基因组文库的构建.ppt
合集下载
目的基因的制备基因文库法ppt课件

组织或细胞染色体DNA 限带的所有基 因组DNA片段的集合.
克隆载体 重组DNA分子
受体菌 含重组分子的转化菌
genomic library
每个细胞含有一个基因组 DNA片段与载体的重组DNA分 子,许多细胞组成一个含有基 因组的色概率基因 该种生物所有基因 应用范围 ◆ 基因原始结构功能的研究 ◆ 对比分析内含子、外显子及调控区域 ◆ /46
逆转录
以单链RNA为模板合成双链DNA的反应。
DNA mRNA
14/46
逆转录病毒细胞内的逆转录现象:
RNA 模板 逆转录酶
RNA酶
DNA-RNA 杂化双链
Байду номын сангаас
单链DNA
双链DNA
逆转录现象说明:至少在某些生物,RNA同样具
有遗传物质的传代与表达功能。
15/46
逆转录酶的应用:
应用于基因工程
基因
mRNA 蛋白质多肽链
7/46
2.经验值
Transcription 从cDNA中获得的是经过剪切去除内含子的基因。
ln(1-P)
start codon Plasmid or phage
N=
非翻译区的序列特征对基因的表达具有重要的调控作用,编码序列则是合成基因产物—蛋白质模板。
ln(1-f/g) 某细胞的mRNA分子数为500 000,某个丰度为3500拷贝/细胞的mRNA基因,最小值为500 000/3500=143,丰度为14拷贝/细胞的mRNA
58330
人
3.2×109
320000
1473652
128000
589459
71111
327476
10/46
基因文库的构建

洗菌体碎片和 Pr
使单链DNA牢 固结合在膜上
• ⑵ 免疫结合法
• 噬菌斑转印到硝酸纤维膜上,各种cDNA 表达并呈现在噬菌体表面的蛋白质便牢 固地结合在膜上。然后将膜放入含有特 异抗体的溶液中进行免疫结合反应,洗 去未结合的抗体后,再与125 I 标记的第 二抗体结合, 从而找出带有放射性示踪信 号的斑点, 进而找出对应噬菌斑, 克隆扩 增, 提取DNA后经酶切可获目的基因。
衔接头:一端为平头,一端为粘末端。
(2)cDNA与载体相连
cDNA与一个载体在T4连接酶作用下以粘末端 互相连接,即为重组DNA 分子。
(3)导入宿主细胞 重组体在一定条件下导入宿主细胞培
养或保存。 宿主细胞必须是来自一个细胞的克隆
体,以保证它们的遗传性状相同。
筛选目的基因
⑴ 核酸杂交法 特殊标记的寡核苷酸链为 探针
利
用
基
因
文
库
筛
选某一Fra bibliotek基因
片 段
An efficient technique commonly used to detect a bacterial colony carrying a particular DNA clone.
4.7 DNA与蛋白质相互作用技术
•凝胶阻滞法
•DNase Ⅰ足迹法
4.7.1 凝胶阻滞实验
DNA来 源
基因组的大 小
(bp)
被克隆的 DN95 P=0.99
大肠杆 菌 酵母 果蝇
4.639X106 1.4X107 1.8X108
1.4X104 1.4X104 1.4X104
720 2300 2300
0
940 300皿内所形成的菌落或噬 菌斑)转印到硝酸纤维素膜上,碱解后加 带标记的探针进行杂交,能显示特异结合 探针的点(克隆)便是目的基因所在处。 确定菌落或噬菌斑的位置,扩增,提取, 再用限制性酶切出cDNA基因片断,即为目 的基因。(如图)
使单链DNA牢 固结合在膜上
• ⑵ 免疫结合法
• 噬菌斑转印到硝酸纤维膜上,各种cDNA 表达并呈现在噬菌体表面的蛋白质便牢 固地结合在膜上。然后将膜放入含有特 异抗体的溶液中进行免疫结合反应,洗 去未结合的抗体后,再与125 I 标记的第 二抗体结合, 从而找出带有放射性示踪信 号的斑点, 进而找出对应噬菌斑, 克隆扩 增, 提取DNA后经酶切可获目的基因。
衔接头:一端为平头,一端为粘末端。
(2)cDNA与载体相连
cDNA与一个载体在T4连接酶作用下以粘末端 互相连接,即为重组DNA 分子。
(3)导入宿主细胞 重组体在一定条件下导入宿主细胞培
养或保存。 宿主细胞必须是来自一个细胞的克隆
体,以保证它们的遗传性状相同。
筛选目的基因
⑴ 核酸杂交法 特殊标记的寡核苷酸链为 探针
利
用
基
因
文
库
筛
选某一Fra bibliotek基因
片 段
An efficient technique commonly used to detect a bacterial colony carrying a particular DNA clone.
