大豆起源与进化的研究进展
大豆起源于中国的分子证据

大豆起源于中国的分子证据大豆(学名:Glycine max),是人们饮食中非常重要的植物,也是世界上最重要的粮食和经济作物之一。
大豆的起源一直是植物学家和遗传学家们关注的重要问题之一。
许多研究表明,大豆起源于中国,并且有丰富的分子证据来支持这一观点。
首先,大豆与中国古代文化和历史有着密切的联系。
中国是世界上最早种植和利用大豆的国家之一。
早在公元前3000年的新石器时代,中国就已经开始种植大豆。
古代的农业文献《神农本草经》中就有对大豆的描述。
此外,在中国的陶器、织物和书籍中也出现了大豆的描绘。
这些文化和历史证据都支持大豆起源于中国。
其次,通过对大豆遗传基因的研究,科学家们发现了大豆起源于中国的分子证据。
研究表明,中国的野生大豆(Glycine soja)和栽培大豆(Glycine max)之间存在着很高的遗传相似性。
野生大豆是大豆的近缘物种,通常被认为是大豆的祖先物种。
研究人员分析了大豆和野生大豆的DNA序列,并通过比较两者的遗传差异来推断它们之间的演化关系。
研究结果表明,大豆和野生大豆之间的遗传距离较近,这表明它们具有共同的起源。
这一结论进一步支持了大豆起源于中国的观点。
此外,对大豆中特有的基因的研究也提供了进一步的证据。
大豆植物中特有的基因是指那些只在大豆中存在,而在其他植物物种中很少或没有的基因。
这些特有基因可以追溯到大豆物种的起源,并提供了关于大豆起源的重要信息。
通过对大豆中特有基因的研究,科学家们发现了一些具有中国特色的基因。
例如,研究发现大豆中的一个基因编码了一种特殊的油酸脂肪酸,这种脂肪酸在大豆中比其他作物更为丰富。
这种基因在野生大豆中存在,并且在普通的栽培大豆中也能找到。
这一研究结果表明,这个基因可能是在中国的野生大豆中首次出现,然后通过人工选择和培养而传递到栽培大豆中。
这个例子说明了大豆起源于中国的分子证据。
综上所述,大豆起源于中国的观点得到了丰富的分子证据和历史文化证据的支持。
大豆来源的知识解释

大豆来源的知识解释
大豆,也被称为黄豆,是一种重要的农作物,其来源和历史与中国文化紧密相连。
大豆的起源可以追溯到数千年前,据考古学家研究,中国黄河流域地区是大豆的原产地。
在中国古代,大豆被广泛种植,并成为了重要的粮食作物之一。
大豆不仅是一种营养丰富的食品,还是一种多功能的农作物。
它的种子富含蛋白质、脂肪和碳水化合物,是人们日常饮食中的重要来源。
此外,大豆还可以加工成各种豆制品,如豆腐、豆浆、豆腐干等,这些食品在中国及亚洲其他地区广受欢迎。
大豆的种植技术也经过了长时间的积累和改进。
在中国古代,农民们通过不断试验和实践,培育出了多个适应不同环境的大豆品种。
这些品种具有不同的生长习性和产量表现,使得大豆可以在更广泛的地区种植。
大豆在中国文化中也有着特殊的地位。
它不仅是一种食物,还象征着丰收和富足。
在古代文献中,大豆被多次提及,并被视为重要的农作物之一。
此外,大豆还与中国的传统节日和习俗相关联,如春节时的年糕、中秋节的月饼等,都少不了大豆制品的参与。
总之,大豆作为一种重要的农作物和文化符号,其来源和历史与中国文化紧密相连。
它不仅为人们提供了丰富的营养和美食,还代表了中国人民的智慧和勤劳。
在现代社会,大豆仍然是中国农业的重要组成部分,并在全球范围内发挥着重要的作用。
大豆的故事

大豆的故事大豆,作为一种重要的粮食作物,自古以来就在世界各地广泛种植。
它的历史可以追溯到公元前5000年左右的中东地区,当时人们已经开始种植和食用大豆。
随着时间的推移,大豆逐渐传播到其他地区,成为许多国家的主要农作物之一。
