酵母菌作为外源基因表达受体菌的特征 酵母菌的宿主系统 酵母菌的载体系统 酵母菌的转化系统 酵母菌的表达系统 利用重组酵母生产乙肝疫苗
1974 Clarck-Walker和Miklos发现在多数酿酒酵母 中存在质粒。
1978 Hinnen将来自一株酿酒酵母的leu2基因导入 另一株酿酒酵母,弥补了后者leu2的缺陷, 标志着酵母表达系统建立。
酵母菌有4个泛素编码基因:
UBI1 编码泛素-羧基延伸蛋白52 对数生长期表达 稳定期关闭
UBI2 编码泛素-羧基延伸蛋白52 对数生长期表达 稳定期关闭
UBI3 编码泛素-羧基延伸蛋白76 对数生长期表达 稳定期关闭
UBI4 编码泛素五聚体
对数生长期关闭 稳定期表达
酵母菌有7个泛素连接酶基因:
UBC1、UBC2、UBC3、UBC4、UBC5、UBC6、UBC7
酵母菌表达外源基因的优势: 全基因组测序,基因表达调控机理清楚,遗传 操作简便。 具有真核生物蛋白翻译后加工修饰系统。 能将外源基因表达产物分泌至培养基中。 大规模发酵工艺简单、成本低廉。
不含特异性病毒、不产毒素,被美国FDA认定为 安全的基因工程受体系统。
酵母菌表达外源基因的缺点:
表达产物的糖基化位点和结构特点 与高等真核生物有差距。
特点:
可利用甲醇作为唯一碳源; 表达水平高; 产物糖基化更合理。
其它酵母:
已有60多种酵母菌建立了转化系统。 乳酸克鲁维酵母(Kluyveromyces lactis) 非洲酒裂殖酵母(Schizosaccharomyces pombe)
酿酒酵母的遗传学和分子生物学 研究最详尽,但巴斯德毕赤酵母表达 外源基因最理想。
泛素降解途径衰减的酿酒酵母
UBI4缺陷型: 在酿酒酵母菌中,泛素主要由UBI4基因
表达。 UBI4突变株能正常生长,但细胞内游离
泛素分子的浓度比野生株低得多,是外源基 因表达的理想受体。
UBA1缺陷型:
UBA1编码泛素激活酶E1,是一种看 家基因。
UBA1突变株是致死性的,但其等位 基因缺陷是非致死性的,且能削弱泛素介 导的蛋白降解。
3、提高外源蛋白表达产率的突变宿主菌
能提高外源基因表达或分泌的突变类型
突变类型 生物效应 ssc1 改善外源蛋白分泌 ssc2 提高外源基因表达 rgr1 提高外源基因表达 ose1 提高外源基因表达 ssc11 改善外源蛋白分泌 rho 提高外源基因表达
作用位点 钙离子依赖型ATP酶 转录后加工 转录水平 转录水平 羧肽酶Y 转录水平
外侧糖链添加缺陷
如:人1-抗胰蛋白酶、人tPA在酿酒酵母 mnn9和och1突变株中高活性表达。
5、减少泛素依赖型蛋白降解的突变宿主菌
泛素介导的蛋白质降解作用
靶蛋白 Lys HOOC
泛素 76aa
泛素连接酶E3
靶蛋白 Lys 泛素连接酶E3
靶蛋白 Lys
蛋白 酶体
靶蛋白降解 泛素释放
酵母菌泛素依赖型蛋白降解系统的编码基因:
1981 Hitzeman用酿酒酵母成功表达人IFN。
1983 我国首次用酵母菌表达HBV HBsAg基因。
1996 完成第一个真核生物--酿酒酵母全基因组测序。
(一)酵母菌作为外源基因表达受体菌的特征
酵母菌(Yeast)是一群以芽殖或裂殖方式 进行无性繁殖的单细胞真核生物。
如果说E.coli是外源基因最成熟的 原核生物表达系统,Yeast则是最成熟 的真核生物表达系统。
酿酒酵母中的2环状质粒:
野生型,双链、环状, 6318bIRs: 反向重复序列,同源重组 FLP: 编码产物驱动IRs同源重组
REP1
FLP
IR
A ori IR
REP2
同源重组
REP: 编码产物控制质粒稳定性
B
STB: REP结合位点
人工构建酵母质粒的共同特点: 含有E.coli质粒复制起点,便于克隆操作; 含有多个单一酶切位点,便于克隆操作; 含有在酵母和E.coli中进行选择的双标记; 除YIp型质粒外均为穿梭载体。
无性繁殖(芽殖或裂殖)、单细胞、 真核生物; 繁殖方式与原核类似,易于操作; 基因表达调控机理与高等真核类似。
2、用于外源基因表达的酵母宿主菌
酿酒酵母(Saccharomyces cerevisiae):
最成熟的真核细胞表达系统; 表达水平低; 产物过度糖基化。
甲醇酵母:
巴斯德毕赤酵母 (Pichia pastoris) 多形汉逊酵母 (Hansenula polymorpha)
(二)酵母菌的宿主系统
用作模式真核生物的酵母宿主菌 用于外源基因表达的酵母宿主菌 提高重组蛋白表达产率的突变宿主菌 抑制超糖基化作用的突变宿主菌 减少泛素依赖型蛋白降解作用的突变宿主菌 内源性蛋白酶缺陷型的突变宿主菌
1、用作模式真核生物的酵母宿主菌
酿酒酵母(Saccharomyces cereviasiae):
如: rho突变株中人溶菌酶表达量提高10倍。
4、抑制超糖基化的突变宿主菌
酵母菌普遍拥有蛋白质糖基化系统, 但野生型酵母菌对外源蛋白的糖基化反应 很难控制,呈超糖基化倾向。
措施:构建超糖基化缺陷突变株。
抑制超糖基化的突变类型
突变类型
生物效应
mnn
甘露糖生物合成缺陷
alg
Asn侧链糖基化缺陷
och
野生型质粒:2质粒(酿酒酵母) 人工构建质粒:
酵母附加型质粒(YEp) 酵母复制型质粒(YRp) 酵母着丝粒质粒(YCp) 酵母人工染色体(YAC) 酵母整合型质粒(YIp)
除酵母外,其它几种酵母的细胞内也 含有野生型的相似质粒。
结合酵母属:pSR1、pSB1、pSB2 克鲁维酵母属:pKD1
YEp质粒:
复制子:源自2质粒;
拷贝数:50-100;
不稳定:培养几代后 ,质粒丢失率高达 50-70%,主要由于 分配不均匀所致。
YRp质粒:
复制子:源自酵母染色体的ARS;