4.7 DNA与蛋白质相互作用技术
•凝胶阻滞法
•DNase Ⅰ足迹法
4.7.1 凝胶阻滞实验
DNA来 源
基因组的大 小
(bp)
被克隆的 DN95 P=0.99
大肠杆 菌 酵母 果蝇
4.639X106 1.4X107 1.8X108
1.4X104 1.4X104 1.4X104
720 2300 2300
0
940 300皿内所形成的菌落或噬 菌斑)转印到硝酸纤维素膜上,碱解后加 带标记的探针进行杂交,能显示特异结合 探针的点(克隆)便是目的基因所在处。 确定菌落或噬菌斑的位置,扩增,提取, 再用限制性酶切出cDNA基因片断,即为目 的基因。(如图)
遗传学幻灯13ppt课件
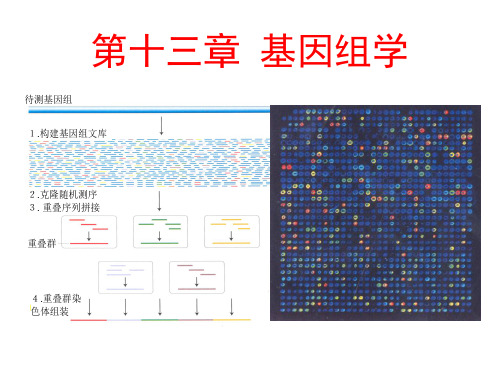
DNA片段之间的重叠顺序构建重叠群 (contig), 绘制物理连锁图 克隆DNA指纹图
重叠群:相互重叠
的DNA片段组成的
凝
物理图。克隆重叠
胶 电
泳
群的组建采用染色
分 离
酶
体步移法
切 片
段
物理图
③ 荧光标记原位杂交(FISH):
将荧光标记的探针与染色体杂
交确定分 子标记所
荧光原位杂交
在位置的
荧光染料 标记
检测
3、生物信息学的应用
1、发现新基因和新的单核苷酸 多态性
2、分析基因组中非编码蛋白质 区域功能
3、在基因组水平上研究生物进化 4、完整基因组比较研究
第四节 蛋白质组学
1、概念及研究内容 蛋白质组: 细胞、器官或组织的蛋 白质成分的总称 蛋白质组学: 研究这些成分在指定 的时间或特定的环境条件下的表达 研究内容:蛋白质表达模式,蛋白 质组功能模式
方法
变性与 杂交
④ 顺序标签位点(STS):
STS是长度在100~500 bp的DNA顺序,每个 基因组仅1份拷贝,很易分辨
当两个片段含有同一STS时,可以确认这两 个片段彼此重叠
两个不同的STS出现在同一片段的机会取决 于其在基因组中的位置。如果彼此邻接, 这两个STS总会同时出现在相同片段上。 如果相距甚远,有时会在同一片段,有时 则在不同片段
4、蛋白质间的相互作用
研究方法: 酵母双杂交系统 表面等离子共振技术
从 1996 年 酵 母 菌 基 因 组 全 序 列 测 定 后的4年多时间里: 全 世 界 1000 多 个 实 验 室 , 5000 多 名 科学家从事酵母菌后基因组学的研 究 发 表 论 文 7000 多 篇 , 鉴 定 1060 个 新 基 因 的 功 能 , 但 仍 然 还 有 约 1600 个阅读框架的功能不清楚 这些结果充分说明后基因组学研究 的复杂性
重叠群:相互重叠
的DNA片段组成的
凝
物理图。克隆重叠
胶 电
泳
群的组建采用染色
分 离
酶
体步移法
切 片
段
物理图
③ 荧光标记原位杂交(FISH):
将荧光标记的探针与染色体杂
交确定分 子标记所
荧光原位杂交
在位置的
荧光染料 标记
检测
3、生物信息学的应用
1、发现新基因和新的单核苷酸 多态性
2、分析基因组中非编码蛋白质 区域功能
3、在基因组水平上研究生物进化 4、完整基因组比较研究
第四节 蛋白质组学
1、概念及研究内容 蛋白质组: 细胞、器官或组织的蛋 白质成分的总称 蛋白质组学: 研究这些成分在指定 的时间或特定的环境条件下的表达 研究内容:蛋白质表达模式,蛋白 质组功能模式
方法
变性与 杂交
④ 顺序标签位点(STS):
STS是长度在100~500 bp的DNA顺序,每个 基因组仅1份拷贝,很易分辨
当两个片段含有同一STS时,可以确认这两 个片段彼此重叠
两个不同的STS出现在同一片段的机会取决 于其在基因组中的位置。如果彼此邻接, 这两个STS总会同时出现在相同片段上。 如果相距甚远,有时会在同一片段,有时 则在不同片段
4、蛋白质间的相互作用
研究方法: 酵母双杂交系统 表面等离子共振技术