本文将讲述大豆的起源、传播、品种、营养价值以及在人类历史和现代社会中的作用。
一、大豆的起源大豆起源于中东地区的两河流域(今伊拉克一带),最早的栽培记录可以追溯到公元前5000年左右。
据考古学家发现,当时的人们已经开始种植和食用大豆,而且还用豆荚制作了各种工具和装饰品。
在古代印度次大陆,大豆也得到了广泛的种植和应用,如印度教的神像上常常雕刻有大豆的形象。
此外,大豆还在古代中国、希腊、罗马等文明中得到了推广和发展。
二、大豆的传播随着人类文明的不断发展,大豆逐渐传播到其他地区。
在公元前1世纪,张骞出使西域时,将大豆带回了中国。
此后,大豆在中国得到了广泛种植和应用,成为了中国人的重要粮食作物之一。
同时,大豆也通过丝绸之路传播到了欧洲和亚洲的其他地区。
在公元1世纪,罗马帝国开始大规模种植大豆,并将其作为军队的军粮。
此外,大豆还通过非洲和美洲大陆的探险家和殖民者传播到了新世界。
三、大豆的品种随着时间的推移,人们在大豆的培育过程中发现了众多的优良品种。
目前世界上主要的大豆品种包括:美国大豆、巴西大豆、阿根廷大豆、中国大豆等。
这些品种在产量、抗病性、营养价值等方面存在一定的差异。
例如,美国大豆通常具有较高的蛋白质含量和较低的脂肪含量;而巴西大豆则具有较强的抗旱能力和较高的产量。
此外,还有一些专门用于制作豆腐和豆浆的豆品种,如黄豆、青豆等。
四、大豆的营养价值大豆是一种营养丰富的食物,含有丰富的蛋白质、脂肪、碳水化合物、维生素和矿物质等营养成分。
其中,蛋白质是大豆的主要营养成分之一,占总重量的20%左右。
此外,大豆还含有丰富的不饱和脂肪酸、钙、铁、锌等矿物质以及维生素B族和E族等维生素。
因此,经常食用大豆有助于增强人体免疫力、预防心血管疾病、改善肝脏功能等作用。
大豆基因组研究进展

序列 中。 大豆 2 条染色体中 . 8 0 有 条染 色体在染 色体两端均有端粒重 复序 列( T GG Tr A G或 C C AA . 1 条染 色体仅在一 端有端粒重 C T A)有 1 复序列 ,有 1 9条染色体包含一个 9b 1p或 9 b 2 p的大片段着丝粒重复 区段 。 于大豆基因组序列共 预测出 6 , 5 基 91 个大豆基 因. 中 4 。 0 4 其 6 3 4 个基 因具有高置信 的蛋 白质编码 区 预测的基因中有 7 % 8 位于染色体 的末 端 . 尽管末端序 列仅包含 了不到一半 的基 因组序列 . 但却囊括 了 几乎所有 的遗传重组位点
【 关键词 】 大豆基 因组 ; 多倍体化 ; 重组 ; 测序
大豆[ i o ( . e 】 于我 国, 栽培历史 已有 5 0 G n m  ̄ L ) r 起源 e M r 其 00多 年之久【 十八世纪后期陆续传人欧洲和美洲 l _ , 大豆是世界上最重要的 油料作物和高蛋 白作物 . 具有重要经济价值 大豆蛋 白质 的氨基酸成 分与人类 的必需氨基酸模 式相类似 .尤其是含有 丰富的不饱 和脂肪 酸、 维生素和微量元素 . 是人类理想的食 品营养来源。 大豆还有很高的 药用价值 , 其含有 的两种有效成分—— 大豆异黄 酮和大豆皂苷 . 具有 广泛的药理作用和重要的药用价值 此外 . 大豆对新 的生 物能源的开 发和利用也具有重要作用 因此 . 大豆的遗传研究一直受 到广 泛的重 ◇ 职业教育◇ Fra bibliotek科技 圈向导
21 年第0 期 02 9
大豆基 因组研 究进展
张 乐 郭政阳 2 陈 曦 , 杨 靖 (. 1天津现代职业技术学 院 中国 天津 3 0 5 ; 0 3 0 2天 津 职 业 技 术 师 范 大 学 中 国 天 津 3 0 2 . 0 2 2 1