从 1996 年 酵 母 菌 基 因 组 全 序 列 测 定 后的4年多时间里: 全 世 界 1000 多 个 实 验 室 , 5000 多 名 科学家从事酵母菌后基因组学的研 究 发 表 论 文 7000 多 篇 , 鉴 定 1060 个 新 基 因 的 功 能 , 但 仍 然 还 有 约 1600 个阅读框架的功能不清楚 这些结果充分说明后基因组学研究 的复杂性
基因组文库、cDNA文库的构建和筛选

mRNA的提取
• 全长mRNA具有 一个polyA的3’ 末端,可以被用 于mRNA的分离;
• 可以用寡聚纤维 素柱,选择性地 吸附mRNA
• 纯化
• 用结合有寡聚(dT )的磁珠加到细胞裂解 液,在用强磁铁吸出磁珠并洗出mRNA。
检测mRNA
• 翻译,检测翻译产物: 用无细胞翻译系统(如麦胚抽提液 或兔网织红细胞裂解液)来检测 mRNA的完整性,也可用凝胶电泳 直接检测,用杂交法分离 mRNA。
特特点点:: 对应的菌斑或噬菌斑位置不变, 可以直接找出阳性菌落。
寡聚核苷酸探针
抗体探针
• 表达筛选 通过抗体筛选来检测cDNA编码 的蛋白质
cDNA表达为与β半乳糖苷酶相嵌合的目的基因的融合 蛋白,用抗血清来筛选目的cDNA编码的融合蛋白。 筛选流程与噬菌斑杂交程序相似。
质粒蛋白 A
外源蛋白B
• 质粒载体:cDNA相对较短。 • 噬菌体载体:构建表达cDNA与载体的连接筛选流程
筛选• 从基因的大量克隆中鉴定出某个含有目的基 因的特定克隆的过程,称为筛选
• 筛选过程通常需要一个核酸探针进行杂交,该探 针可以与其互补序列结合从而检测出含目的基因 的克隆。
• 将菌落或噬菌斑转移到膜上,将其置入含放射性 标志探针溶液中保温,检出相应克隆。
• 合适的酶切片段可由琼脂糖凝胶电泳
质粒载体
蓝白筛选
YAC载体
置换型载体
• 允许外源 DNA 片段替换非必须 DNA 片段 的载体,称为置换型载体(replacement vectors)。一般情况下,置换型载体克隆 外源片段的大小范围是 9-2究 人 员 迫 切 地 寻 找 CFTR 及 HD 基 因 时 尚 无 YAC 可 用 。 疾 病 基 因 与 标 记 DNA 的 距 离 远 比 载 体 所 携 带 的 DNA 片 段 还 长 。 他 们 想 出 了染色体步移的方法。虽然标记 DNA 所 制 成 的 探 针 无 法 找 到 包 含 疾 病 基 因 的 片 段 , 但 所 找 到 的 片 段 (即 与 探 针 杂 交 的 片 段) 有 一 部 分 可 能 比 标 记 DNA 更 接 近 疾 病 基 因 。 在 他 们 制 作 的 基 因 库 里 , DNA 片 段 之 间 有 部分重叠。因此可以用新找到的片 段做为探针寻找与它重叠的片段。 如此重复许多次后,应该可以找到 包 含 疾 病 基 因 的 片 段。
7.DNA 文库的构建

用小体系连接来寻找最适的比 例。 在电转化和包装侵染过程中, 在电转化和包装侵染过程中,首先选择转化 效率高的感受态细胞或行脱盐处理也可以明显地提高 其效率。 其效率。 对于电转化而言,选择合适的转化电压对不 对于电转化而言, 同插入片段的 DNA 的转化效率也有所不同。 的转化效率功与否的关键,载 载体的好坏是影响连接成功与否的关键, 体的制备要求两点: 体的制备要求两点: 一是纯度高; 一是纯度高; 二是去磷酸化好。 二是去磷酸化好。
Hale Waihona Puke 2.高质量大分子量基因组 DNA的提取和 mRNA 分离; 分离; 第一, 第二,第一链 cDNA 合成; 合成; 第二, 第三,第二链 cDNA 合成; 合成; 第三, 第四,双链 cDNA 克隆进质粒或噬菌体载体并 第四, 导入宿主中繁殖。 导入宿主中繁殖。
表示重组克隆平均插入片段的大小和基表示重组克隆平均插入片段的大小和基因组因组dnad体噬菌体phagephage粘粒载体粘粒载体cosmidcosmid细菌人工染色体细菌人工染色体bacterialcosmidcosmid细菌人工染色体细菌人工染色体bacterialbacterialbacterial载体的选择载体的选择artificialchromosomesbacartificialchromosomesbac酵母人工染酵母人工染色体色体yacyacyeastartificialchromosomesyeastartificialchromosomesyacsyacsp1p1bacteriophagep1bacteriophagep1和DNA 片段是互补 DNA (complementary DNA , cDNA)。 cDNA cDNA)。 是由生物的某一特定器官或特定发育时期细胞内 的 mRNA 经体外反转胞内转录水平上的基因的群体, 时期细胞内转录水平上的基因的群体,并不能包 括该生物的全部基因,且这些基因在表达丰度上 括该生物的全部基因, 存在很大差异。 存在很大差D表明,提取的基因组 DNA 分子 实践表明, 量越大,所得到的重组克隆的插入片段越大。 量越大,所得到的重组克隆的插入片段越大。
Hale Waihona Puke 2.高质量大分子量基因组 DNA的提取和 mRNA 分离; 分离; 第一, 第二,第一链 cDNA 合成; 合成; 第二, 第三,第二链 cDNA 合成; 合成; 第三, 第四,双链 cDNA 克隆进质粒或噬菌体载体并 第四, 导入宿主中繁殖。 导入宿主中繁殖。
表示重组克隆平均插入片段的大小和基表示重组克隆平均插入片段的大小和基因组因组dnad体噬菌体phagephage粘粒载体粘粒载体cosmidcosmid细菌人工染色体细菌人工染色体bacterialcosmidcosmid细菌人工染色体细菌人工染色体bacterialbacterialbacterial载体的选择载体的选择artificialchromosomesbacartificialchromosomesbac酵母人工染酵母人工染色体色体yacyacyeastartificialchromosomesyeastartificialchromosomesyacsyacsp1p1bacteriophagep1bacteriophagep1和DNA 片段是互补 DNA (complementary DNA , cDNA)。 cDNA cDNA)。 是由生物的某一特定器官或特定发育时期细胞内 的 mRNA 经体外反转胞内转录水平上的基因的群体, 时期细胞内转录水平上的基因的群体,并不能包 括该生物的全部基因,且这些基因在表达丰度上 括该生物的全部基因, 存在很大差异。 存在很大差D表明,提取的基因组 DNA 分子 实践表明, 量越大,所得到的重组克隆的插入片段越大。 量越大,所得到的重组克隆的插入片段越大。
基因文库的构建

物体中,不同组织和细胞在不同时段的mRNA 种类不同(即基因基因组基因的构建程序载体和受体的选择
出于压缩重组克隆的数量,用于基因组构建的载体通常选装载量较大的l-DNA或考斯质粒;对于大型基因组(如动植物和人类)需使用YAC或BAC载体
由于绝大多数真核生物的几种载体的最大装载量如下:
质粒
01
02
03
04
05
06
07
08
09
10
ABC
DEF G
11
12
13
14
15
16
17
18
19
20
01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20
A
1
1
1
1
B
1
11
1
C
1
1111D Nhomakorabea1
1
11
1
E
1
1
1
1
F
1
1
11
G
1
1
1
1
01 04 12 06 13 14 02 14 07 19 11 15 05 08 16 0的克隆都是随机序列,必须将所有的克隆排 列成一个像天然染色体DNA上所表现出的信息基因的基本概念 基因的构建程序 基因组重组克隆的排序
基因的基本概念基因库与基因
基因库(gene pool) 特定生物e bank) 从特定生物个体中分离的全部基因,这些基因以克隆的形式 存序列为探针,进行第二步走 读,直至线型染色体DNA的端点