大豆起源

精心整理一、栽培大豆起源于中国是国内外学者所公认的栽培大豆起源于中国,早已为国内外学者所公认。
正像《美国大百科全书》所指出的:“中国古文献认为,在有文献记载以前,大豆便因营养价值高而被广泛地栽培。
同时在公元2000年前大豆便被看作是最重要的豆科植物。
”《苏联大百科全书》更明确指出:“栽培大豆原产于中国,中国在5000年前就开始栽培这个作物,并由中国向南部及东南各国传播,于16世纪进入欧洲。
”苏联着名的学者瓦维洛夫在《主要栽培植物的世界起源中心》一书中指出:“第一个最大的独立的世界农业发源和10“大豆的,本(● 1712● 1740● 1790● 1873 ● 1840● 1822● 1910● 1915● 1929年美国已种植大豆400万亩。
● 1941年二战期间,美国由于缺乏食用油,开始大规模种植大豆。
到1944年美国大豆种植面积达6000多万亩。
1972年美国大豆种植面积达4.28亿亩,产额达到1,234亿斤。
二、关于栽培大豆起源地的几种假说栽培大豆起源于中国,早已为国内外学者所公认,但是,栽培大豆究竟起源于中国何地何时却是众说纷纭的问题。
我们在这里对主要的几种假说作以下介绍并加以评论:1.黄河中下游起源说有的学者根据古代文献、考古文物、栽培大豆品种资源和野生大豆的分布,对栽培大豆的起源进行了分析(1985),提出栽培大豆起源数千年前,最早记载栽培大豆的地区是黄河中下游的河南、山西、陕西等地。
还有学者从农业的起源、以及古代文献、考古资料等方面对大豆起源进行分析以外,还联系野生大豆的分布和性状演化作了分析(1989),他们在全国野生大豆资源考察中发现黄河中下游的陕西、山西、河南,不仅野生大豆分布普遍,而且群落大,类型最为丰富,在野生的环境中,有多种进化程度的野生、半野生大豆存在。
因此,认为栽培大豆的主要起源地应该是黄河中下游地区。
2.河北东北部及东北中南部起源说有的学者认为,中国的北方地区不仅有考古发掘最早的大豆实物,最早的文字记载,而且也是我3.4.,加栽培大豆起源于黄河中下游的假说,所引用的古代文献和考古资料是不全面的,不准确的。
大豆生物学与基础遗传学研究

大豆生物学与基础遗传学研究大豆,是一种极为重要的作物,是我们日常饮食中的重要来源之一。
然而,对于许多人来说,大豆只是一个食材,一个常见的食品原材料。
实际上,大豆在农业、生物学甚至人类历史上都有着极为重要的地位,尤其是在基础遗传学研究中,大豆也是一个极为重要的模式生物。
从生物学层面来说,大豆是一种十分特别的植物,作为固氮植物,大豆拥有着许多独特的生理特征。
固氮植物能够通过根瘤中的根瘤菌将空气中的氮转化为植物可用的氮素,从而为自身提供重要的养分。
这对于农业来说具有巨大的意义,在农业生产中,种植水稻与小麦等传统农作物的同时,通过种植大豆来提高土地的养分,一直是农业生产中的主要策略之一。
另外,大豆还是一种极为复杂的植物系统,其基因组大小为950Mb,是除了烟草外已知的与基因组大小最近似的植物。
大豆的基因组拥有40,000-50,000个基因,其中富含优良的农艺性状基因,有着广泛的应用前景。
这也是为什么大豆成为了基础遗传学研究的理想模式生物的原因之一。
基础遗传学研究是现代遗传学的基础研究,是研究生命的基本结构和功能等遗传基础的学科。
在基础遗传学的研究中,大豆是一种非常重要的模式生物。
首先,大豆是一种极为复杂的群体,其基因组表现出了显著的基因组重塑现象。
研究大豆的基因组重塑在深入理解物种进化、遗传演化和基因组进化等方面均具有重要意义。