染色体走读法(chromosome walking)
亚克隆旁测序列
出于压缩重组克隆的数量,用于基因组构建的载体通常选装载量较大的l-DNA或考斯质粒;对于大型基因组(如动植物和人类)需使用YAC或BAC载体
由于绝大多数真核生物的几种载体的最大装载量如下:
质粒
01
02
03
04
05
06
07
08
09
10
ABC
DEF G
11
12
13
14
15
16
17
18
19
20
01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20
A
1
1
1
1
B
1
11
1
C
1
1111D Nhomakorabea1
1
11
1
E
1
1
1
1
F
1
1
11
G
1
1
1
1
01 04 12 06 13 14 02 14 07 19 11 15 05 08 16 0的克隆都是随机序列,必须将所有的克隆排 列成一个像天然染色体DNA上所表现出的信息基因的基本概念 基因的构建程序 基因组重组克隆的排序
基因的基本概念基因库与基因
基因库(gene pool) 特定生物e bank) 从特定生物个体中分离的全部基因,这些基因以克隆的形式 存序列为探针,进行第二步走 读,直至线型染色体DNA的端点
染色体走读法(chromosome walking)
亚克隆旁测序列
基因组作图ppt课件

➢ 经典遗传学中,遗传多态性指等位基因的变异;现代遗传 学中,遗传多态性指基因组中任何座位上的相对差异或 DNA序列的差异;
➢ 遗传标记可用于连锁分析、基因定位、遗传作图、基因转 移、辅助选择育种等;
15
ppt课件.
形态标记 (morphological markers)
细胞学标记 (cytological markers)
➢ 用具染色体变异的材料与正常材料杂交,特定染色体上的 基因在减数分裂过程中的分离和重组发生偏离,由此可测 定基因所在染色体及其位置;
➢ 克服了形态标记易受环境影响的缺点,但标记材料的产生 需大量的人力物力进行培养选择;
➢ 有些物种对染色体变异的耐受性差,难以获得相应的标19 记 材料。
ppt课件.
➢ 形态标记简单直观、经济方便, 容易观察记载。
17
ppt课件.
形态标记的不足
➢ 可以观察到的标记非常有限,难以建立饱和的遗传图谱; ➢ 许多形态标记受环境、生育期等因素的影响; ➢ 复等位基因位点很难全部鉴定、标记出来。
18
ppt课件.
2.1.2 细胞学标记
➢ 指能明确显示遗传多态性的细胞学特征。染色体的结构和 数量特征是常见的细胞学标记;
20世纪80年代后期,人们开始应用微卫星序列(microsatellite,MS)绘制图谱。1994
年底,美、法完成了以RFLP及微卫星DNA为标志的遗传图谱.图谱包含了
5826位点,覆盖4000cM,分辨率高达0.7cM.1996年法国报道了完全以微卫星
DNA标志构建的遗传连锁图,包含2335位点,分辩率为1.6cM
29
ppt课件.
30
RFLP标记的特征
ppt课件.
➢ 同一亲本及其子代相同位点上的多态性不变;
➢ 遗传标记可用于连锁分析、基因定位、遗传作图、基因转 移、辅助选择育种等;
15
ppt课件.
形态标记 (morphological markers)
细胞学标记 (cytological markers)
➢ 用具染色体变异的材料与正常材料杂交,特定染色体上的 基因在减数分裂过程中的分离和重组发生偏离,由此可测 定基因所在染色体及其位置;
➢ 克服了形态标记易受环境影响的缺点,但标记材料的产生 需大量的人力物力进行培养选择;
➢ 有些物种对染色体变异的耐受性差,难以获得相应的标19 记 材料。
ppt课件.
➢ 形态标记简单直观、经济方便, 容易观察记载。
17
ppt课件.
形态标记的不足
➢ 可以观察到的标记非常有限,难以建立饱和的遗传图谱; ➢ 许多形态标记受环境、生育期等因素的影响; ➢ 复等位基因位点很难全部鉴定、标记出来。
18
ppt课件.
2.1.2 细胞学标记
➢ 指能明确显示遗传多态性的细胞学特征。染色体的结构和 数量特征是常见的细胞学标记;
20世纪80年代后期,人们开始应用微卫星序列(microsatellite,MS)绘制图谱。1994
年底,美、法完成了以RFLP及微卫星DNA为标志的遗传图谱.图谱包含了
5826位点,覆盖4000cM,分辨率高达0.7cM.1996年法国报道了完全以微卫星
DNA标志构建的遗传连锁图,包含2335位点,分辩率为1.6cM
29
ppt课件.
30
RFLP标记的特征
ppt课件.
➢ 同一亲本及其子代相同位点上的多态性不变;
第三章基因文库的构建

目标基因组的大小 载体装载的平均片段= 重uestion2装载量 ➢ 所用的内切酶对基因组DNA的完全切割
频率和6Kb
★
G
Chemically synthesized
oligonucleotide
★
Anneal
TCAGGCT
T
★
Wild-type
(1) (1) DNA polymerase + 4 dNTPs
(2) T4 DNA ligase + ATP
正链DNA的合成
按照体外
DNA重组技术,将待突变的DNA片
G
段插入到M13噬菌体上. 然后制备
平末端 连接
加接头造成 粘性末端
细菌转化
重组噬菌体 DNA转染
重组质粒 的转化
体外包装 进行转导
重组体 的鉴定
表型鉴定
核酸杂交
免疫学分析
酶切A,把 所得到的大量基因组DNA片段与载体连接, 然后转化到细胞中去,让宿主菌长成克隆。 是某个生物体全部基因的随机片段的重组 DNA克隆群体。
SI核酸酶处理 Klenow 酶, 连接酶处理
16
一、缺失诱变 2、通过酶切图谱完成中间缺失
17
第三节:克隆DNA序列的体外诱变
二、盒式突变 1、原理:利用一段人工合成的具有突变序
列的寡核苷酸片段,取代野生型基因中的 相应序列。 2、人工合成的突变序列的特征
由两条合成的寡核苷酸组成的,当它们退火时,会按 设计要求产生出克隆需要的粘性末端, 由于不存在 异源双链的中间体,因此重组质粒全部是突变体。
(1) Screen plaques with 32P-labelled oligonucleotide as hybridization probe; (2) Isolate mutant
频率和6Kb
★
G
Chemically synthesized
oligonucleotide
★
Anneal
TCAGGCT
T
★
Wild-type
(1) (1) DNA polymerase + 4 dNTPs
(2) T4 DNA ligase + ATP
正链DNA的合成
按照体外
DNA重组技术,将待突变的DNA片
G
段插入到M13噬菌体上. 然后制备
平末端 连接
加接头造成 粘性末端
细菌转化
重组噬菌体 DNA转染
重组质粒 的转化
体外包装 进行转导
重组体 的鉴定
表型鉴定
核酸杂交
免疫学分析
酶切A,把 所得到的大量基因组DNA片段与载体连接, 然后转化到细胞中去,让宿主菌长成克隆。 是某个生物体全部基因的随机片段的重组 DNA克隆群体。
SI核酸酶处理 Klenow 酶, 连接酶处理
16
一、缺失诱变 2、通过酶切图谱完成中间缺失
17
第三节:克隆DNA序列的体外诱变
二、盒式突变 1、原理:利用一段人工合成的具有突变序
列的寡核苷酸片段,取代野生型基因中的 相应序列。 2、人工合成的突变序列的特征
由两条合成的寡核苷酸组成的,当它们退火时,会按 设计要求产生出克隆需要的粘性末端, 由于不存在 异源双链的中间体,因此重组质粒全部是突变体。
(1) Screen plaques with 32P-labelled oligonucleotide as hybridization probe; (2) Isolate mutant
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
重 组cDNA的总和,通常是在细菌或酵母中。 这些克隆代表从某个特定物种或器官的所 有mRNA制备得到的cDNA。
cDNA分子克隆(cDNA clone) :将 cDNA片段装在载体上转化细菌,组中分离单拷贝的基因;N
AAAAAAA mRNA
(b)寡聚(dT)引物
第一条 cDNA 的合成
An
An
Tn
(c)发夹引物
第二条 cDNA 的合成
TTTTTTຫໍສະໝຸດ GGGGGGCCCCCC
加接头 1
TTTTTT
GGGGGG
AAAAAA
CCCCCC
(d)同聚物加尾反应 TTTTTT
TTTTTT AAAAAA
连接 DNA 片段和载体
重组载体 DNA 导入 E.coli (体外包装或转化)
基因库的鉴定与段的产生与分离
一.基因组DNA的片段化
1.用限制酶片段化:
用限制酶消化DNA,不经凝胶电泳分部离,直接和载体 连接的克隆方法叫鸟枪法(shot-gan approach)
(2)有的基因表达具有严格的时空性,要获得 其mRNA并非易事。用不同发育阶段,或差A。不能用于基因结构和调节的研 究。一个基因经不同的剪接可产生不同的部 分重叠的cDNA克隆。
高速搅拌:1500转/分×30分 长度8kb的分子群体。
可切 平均
3.双酶消化
单酶消化的产物一般分子较大,选择时要考 虑到载体容量,如噬菌体插入片段不超过 25kb,为此可选用识别4bp的限制酶(切割平 均长度为256bp) ,识别6bp的限制酶切割平 均长度4096 bp.