另外,由于大豆拥有着丰富的遗传变异,这使得研究人员可以从大豆中挑选出具有特定性状的个体进行深入研究。
这对于遗传学研究来说具有重要的意义,使得研究人员可以深入理解不同基因对物种形成和进化过程的影响。
在大豆的基因研究中,许多经典遗传学实验方法都能够被很好地应用。
例如孟德尔守则的适用已经被多次证实。
此外,大豆还被广泛应用在等位基因分析、基因定位和基因克隆方面的研究中,并在这些方面都取得了重要的研究成果。
在国内,大豆研究得到了广泛的关注和支持。
例如,华南农业大学成立了“国家基础研究重点发展计划大豆分子育种创新团队”,有力地促进了大豆基因研究和育种工作的开展。
大豆(Glycinemax)遗传转化实验技术原理
大豆(Glycinemax)遗传转化实验技术原理
展开全文
大豆(Glycine max)是一年生草本豆科植物,起源于东亚。
它的种子和豆荚都可食用,是人类重要的油和蛋白质来源。
另外,大豆还是研究植物各种生理功能如固氮和互利共生等的模式植物。
因此,大豆被认为是世界上非常有价值的作物之一。
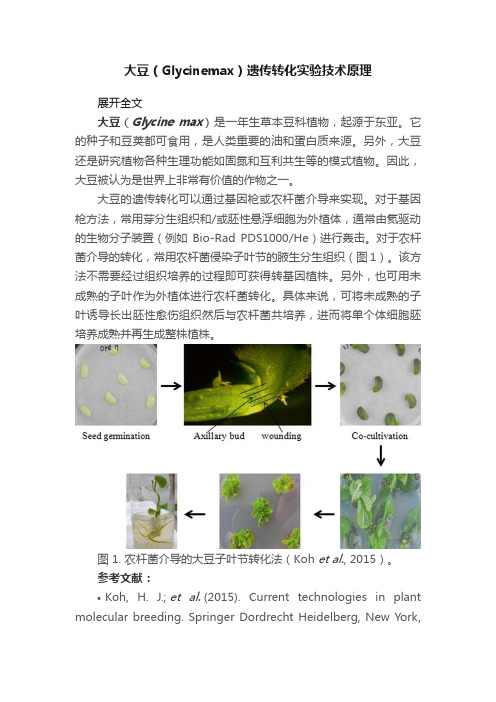
大豆的遗传转化可以通过基因枪或农杆菌介导来实现。
对于基因枪方法,常用芽分生组织和/或胚性悬浮细胞为外植体,通常由氦驱动的生物分子装置(例如Bio-Rad PDS1000/He)进行轰击。
对于农杆菌介导的转化,常用农杆菌侵染子叶节的腋生分生组织(图1)。
该方法不需要经过组织培养的过程即可获得转基因植株。
另外,也可用未成熟的子叶作为外植体进行农杆菌转化。
具体来说,可将未成熟的子叶诱导长出胚性愈伤组织然后与农杆菌共培养,进而将单个体细胞胚培养成熟并再生成整株植株。
图 1. 农杆菌介导的大豆子叶节转化法(Koh et al., 2015)。
参考文献:
•Koh, H. J.; et al. (2015). Current technologies in plant molecular breeding. Springer Dordrecht Heidelberg, New York,
London.。
我国大豆起源和历史发展研究

我国大豆起源和历史发展研究
大豆是我国传统的农作物之一,其起源历史可以追溯到古代。
大豆在我国的历史发展过程中扮演着重要的角色。
本文将从大豆起源、历史演变、生产情况等方面对其进行探讨。
一、大豆的起源
大豆的起源可以追溯到新石器时代,早期的野生大豆生长在中国的黄河流域、长江流域等地。
大豆是我国自主培育的一种农作物,对于世界范围内的农业和食品生产领域有着重要的贡献。
二、大豆的历史演变
大约在公元前2000年左右,我国出现了种植大豆的记录。
在这一时期,大豆主要被用作油料和饲料。
在唐朝时期,人们开始将大豆用于食品加工。
随着时间的推移,大豆的种植范围逐渐扩大,在我国的南方也开始广泛种植。
在清朝时期,大豆已经成为了我国的主要农作物之一。
三、大豆的生产情况
目前,我国是全球第一大大豆生产国。
大豆的种植主要分布在东北、华北以及黄淮海平原等地区。
大豆种植主要分为早稻、中稻、晚稻三个季节,其中早稻种植主要分布在黑龙江、吉林等地,晚稻种植则主要分布在江苏、安徽、湖南等地区。
四、大豆的研究与发展
随着科技的发展,大豆的研究也日益深入。
目前,我国食品加工领域对于大豆的利用越来越广泛,大豆可以被用于豆腐、豆浆、豆饼等食品的制作中。
同时,大豆的营养价值也得到了极大的关注,研究表明大豆富含蛋白质、维生素和多种营养物质,对人体健康有着很大的益处。
总之,大豆作为我国的传统农作物之一,具有着重要的历史渊源和生产价值。