双酶切产生的DNA片段平均大小不超过1kb, 但如采用双酶部分消化,产物的分子量 可达10~30kb,它们存在着随机序重叠, 用蔗糖梯度离心或凝胶电泳可将这些片 段群体按大小分开,可得到的分子量大 小约为20kb的随机DNA片段群体。
一般为99%; I:待克隆DNA片段的长度,假定为17kb; G: 基因组DNA的总长度,如人类为3×109bp
将数据代入上式
N ln(1 0.99) ln(1 17kb / 30Mb)
= 8.1×105 N的含义是克隆大小为1799%的RNA和tRNA因其丰 富和大小的一致性,可以通过分级直接分离)。 mRNA的提取可以通过多胸腺嘧啶的柱子分离。 然后被反转录。cDNA第一链的合成用oligo(dT) 引物,因为长的mRNA没有被完全反转录,因此 中部可加另一引物。第二链cDNA的合成是用 自身引物的,一个更有效的方法是在cDNA第 一链加尾(如多聚胞嘧啶),然后用oligo(G)为 引物起始第二链的合成。这样产生的双链 cDNA可以通过加接头或同聚尾巴插入载体。
存在的问题:
①所形成的重组体分子是一群带有大小不同插入片段 的混合群体。以每个载体可插入1~3kbDNA计算,
基因库至少含10~30万个重组质粒。从中筛出目的基 因工作量是很大的。
②目的基因内部可能有一个以上的酶切位点,即一个 基因分载在 几个重组质粒上,完整筛出更不容易。
2.随机片段化:
超声波:可产生300bp的短片段。
切除发夹
TTTTTT AAAAAA
通过以下途径将 cDNA 插入载体 a. 平端克隆 b. 加接头 c. 进一步进行同聚物加尾反应
加接头 2
TTTTTT AAAAAA 经酶切进行定向克隆
分离稀有的cDNA克隆。 建库的关键: 如何产生足够数量的重组DNA 建库应注意: 1.保证载体DNA和靶DNA均不被外源DNA
序列污染; 2.尽可能用“ 电话克隆”。
组织
mRNA cDNA
甲基化,加接头 的双链 DNA
DNA
部分或完全 消化的 DNA
载体
( 选择一种)
大小分级
可供克隆的 DNA 片段
二. DNA片断大小的分部
如已知含目的基因克隆片段的大小范围, 可在克隆前用蔗糖梯度离心或琼脂糖凝 胶电泳将DNA群体按大小分部分离。
三.目的基因克隆片段的富集。
) ln(1 I / G)
N:该序列的构建基因组(genomic library)
即基因(gene bank)是含有某种生物体全部基因的随机片断酶切使其成为一定大小的片段,连 接到适当载体(常为噬菌体)上,经体外包装 转染细菌,得到一群含不同DNA片段的重组 噬菌体颗粒。此即是基因。这群DNA片 段含盖基因组全部基因。
五. cDNA 一.概况: cDNA是通过反转录mRNA,并制备双链
cDNA(double strandeA的相对丰度;
(2)筛选方法:
除简单的分子杂交法外还可对表达产易于
筛4)建库时已排除了其它的RNA,使假筛选。
但应注意:
(1) 细胞中不同mRNA的丰度不同,低丰度的 mRNA要求很高的克隆数(要用公式来计算)。
cDNA分子克隆(cDNA clone) :将 cDNA片段装在载体上转化细菌,组中分离单拷贝的基因;N
AAAAAAA mRNA
(b)寡聚(dT)引物
第一条 cDNA 的合成
An
An
Tn
(c)发夹引物
第二条 cDNA 的合成
TTTTTTຫໍສະໝຸດ GGGGGGCCCCCC
加接头 1
TTTTTT
GGGGGG
AAAAAA
CCCCCC
(d)同聚物加尾反应 TTTTTT
TTTTTT AAAAAA
连接 DNA 片段和载体
重组载体 DNA 导入 E.coli (体外包装或转化)
基因库的鉴定与段的产生与分离
一.基因组DNA的片段化
1.用限制酶片段化:
用限制酶消化DNA,不经凝胶电泳分部离,直接和载体 连接的克隆方法叫鸟枪法(shot-gan approach)
(2)有的基因表达具有严格的时空性,要获得 其mRNA并非易事。用不同发育阶段,或差A。不能用于基因结构和调节的研 究。一个基因经不同的剪接可产生不同的部 分重叠的cDNA克隆。
高速搅拌:1500转/分×30分 长度8kb的分子群体。
可切 平均
3.双酶消化
单酶消化的产物一般分子较大,选择时要考 虑到载体容量,如噬菌体插入片段不超过 25kb,为此可选用识别4bp的限制酶(切割平 均长度为256bp) ,识别6bp的限制酶切割平 均长度4096 bp.
双酶切产生的DNA片段平均大小不超过1kb, 但如采用双酶部分消化,产物的分子量 可达10~30kb,它们存在着随机序重叠, 用蔗糖梯度离心或凝胶电泳可将这些片 段群体按大小分开,可得到的分子量大 小约为20kb的随机DNA片段群体。
一般为99%; I:待克隆DNA片段的长度,假定为17kb; G: 基因组DNA的总长度,如人类为3×109bp
将数据代入上式
N ln(1 0.99) ln(1 17kb / 30Mb)
= 8.1×105 N的含义是克隆大小为1799%的RNA和tRNA因其丰 富和大小的一致性,可以通过分级直接分离)。 mRNA的提取可以通过多胸腺嘧啶的柱子分离。 然后被反转录。cDNA第一链的合成用oligo(dT) 引物,因为长的mRNA没有被完全反转录,因此 中部可加另一引物。第二链cDNA的合成是用 自身引物的,一个更有效的方法是在cDNA第 一链加尾(如多聚胞嘧啶),然后用oligo(G)为 引物起始第二链的合成。这样产生的双链 cDNA可以通过加接头或同聚尾巴插入载体。
存在的问题:
①所形成的重组体分子是一群带有大小不同插入片段 的混合群体。以每个载体可插入1~3kbDNA计算,
基因库至少含10~30万个重组质粒。从中筛出目的基 因工作量是很大的。
②目的基因内部可能有一个以上的酶切位点,即一个 基因分载在 几个重组质粒上,完整筛出更不容易。
2.随机片段化:
超声波:可产生300bp的短片段。
切除发夹
TTTTTT AAAAAA
通过以下途径将 cDNA 插入载体 a. 平端克隆 b. 加接头 c. 进一步进行同聚物加尾反应
加接头 2
TTTTTT AAAAAA 经酶切进行定向克隆
分离稀有的cDNA克隆。 建库的关键: 如何产生足够数量的重组DNA 建库应注意: 1.保证载体DNA和靶DNA均不被外源DNA
序列污染; 2.尽可能用“ 电话克隆”。
组织
mRNA cDNA
甲基化,加接头 的双链 DNA
DNA
部分或完全 消化的 DNA
载体
( 选择一种)
大小分级
可供克隆的 DNA 片段
二. DNA片断大小的分部
如已知含目的基因克隆片段的大小范围, 可在克隆前用蔗糖梯度离心或琼脂糖凝 胶电泳将DNA群体按大小分部分离。
三.目的基因克隆片段的富集。
) ln(1 I / G)
N:该序列的构建基因组(genomic library)
即基因(gene bank)是含有某种生物体全部基因的随机片断酶切使其成为一定大小的片段,连 接到适当载体(常为噬菌体)上,经体外包装 转染细菌,得到一群含不同DNA片段的重组 噬菌体颗粒。此即是基因。这群DNA片 段含盖基因组全部基因。
五. cDNA 一.概况: cDNA是通过反转录mRNA,并制备双链
cDNA(double strandeA的相对丰度;
(2)筛选方法:
除简单的分子杂交法外还可对表达产易于
筛4)建库时已排除了其它的RNA,使假筛选。
但应注意:
(1) 细胞中不同mRNA的丰度不同,低丰度的 mRNA要求很高的克隆数(要用公式来计算)。