大豆的研究和发展将促进我国的农业和食品工业的进步,同时也将带动我国经济的健康发展。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
大豆起源与进化的研究进展摘要:只有了解大豆的起源问题,才能更科学地指导大豆种质的搜集、分类、研究和利用。
此外,研究大豆的进化,可以更加清楚地了解大豆遗传特性的演替规律,为改良和培育大豆新品种提供理论依据。
栽培大豆起源于中国,但是起源的时间及在我国的发源地是一个历史久远的问题,本文就大豆起源的假说及大豆进化的研究进展加以论述。
关键词:大豆;起源;进化中国自古就栽培大豆,商朝甲骨文中有菽的象形文字。
我国最古老的、记述史前文化的、著于元前六世纪的《诗经》,便记载有“蓺之荏菽、荏菽旆旆”。
由此推断我国大豆种植历史约在5 000年上下。
世界其它国家的大豆,都是直接或间接从我国传播去的。
公元前三世纪大豆由我国传入朝鲜,六世纪传至日本。
约在300年前,大豆传入菲律宾、印度尼西亚。
欧美认识大豆则在18 世纪以后[1,2]。
国际学者均认同栽培大豆起源于中国,但在中国何处有多种假说。
瓦维洛夫在作物八大起源中心学说中认为大豆为温带物种,起源于中国中西部山地及毗邻低地[3]。
Hymowitz[4]提出栽培大豆起源于黄河流域并得到许多研究者的支持。
Fukuda[5]认为栽培大豆起源于中国东北。
王金陵[6,7]则推测大豆起源于华南或印度附近。
吕世霖[8]认为大豆可能有多个起源中心。
近期一些研究支持后两种假说[9~12]。
还有一些日本学者则认为有些日本栽培大豆可能不是从中、朝传播过去而是直接由日本本地野生大豆群体驯化的[12~14]。
研究一个古老作物的起源应该回答5个方面的问题:出自何物种何种群、何地、何时、由何人群驯化以及演化的经过。
这是一个难题,因史前文化难以确切追踪取得实据,但又是一个重要命题,这对了解一个古国的文化,民族的历史,物种的进化,科学的发展都至关重要。
笔者将对以上多种大豆起源假说作一评述,从大豆属物种的系统进化和栽培大豆起源研究的方法等方面分析栽培大豆起源的多种假设及依据,并提出进一步研究的问题和意见。
1 大豆起源1.1 东北起源学说日本学者福田[15]认为大豆起源于中国东北,其理由一是中国东北广泛分布有半野生大豆,中国其它地方则不多。
二是中国东北有众多的大豆品种,其中许多品种似乎有原始性。
李福山[16]也认为,中国的东北地区有考古发掘的最早的大豆文物,还有最早的大豆文字记载,同时东北也是我国野生大豆种质资源多样性最丰富的地区。
东北起源学说主要基于古文字、考古出土遗物及古农史。
考古发现有其必然性的一面,也有局限性的一面。
局限性的一面就在于出土的文物及文献年代久远,在准确性和全面性上不能令人信服。
而且古农史对南北大豆的栽培都有记载,因此仅凭考古和古农史的记载是不够的。
1.2 南方起源学说该学说是根据不同生态地区间各种进化类型大豆DNA指纹的差异比较,为大豆起源提供有力证据。
庄炳昌等[17]利用RAPD标记对不同纬度的野生大豆及栽培大豆进行了比较研究,发现南方野生大豆和栽培大豆的指纹图谱相似性要高于北方,指出南方北纬25地区在大豆起源中应值得重视。
盖钧镒等[18]发现南方野生大豆与栽培大豆的细胞器DNA RFLP标记最为接近,结合等位酶、形态等方面的研究结果,提出栽培大豆起源于南方的观点。
应该说南方起源学说所提出的论据是最先进的,它采用了分子生物学的方法对南方栽培大豆的DNA指纹和细胞器DNA与野生大豆进行比较,其相似性均高于北方。
田清震认为与其它各种方法相比较,来自分子生物学的证据更有说服力。
1.3 黄河中下游起源说Hymowitz[19]在《论大豆的驯化》一文中提出,大豆起源于中国北部的东半部,时间是我国的周朝。
他指出大豆在全国都有种植,但主要产区为高粱一大豆区(辽宁、吉林和黑龙江)和冬麦高粱区(山东以及河南、河北、江苏和安徽省大部分,即黄河中下游地区)。
王连铮[20]和王书恩[21]分别从古代文献、甲骨文、新石器时期的文化遗存和农业起源,考古发现,大豆演化以及植物生态学和植物学诸方面论证了大豆起源于黄河流域中下游。
徐豹等从生态学、品质化学和蛋白质生化遗传学三个方面研究了大豆的起源地问题。
在短光照处理试验中,北纬35的野生大豆与栽培大豆的开花日数最小。
由此向北,野生大豆比栽培大豆提早开花,但野生大豆开花日数短于栽培大豆;向南则延迟开花,但开花日数较长。
从而认为北纬35地区是光周期反应的临界点。
同时,徐豹还对来于不同纬度的1695份野生大豆和1635份栽培大豆蛋白含量进行了测定,其结果表明: 北纬34 ~北纬35地带野生大豆与栽培大豆的蛋白质含量最为接近。
向北野生大豆蛋白质含量上升,栽培大豆则下降; 向南野生大豆上升多,栽培大豆上升少,这一现象也被认为是大豆起源于北纬34 ~北纬35地带的生物学证据。
蛋白质电泳分析结果指出,我国北纬32 ~北纬37地区包括长江以北广大的黄河流域,野生大豆种子胰蛋白酶抑制剂的等位基因T i频率为100%,与栽培大豆频率相似。
由此向东南和东北方向,T i频率递减。
因此认为原始栽培大豆是由以北纬35为中心的黄河流域的野生大豆演化来的。
徐豹[22]对不同纬度的5147份野生、半野生大豆脂肪含量分析,发现野生大豆与半野生大豆脂肪含量相近地区,也在黄河流域,这也可能与大豆起源有关。
这一假说无论是从古农史、生态学还是在生物化学方面均有论证,因此支持这一观点的学者较多。
但这一假说缺少最令人信服的分子生物学方面的证据,而且也有学者认为栽培大豆起源于黄河中下游的假说,所引用的古代文献和考古资料是不全面的、不准确的。
因此这一观点也没受到全面的认同。
1.4 起源于长江流域或长江以南的假说有的学者根据长江流域及其以南地区有野生大豆分布,而且种植比较原始的泥豆与野生大豆,加上南方大豆短光性强于北方,因此认为大豆起源于长江流域及其以南地区。
栽培大豆起源于长江流域或长江以南的假说,主要是以大豆短日性的强弱为依据的。
持此种说法的学者认为大豆的短光性是大豆的原始性状,而南方的大豆短光性最强,因此,认为长江以南应该是大豆的起源中心。
这一假说论据单一,而且其论据也不具备很强的说服力。
虽然大豆的短光性是大豆的原始性状,但不一定越原始的大豆短光性就越强。
不论是野生大豆还是栽培大豆,其短日性的强弱都是自然选择的结果。
具体来说,在日照较长的高纬度地区(例如华北和东北),只能形成短日性较弱生态类型的大豆; 而在日照较短的低纬度地区(例如长江以南地区) ,只能形成短日性较强生态类型的大豆。
因此,短日性强弱,不能成为大豆起源的依据。
1.5多中心起源学说提出栽培大豆起源多中心说有三点依据:其一、栽培大豆的近缘祖先种野生大豆在我国各地均有分布,而且在我国古代文献中,黄河流域、华南地区以及四川等地均有栽培大豆的记录;我国各地都有文化发达较早的地区,因此,当地的远古居民都有将野生大豆驯化为栽培大豆的可能。
其二、我国各地不仅有野生大豆的分布,而且均有进化程度近于野生大豆的中间过渡类型和进化程度较高的栽培大豆,各地不同进化类型大豆的形态性状又非常相似。
其三、野生大豆的短日照性强弱具有明显的地区特异性,这种差异是由于长期适应当地自然环境而形成的,而野生大豆短日性的强弱不是衡量野生化程度的标准,因此野生大豆短日性的地区性和野生大豆分布的广泛性,恰恰证明栽培大豆起源多中心的可能性。
周新安等[23]认为中国栽培大豆起源中心为由西南向东偏北方向延伸的带状区域,包括河北(含北京) 、山东、山西、河南、陕西、四川等省(市); 北方春大豆起源中心可能在我国黄河流域中下游地区,以后向东北和西北扩散,南方地区春大豆和南方夏大豆的起源地可能在四川,以后向南、东南方向传播。
多起源学说虽然提出了三点论据,但每一点都有不足之处。
第一,黄河流域、四川及华南地区对大豆的最早记载分别距今4000年、2100年和2500年以上,彼此相距时间太长,不可能同时成为起源中心。
所以通过文史不能证明大豆的多起源中心。
第二,中国北方大粒及主茎进化程度显著高于南方地区,因此,各地均有进化程度较高,粒大、粒圆、茎秆粗壮的栽培大豆之说不并准确。
第三,论据并未说明其证据与栽培大豆的生态类型的相关性,因此具有不足之处。
以上各种学说都有各自的证据,但证据的丰富程度不同,研究角度也不一样,还没有得到能解释多方面现象的结论,都有待于进一步的证实。
2大豆遗传进化研究进展2. 1形态及农艺性状栽培大豆(G ly cine m ax (L. ) M err. )是由野生大豆(G ly cine soj a Sieb. et Zucc. ) ,经过人工选择,在栽培环境长期不断积累细小变异的结果,与人工选择关系密切的形态及农艺性状,最先受到大豆遗传进化学家们的重视。
王金陵[24]比较了野生、半野生和栽培大豆的一些性状指出,在野生大豆向栽培大豆的进化是细小变异定向积累的过程,粒大小、开花期早晚,茎粗细最足以代表进化的程度。
舒世珍[25]指出与进化相关程度依次为叶面积、产量性状、生育期、株高、节数、分枝数。
徐豹等[22]发现生物学相关性状在不同进化类型大豆上有连性表现,为栽培大豆由野生大豆积累微小基因突变进一步进化提供了具体资料。
陆静梅[26]还发现,野生到栽培大豆,花由分离状龙骨瓣趋于愈合状,符合Goethe关于花是适合于繁殖作用的变态枝理论。
形态学性状受环境影响较大,不同材料、不同试验条件都会对试验结果产生较大的影响。
严格控制试验条件,多种纬度不同进化类型的代表性材料综合比较分析,可能会提供更有价值的资料。
豆幼根在紫外光下呈现萤光,这是受单显性基因F r控制的。
李福山[16]对1210份栽培大豆和1310份野生大豆分析的结果,栽培大豆的有萤光的占97% ,野生大豆占64% ,即随着进化程度的提高,这种基因型的频率增加。
2.2 大豆组织器官的进化朱俊义对野生大豆及栽培大豆根皮层的研究表明,从野生大豆到栽培大豆根的皮层数目由少到多,皮层厚度由薄到厚,内皮层凯式带从无到有。
周波对大豆叶片进行研究表明,野生大豆叶表面的气孔数目是栽培大豆的三分之一,并且叶肉细胞中的栅栏组织较短。
陆静梅对大豆花结构的研究表明,野生大豆花的龙骨瓣是分离的,而栽培大豆则是完全愈合的。
陆静梅[26]通过扫描电镜还发现大豆次生木质部中的射线结构、导管的侵填体分布等,从野生、半野生到栽培大豆呈连续性变化。
Takhtaian[27]认为导管尾端长者为原始,短者则进化,而尾端完全消失为最进化。
2.3 生理分析研究杨文杰等[28]对野生、栽培大豆的生理测定表明,野生大豆单位叶片鲜重叶绿素含量高于栽培大豆,而叶绿素a、b值小于栽培大豆;野生大豆光饱和点低于栽培大豆,具有阴生植物的某些光合特性;开花结荚期的光合速率,随进化程度的提高而增加。
ИгамберлиеВ等[29]研究表明,栽培种羟基乙酸氧化酶和过氧化氢酶活性相当高,栽培大豆叶片光呼吸作用较强,由野生大豆进化到栽培大豆,强化了微体-乙醛酸循环体和过氧化物酶体有关的代谢。